
Laminaria hyperborea as a Source of Valuable Glyceroglycolipids—A Characterization of Galactosyldiacilglycerols in Stipe and Blade by HPLC-MS/MS



Abstract
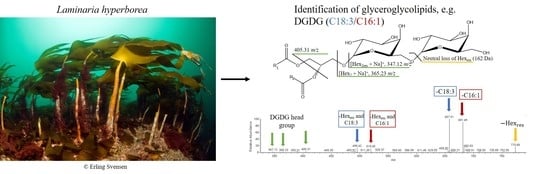
:
1. Introduction
2. Materials and Methods
2.1. Sampling and Pretreatment
2.2. Lipid Extraction and Determination of Total Lipid Content
2.3. Solid Phase Extraction
2.4. Analysis of Fatty Acid Methyl Esters by Gas Chromatography-Mass Spectrometry
2.5. Analysis of Glyceroglycolipids by High Performance Liquid Chromatography-Tandem Mass Spectrometry
2.6. Data Analysis
3. Results and Discussion
3.1. Total Lipid Content and Fatty Acid Profile
3.2. Identification of Glyceroglycolipids by HPLC-ESI-MS/MS Analysis
3.3. Characterization of Mono- and Digalactosyldiacylglycerols in Laminaria Hyperborea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MGDG | ||||||||
|---|---|---|---|---|---|---|---|---|
| Blade | Stipe | |||||||
| C:DBE | m/z [M + Na]+ | Bioactivity | Peaks | Isomers | Acyl Chain Pair a | Peaks | Isomers | Acyl Chain Pair a |
| 28:0 | 697.48 | 1 | 2 | 14:0/14:0 | ||||
| 30:0 | 725.51 | 1 | 1 | 14:0/16:0 | 1 | 1 | 14:0/16:0 | |
| 30:1 | 723.50 | AT [11] | 1 | 2 | 14:0/16:1 | 1 | 2 | 14:0/16:1 |
| 31:1 | 737.51 | 2 | 4 | 14:0/17:1 | ||||
| 32:1 | 751.53 | 2 | 3 | 14:0/18:1 | 1 | 2 | 14:0/18:1 | |
| 32:2 | 749.51 | 1 | 2 | 14:0/18:2 | 2 | 3 | 14:0/18:2 | |
| 32:3 | 747.50 | 1 | 1 | 14:0/18:3 | 4 | 11 | 14:0/18:3 | |
| 33:1 | 765.54 | 1 | 3 | 15:0/18:1 | 2 | 5 | 15:0/18:1 | |
| 34:1 | 779.56 | I [42], AA [7] | 1 | 1 | 16:0/18:1 | 1 | 2 | 16:0/18:1 |
| 34:2 | 777.54 | 2 | 2 | 16:0/18:2 | ||||
| 34:3 | 775.53 | 1 | 1 | 16:0/18:3 | 6 | 14 | 16:2/18:1 | |
| 34:4 | 773.51 | AI [15] c | 2 | 7 | 18:4/16:0 | 6 | 15 | 14:0/20:4 |
| 34:5 | 771.50 | AI [43] d | 3 | 7 | 16:1/18:4 | 2 | 4 | 20:5/14:0 |
| 34:6 | 769.48 | 1 | 2 | 16:2/18:4 | ||||
| 34:7 | 767.69 | 4 | 9 | 14:0/20:7 or 15:7/19:0 b | ||||
| 34:8 | 765.54 | 1 | 1 | 16:4/18:4 | ||||
| 35:1 | 793.58 | 1 | 3 | 17:0/18:1 | ||||
| 36:1 | 807.59 | 3 | 8 | 18:0/18:1 | ||||
| 36:2 | 805.58 | 2 | 5 | 18:1/18:1 | 3 | 6 | 18:1/18:1 | |
| 36:3 | 803.56 | 1 | 1 | 18:2/18:1 | 4 | 6 | 18:2/18:1 | |
| 36:4 | 801.54 | 1 | 1 | 18:3/18:1 | ||||
| 36:5 | 799.53 | AI [15] d | 4 | 11 | 18:4/18:1 | 5 | 11 | 20:5/16:0 |
| 36:6 | 797.51 | 3 | 8 | 20:5/16:1 | 9 | 26 | 18:4/18:2 | |
| 36:7 | 795.59 | 2 | 3 | 18:4/18:3 | 6 | 12 | 18:4/18:3 | |
| 36:8 | 793.48 | 1 | 1 | 18:4/18:4 | 7 | 13 | 18:4/18:4 | |
| 36:9 | 791.58 | 2 | 3 | 18:4/18:5 | 5 | 10 | 20:5/16:4 | |
| 38:5 | 827.56 | 3 | 6 | 20:4/18:1 | 8 | 21 | 20:4/18:1 | |
| 38:6 | 825.54 | 1 | 1 | 20:4/18:2 | 6 | 18 | 20:5/18:1 | |
| 38:7 | 823.53 | AI [43] | 6 | 12 | 20:5/18:2 | 6 | 13 | 20:5/18:2 |
| 38:8 | 821.51 | AI [16], AF [19] | 3 | 6 | 20:5/18:3 | 2 | 4 | 20:5/18:3 |
| 38:9 | 819.50 | AI [16], AF [19] | 1 | 1 | 20:5/18:4 | 1 | 1 | 20:5/18:4 |
| 40:6 | 853.58 | 1 | 2 | 22:5/18:1 | ||||
| 40:8 | 849.54 | AI [15] | 4 | 7 | 20:4/20:4 | 4 | 7 | 20:4/20:4 |
| 40:9 | 847.53 | 2 | 3 | 20:4/20:5 | 4 | 5 | 20:4/20:5 | |
| 40:10 | 845.51 | AI [15] | 1 | 1 | 20:5/20:5 | 1 | 1 | 20:5/20:5 |
| DGDG | ||||||||
|---|---|---|---|---|---|---|---|---|
| Blade | Stipe | |||||||
| C:DBE | m/z [M + Na]+ | Bioactivity | Peaks | Isomers | Acyl Chain Pair a | Peaks | Isomers | Acyl Chain Pair a |
| 30:1 | 885.64 | 1 | 2 | 14:0/16:1 | 1 | 2 | 14:0/16:1 | |
| 32:1 | 913.67 | 1 | 2 | 14:0/18:1 | 1 | 2 | 14:0/18:1 | |
| 32:2 | 911.65 | 1 | 2 | 14:0/18:2 | 3 | 4 | 14:0/18:2 | |
| 32:3 | 909.64 | AT [11] | 1 | 1 | 14:0/18:3 | |||
| 34:1 | 941.70 | 1 | 2 | 16:0/18:1 | 1 | 2 | 16:0/18:1 | |
| 34:2 | 939.69 | 2 | 4 | 16:0/18:2 | 2 | 2 | 16:1/18:1 | |
| 34:3 | 937.67 | 7 | 13 | 16:1/18:2 | ||||
| 34:4 | 935.72 | 2 | 5 | 18:4/16:0 | 6 | 14 | 20:4/14:0 | |
| 34:5 | 933.56 | AI [14] | 2 | 2 | 20:5/14:0 | |||
| 35:1 | 955.64 | 1 | 2 | 17:0/18:1 | 1 | 3 | 17:0/18:1 | |
| 36:1 | 969.65 | 1 | 1 | 18:1/18:0 | ||||
| 36:2 | 967.72 | 1 | 2 | 18:1/18:1 | 1 | 3 | 18:1/18:1 | |
| 36:3 | 965.70 | 1 | 2 | 18:2/18:1 | 3 | 6 | 18:2/18:1 | |
| 36:4 | 963.60 | 2 | 5 | 18:3/18:1 | 8 | 17 | 18:3/18:1 | |
| 36:5 | 961.59 | AI [15] | 3 | 7 | 20:5/16:0 | 3 | 6 | 20:5/16:0 |
| 36:6 | 959.57 | AI [44] | 1 | 1 | 20:5/16:1 | 6 | 12 | 20:5/16:1 |
| 36:7 | 957.65 | 3 | 5 | 18:3/18:4 | 2 | 5 | 20:5/16:2 | |
| 36:8 | 955.54 | 1 | 1 | 18:4/18:4 | ||||
| 38:5 | 989.62 | 3 | 7 | 20:4/18:1 | 4 | 7 | 20:4/18:1 | |
| 38:6 | 987.60 | 2 | 3 | 20:5/18:1 | 5 | 14 | 20:5/18:1 | |
| 38:7 | 985.68 | AI [43] | 2 | 6 | 20:5/18:2 | 6 | 17 | 20:5/18:2 |
| 38:8 | 983.67 | 2 | 3 | 20:5/18:3 | 3 | 6 | 20:5/18:3 | |
| 38:9 | 981.65 | 1 | 1 | 20:5/18:4 | 1 | 1 | 20:5/18:4 | |
3.3.1. Most Abundant Glyceroglycolipid Species
The Distribution of Carbon and Double Bond Equivalents in the Acyl Chains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gundersen, H.; Christie, C.H.; de Wit, H.; Norderhaug, K.M.; Bekkby, T.; Walday, M.G. Utredning om CO2-Opptak i Marine Naturtyper [CO2 Uptake in Marine Habitats—An Investigation]; Technical Report; Norwegian Institute for Water Research (NIVA): Oslo, Norway, 2011. [Google Scholar]
- Statistics Norway. 12847: Catch, by Landing County and Main Group of Target Species (C) 2014–2019. Statbank Norway. Available online: https://www.ssb.no/en/statbank/table/12847 (accessed on 8 September 2022).
- Harwood, J.L. Membrane lipids in algae. In Lipids in Photosynthesis: Structure, Function and Genetics; Siegenthaler, P.A., Murata, N., Eds.; Springer: Amsterdam, The Netherlands, 1998; pp. 53–64. [Google Scholar] [CrossRef]
- Hölzl, G.; Dörmann, P. Structure and function of glycoglycerolipids in plants and bacteria. Prog. Lipid Res. 2007, 46, 225–243. [Google Scholar] [CrossRef]
- Bruno, A.; Rossi, C.; Marcolongo, G.; Di Lena, A.; Venzo, A.; Berrie, C.P.; Corda, D. Selective in vivo anti-inflammatory action of the galactolipid monogalactosyldiacylglycerol. Eur. J. Pharmacol. 2005, 524, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Lopes, D.; Melo, T.; Meneses, J.; Abreu, M.H.; Pereira, R.; Domingues, P.; Lillebø, A.I.; Calado, R.; Domingues, M.R. A new look for the red macroalga Palmaria palmata: A seafood with polar lipids rich in EPA and with antioxidant properties. Mar. Drugs 2019, 17, 533. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Wang, H.; Guo, G.L.; Pu, Y.F.; Yan, B.U.; Wang, C.H. Isolation, purification, and identification of antialgal substances in green alga Ulva prolifera for antialgal activity against the common harmful red tide microalgae. Environ. Sci. Pollut. Res. 2016, 23, 1449–1459. [Google Scholar] [CrossRef]
- Imbs, T.I.; Ermakova, S.P.; Fedoreyev, S.A.; Anastyuk, S.D.; Zvyagintseva, T.N. Isolation of fucoxanthin and highly unsaturated monogalactosyldiacylglycerol from brown alga Fucus evanescens C Agardh and in vitro investigation of their antitumor activity. Mar. Biotechnol. 2013, 15, 606–612. [Google Scholar] [CrossRef]
- da Costa, E.; Melo, T.; Moreira, A.; Bernardo, C.; Helguero, L.; Ferreira, I.; Cruz, M.; Rego, A.; Domingues, P.; Calado, R.; et al. Valorization of lipids from Gracilaria sp. through lipidomics and decoding of antiproliferative and anti-inflammatory activity. Mar. Drugs 2017, 15, 62. [Google Scholar] [CrossRef] [PubMed]
- Hossain, Z.; Kurihara, H.; Hosokawa, M.; Takahashi, K. Growth inhibition and induction of differentiation and apoptosis mediated by sodium butyrate in Caco-2 cells with algal glycolipids. In Vitro Cell. Dev. Biol.-Anim. 2005, 41, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds Ulva armoricana, and Solieria chordalis from Brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef]
- Ulivi, V.; Lenti, M.; Gentili, C.; Marcolongo, G.; Cancedda, R.; Descalzi Cancedda, F. Anti-inflammatory activity of monogalactosyldiacylglycerol in human articular cartilage in vitro: Activation of an anti- inflammatory cyclooxygenase-2 (COX-2) pathway. Arthritis Res. Ther. 2011, 13, R92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenti, M.; Gentili, C.; Pianezzi, A.; Marcolongo, G.; Lalli, A.; Cancedda, R.; Cancedda, F.D. Monogalacto-syldiacylglycerol anti-inflammatory activity on adult articular cartilage. Nat. Prod. Res. 2009, 23, 754–762. [Google Scholar] [CrossRef]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Lall, S.P.; Craigie, J.S.; Hafting, J.T.; Critchley, A.T. Polar lipids from the marine macroalga Palmaria palmata inhibit lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophage cells. Phytochemistry 2014, 101, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Lall, S.; Craigie, J.S.; Hafting, J.T. Lipids isolated from the cultivated red alga Chondrus crispus inhibit nitric oxide production. J. Appl. Phycol. 2014, 26, 1565–1571. [Google Scholar] [CrossRef]
- Lopes, G.; Daletos, G.; Proksch, P.; Andrade, P.; Valentão, P. Anti-inflammatory potential of monogalactosyl diacylglycerols and a monoacylglycerol from the edible brown seaweed Fucus spiralis Linnaeus. Mar. Drugs 2014, 12, 1406–1418. [Google Scholar] [CrossRef]
- Plouguerné, E.; Ioannou, E.; Georgantea, P.; Vagias, C.; Roussis, V.; Hellio, C.; Kraffe, E.; Stiger-Pouvreau, V. Anti-microfouling activity of lipidic metabolites from the invasive brown alga Sargassum muticum (Yendo) Fensholt. Mar. Biotechnol. 2010, 12, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Al-Fadhli, A.; Wahidulla, S.; D’Souza, L. Glycolipids from the red alga Chondria armata (Kütz.) Okamura. Glycobiology 2006, 16, 902–915. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, E.H.; Lee, C.; Kim, M.H.; Rho, J.R. Two new monogalactosyl diacylglycerols from brown alga Sargassum thunbergii. Lipids 2007, 42, 395–399. [Google Scholar] [CrossRef]
- Reshef, V.; Mizrachi, E.; Maretzki, T.; Silberstein, C.; Loya, S.; Hizi, A.; Carmeli, S. New acylated sulfoglycolipids and digalactolipids and related known glycolipids from cyanobacteria with a potential to inhibit the reverse transcriptase of HIV-1. J. Nat. Prod. 1997, 60, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Deal, M.S.; Hay, M.E.; Wilson, D.; Fenical, W. Galactolipids rather than phlorotannins as herbivore deterrents in the brown seaweed Fucus vesiculosus. Oecologia 2003, 136, 107–114. [Google Scholar] [CrossRef]
- da Costa, E.; Azevedo, V.; Melo, T.; Rego, A.M.; Evtuguin, D.V.; Domingues, P.; Calado, R.; Pereira, R.; Abreu, M.H.; Domingues, M.R. High-resolution lipidomics of the early life stages of the red seaweed Porphyra dioica. Molecules 2018, 23, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, M.; Ishimaru, T.; Itabashi, Y.; Vyssotski, M. Glycerolipid composition of the red macroalga Agarophyton chilensis and comparison to the closely related Agarophyton vermiculophyllum producing different types of eicosanoids. Mar. Drugs 2019, 17, 96. [Google Scholar] [CrossRef]
- da Costa, E.; Melo, T.; Moreira, A.S.; Alves, E.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Decoding bioactive polar lipid profile of the macroalgae Codium tomentosum from a sustainable IMTA system using a lipidomic approach. Algal Res. 2015, 12, 388–397. [Google Scholar] [CrossRef]
- Kostetsky, E.; Chopenko, N.; Barkina, M.; Velansky, P.; Sanina, N. Fatty acid composition and thermotropic behavior of glycolipids and other membrane lipids of Ulva lactuca (Chlorophyta) inhabiting different climatic zones. Mar. Drugs 2018, 16, 494. [Google Scholar] [CrossRef] [PubMed]
- da Costa, E.; Domingues, P.; Melo, T.; Coelho, E.; Pereira, R.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomic signatures reveal seasonal shifts on the relative abundance of high-valued lipids from the brown algae Fucus vesiculosus. Mar. Drugs 2019, 17, 335. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.P.; Rey, F.; Melo, T.; Moreira, A.S.P.; Arbona, J.F.; Skjermo, J.; Forbord, S.; Funderud, J.; Raposo, D.; Kerrison, P.D.; et al. The unique lipidomic signatures of Saccharina latissima can be used to pinpoint their geographic origin. Biomolecules 2020, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Norwegian Centre for Climate Services. Observations and Weather Statistics. Available online: https://seklima.met.no/observations/ (accessed on 8 September 2022).
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Foseid, L.; Devle, H.; Ekeberg, D. Identification of fatty acids in fractionated lipid extracts from Palmaria palmata, Alaria esculenta and Saccharina latissima by off-line SPE GC-MS. J. Appl. Phycol. 2020, 32, 4251–4262. [Google Scholar] [CrossRef]
- Devle, H.; Ulleberg, E.K.; Naess-Andresen, C.F.; Rukke, E.O.; Vegarud, G.; Ekeberg, D. Reciprocal interacting effects of proteins and lipids during ex vivo digestion of bovine milk. Int. Dairy J. 2014, 36, 6–13. [Google Scholar] [CrossRef]
- Foseid, L.; Devle, H.; Stenstrøm, Y.; Naess-Andresen, C.F.; Ekeberg, D. Fatty acid profiles of stipe and blade from the Norwegian brown macroalgae Laminaria hyperborea with special reference to acyl glycerides, polar lipids, and free fatty acids. J. Lipids 2017, 2017, 1029702. [Google Scholar] [CrossRef] [PubMed]
- Pinkart, H.C.; Devereux, R.; Chapman, P.J. Rapid separation of microbial lipids using solid phase extraction columns. J. Microbiol. Methods 1998, 34, 9–15. [Google Scholar] [CrossRef]
- Ruiz, J.; Antequera, T.; Andres, A.; Petron, M.; Muriel, E. Improvement of a solid phase extraction method for analysis of lipid fractions in muscle foods. Anal. Chim. Acta 2004, 520, 201–205. [Google Scholar] [CrossRef]
- Schmid, M.; Stengel, D. Intra-thallus differentiation of fatty acid and pigment profiles in some temperate Fucales and Laminariales. J. Phycol. 2015, 51, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Mæhre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Van Ginneken, V.J.T.; Helsper, J.P.F.G.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from North Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef] [PubMed]
- Maciel, E.; Leal, M.C.; Lillebø, A.I.; Domingues, P.; Domingues, M.R.; Calado, R. Bioprospecting of marine macrophytes using MS-based lipidomics as a new approach. Mar. Drugs 2016, 14, 49. [Google Scholar] [CrossRef]
- Guella, G.; Frassanito, R.; Mancini, I. A new solution for an old problem: The regiochemical distribution of the acyl chains in galactolipids can be established by electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 1982–1994. [Google Scholar] [CrossRef]
- Block, M.A.; Jouhet, J.; Maréchal, E.; Bastien, O.; Joyard, J. Role of the envelope membranes in chloroplast glycerolipid biosynthesis. In Photosynthesis. Advances in Photosynthesis and Respiration; Eaton-Rye, J., Triphaty, B., Sharkey, T., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 34, pp. 191–216. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [CrossRef]
- Mizushina, Y.; Sugiyama, Y.; Yoshida, H.; Hanashima, S.; Yamazaki, T.; Kamisuki, S.; Ohta, K.; Takemura, M.; Yamaguchi, T.; Matsukage, A.; et al. Galactosyl-diacylglycerol, a mammalian DNA polymerase α-specific inhibitor from a sea alga, Petalonia bingbamiae. Biol. Pharm. Bull. 2001, 24, 982–987. [Google Scholar] [CrossRef]
- Banskota, A.H.; Stefanova, R.; Gallant, P.; McGinn, P.J. Mono- and digalactosyldiacylglycerols: Potent nitric oxide inhibitors from the marine microalga Nannochloropsis granulata. J. Appl. Phycol. 2013, 25, 349–357. [Google Scholar] [CrossRef]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Melanson, R.; Osborne, J.A.; O’Leary, S.J. Five new galactolipids from the freshwater microalga Porphyridium aerugineum and their nitric oxide inhibitory activity. J. Appl. Phycol. 2013, 25, 951–960. [Google Scholar] [CrossRef]
- Ma, A.C.; Chen, Z.; Wang, T.; Song, N.; Yan, Q.; Fang, Y.C.; Guan, H.S.; Liu, H.B. Isolation of the molecular species of monogalactosyldiacylglycerols from brown edible seaweed Sergassum horneri and their inhibitory effects on triglyceride accumulation in 3T3-L1 adipocytes. J. Agric. Food Chem. 2014, 62, 11157–11162. [Google Scholar] [CrossRef]
- Rey, F.; Lopes, D.; Maciel, E.; Monteiro, J.; Skjermo, J.; Funderud, J.; Raposo, D.; Domingues, P.; Calado, R.; Domingues, M.R. Polar lipid profile of Saccharina latissima, a functional food from the sea. Algal Res. 2019, 39, 101473. [Google Scholar] [CrossRef]
- Balboa, E.M.; Gallego-Fábrega, C.; Moure, A.; Domíınguez, H. Study of the seasonal variation on proximate composition of oven-dried Sargassum muticum biomass collected in Vigo Ria, Spain. J. Appl. Phycol. 2016, 28, 1943–1953. [Google Scholar] [CrossRef]
- Barbosa, M.; Fernandes, F.; Pereira, D.M.; Azevedo, I.C.; Sousa-Pinto, I.; Andrade, P.B.; Valentao, P. Fatty acid patterns of the kelps Saccharina latissima, Saccorhiza polyschides and Lami-naria ochroleuca: Influence of changing environmental conditions. Arab. J. Chem. 2017, 13, 45–58. [Google Scholar] [CrossRef]
- Fariman, G.A.; Shastan, S.J.; Zahedi, M.M. Seasonal variation of total lipid, fatty acids, fucoxanthin content, and antioxidant properties of two tropical brown algae (Nizamuddinia zanardinii and Cystoseira indica) from Iran. J. Appl. Phycol. 2016, 28, 1323–1331. [Google Scholar] [CrossRef]
- Gosch, B.J.; Paul, N.A.; de Nys, R.; Magnusson, M. Spatial, seasonal, and within-plant variation in total fatty acid content and composition in the brown seaweeds Dictyota bartayresii and Dictyopteris australis (Dictyotales, Phaeophyceae). J. Appl. Phycol. 2015, 27, 1607–1622. [Google Scholar] [CrossRef]
- Surget, G.; Le Lann, K.; Delebecq, G.; Kervarec, N.; Donval, A.; Poullaouec, M.A.; Bihannic, I.; Poupart, N.; Stiger-Pouvreau, V. Seasonal phenology and metabolomics of the introduced red macroalga Gracilaria vermiculophylla, monitored in the Bay of Brest (France). J. Appl. Phycol. 2017, 29, 2651–2666. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Optimisation of fucoxanthin extraction from Irish seaweeds by response surface methodology. J. Appl. Phycol. 2017, 29, 1027–1036. [Google Scholar] [CrossRef]
- Browse, J.; Warwick, N.; Somerville, C.R.; Slack, C.R. Fluxes through the prokaryotic and eukaryotic pathways of lipid synthesis in the ‘16:3’ plant Arabidopsis thaliana. Biochem. J. 1986, 235, 25–31. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foseid, L.; Devle, H.; Naess-Andresen, C.F.; Ekeberg, D. Laminaria hyperborea as a Source of Valuable Glyceroglycolipids—A Characterization of Galactosyldiacilglycerols in Stipe and Blade by HPLC-MS/MS. AppliedChem 2022, 2, 185-198. https://doi.org/10.3390/appliedchem2040013
Foseid L, Devle H, Naess-Andresen CF, Ekeberg D. Laminaria hyperborea as a Source of Valuable Glyceroglycolipids—A Characterization of Galactosyldiacilglycerols in Stipe and Blade by HPLC-MS/MS. AppliedChem. 2022; 2(4):185-198. https://doi.org/10.3390/appliedchem2040013
Chicago/Turabian StyleFoseid, Lena, Hanne Devle, Carl Fredrik Naess-Andresen, and Dag Ekeberg. 2022. "Laminaria hyperborea as a Source of Valuable Glyceroglycolipids—A Characterization of Galactosyldiacilglycerols in Stipe and Blade by HPLC-MS/MS" AppliedChem 2, no. 4: 185-198. https://doi.org/10.3390/appliedchem2040013