
Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges

1
Laboratory of Applied Biotechnology, Azm Center for Research in Biotechnology and Its Applications, Lebanese University, Tripoli 1300, Lebanon
2
Laboratoire de Chimie Agro-industrielle (LCA), Université de Toulouse, Toulouse INP, INRAE, 31030 Toulouse, France
3
School of Engineering, Lebanese American University, Byblos P.O. Box 36, Lebanon
4
Département Génie Biologique, IUT A, Université Paul Sabatier, 32000 Auch, France
5
Faculty of Public Health III, Lebanese University, Tripoli 1300, Lebanon
*
Author to whom correspondence should be addressed.
Phycology 2023, 3(1), 202-210; https://doi.org/10.3390/phycology3010013
Submission received: 31 December 2022
/
Revised: 10 February 2023
/
Accepted: 13 March 2023
/
Published: 19 March 2023
Abstract
:Penicillium digitatum causes severe damage to postharvest and stored citrus fruits, making it necessary to seek acceptable solutions limiting environmental and sanitary impacts. The seaweed liquid extracts (SLEs) obtained from the green macroalgae Ulva lactuca have previously shown anti-fungal activities towards postharvest green mold due to P. digitatum. Moreover, treatment with aqueous and ethanolic extracts had an influence on disease incidence and lesion severity on wounded oranges “Valentia late”. Therefore, we studied the abilities of the U. lactuca extracts and chlorophyll a (as a standard) to activate defense mechanisms in oranges. The analyses showed modifications in the protein concentrations and activities of β-1,3-glucanase and peroxidase in orange tissues, with a maximum activity level reached at 72 h after treatments. It appears that the SLEs were able to fight P. digitatum infections in the absence of direct contact with the pathogen, reflecting that the induction of host resistance is also a probable mode of action. Ulva lactuca fractions are rich in chlorophyll a, polyphenols, organic acids and ulvans, which might be the key molecules involved in the defense mechanism of oranges’ albedo. The use of these natural substances could be an alternative way to control postharvest citrus rot.
1. Introduction
Oranges are the major fruit crop cultivated in the Mediterranean area, which generates substantial economic activity through the productions of juice, essential oils, pectin and co-products [1]. Their vitamin C content plays a significant role in people’s health [2]. However, citrus fruits are infected by the mold Penicillium digitatum during their transportation and storage. It is one of the most devastating pathogens, causing between 30 and 80% of fruit losses [3]. P. digitatum infects fruit throughout the distribution and marketing process through spores’ dissemination in the air. Macarisin et al. [4] suggested that P. digitatum has the ability to suppress host defenses, in particular by inhibiting the production of hydrogen peroxide. Synthetic fungicides are used to control green mold and to reduce postharvest losses, but the use of chemicals has led to increased pathogen resistance, costs of new molecules and sanitary issues [5]. Safer alternatives for the control of green mold through the use of natural substances are being considered, possibly associated with antagonist microorganisms [6,7,8,9,10]. Numerous works have reported that natural bioactive molecules from marine algae are effective in protecting crops against agricultural pathogens [11,12,13]. Analyses performed by Salim et al. [14] revealed lipids and pigments in the ethanolic extract of U. lactuca, as well as monosaccharides, sulfated polysaccharides and organic acids such as succinic acid in the aqueous fraction. Both fractions exhibited anti-fungal activities against P. digitatum on orange tissues according to in vivo and in vitro tests [14,15]. Moreover, when lipids are extracted, they can form a film on the surface of the fruits and act as a physical barrier to pathogens [16]. However, the mechanisms of action of the natural substances within orange cells have yet to be studied. Indeed, some algal elicitors, able to induce a plant defense response, were identified [17,18].
Citrus fruit resistance is correlated with defense-related enzyme concentrations in the albedo (the inner white part), according to Ballester et al. [19]. Youssef et al. [20] proved that both sodium carbonate and sodium bicarbonate amplified the activity of β-1,3 glucanase, peroxidase and phenylalanine ammonia-lyase (PAL) in the tissues of oranges. Fallanaj et al. [21] also reported that an electrolyzed sodium bicarbonate solution induced both the direct inhibition of P. digitatum and an increase in enzyme activity and the accumulation of reactive oxygen species, generating oxidative stress in the fungus. Other studies on grape fruit indicated augmented activity of β-1.3-glucanase induced by chitosan combined with salicylic acid [22]. Abouraïcha et al. [23] highlighted the efficiency of oligosaccharides, including ulvans, in inducing the production of defense-related enzymes and of metabolites (phenols) in apple fruit.
Among the possible solutions to reduce the impacts of P. digitatum, we want to explore the role of some natural substances and identify the possible defense mechanisms activated by the U. lactuca extract. For this purpose, our intention was to measure the activities of enzymes (glucanase and peroxidase) in ‘Valencia late’ orange tissues.
2. Materials and Methods
2.1. Materials
Ulva lactuca was collected in April 2019 from a coastal region of the Mediterranean Sea near Tripoli (Lebanon). The algae were cleaned with distilled water (three times) and dried at room temperature (22 ± 2 °C). The samples of sea lettuce were stored at ambient temperature until their use. The used oranges (Citrus sinensis (L.) Osbeck, “Valencia late”) were collected from a local orchard in Tripoli (Lebanon). They were chosen for their uniform size without symptoms. The surface of the fruit was sterilized using a 2% commercial bleach solution for 2 min. They were then washed with tap water, and air-dried at ambient temperature.
Ethanol (99%) was procured from VWR international. Phosphate-buffered saline (PBS), potato dextrose agar (PDA), nystatin suspension (fungicide), chlorophyll a standard, sodium acetate buffer solution, sodium potassium tartrate, bovine serum albumin (BSA), Bradford reagent, 3,5-dinitrosalicylic acid (DNS), laminarin, guaiacol and malt extract agar were procured from Sigma Aldrich.
2.2. Obtention of the Seaweed Liquid Extracts (SLEs)
2.2.1. Preparation of the U. lactuca Aqueous Extract
The algal extract was prepared as described by Salim et al. [14]. Briefly, 100 mL boiling milli-Q water was mixed with 5 g dried algae in a 100 mL flask. The solution was stirred magnetically at room temperature for one hour. The residual solid material was then filtered through a double layer of sterile muslin cloth. The obtained liquid extract was stored at 4 °C until use.
2.2.2. Preparation of the U. lactuca Ethanolic Extract
Algal powder was subjected to maceration by adding 5 g to 100 mL ethanol. The mix was maintained under stirring at room temperature for 24 h. After filtration of the solid material, the liquid extract was stored at 4 °C.
2.3. Evaluation of U. lactuca Extracts as Protection Inducers
Oranges were wounded once (5 mm deep × 3 mm wide) with a sterile nail head along the equatorial axis. Extract of sea lettuce (100 µL) was deposited on each wound. The control oranges were those treated with sterile distilled water, ethanol, chlorophyll a standard (0.5 mg·mL–1 ethanol) or nystatin (fungicide). The treated oranges were positioned in a tray, which was positioned inside a polyethylene film. After two days of incubation at 20 °C and a high relative humidity, another wound was made approximately 5 mm away from the first one. This latter wound was air-dried and inoculated with 10 µL of a 104 conidia mL–1 suspension of P. digitatum. Each treatment was carried out in triplicate of four oranges, which were placed inside a polyethylene film (80% relative humidity) and maintained at room temperature for two weeks. The percentage of wounds with disease symptoms (disease incidence) and disease severity (lesion diameter, mm) were recorded.
2.4. Tissue Sampling for Analysis
Each orange was individually wounded with a sterile nail at eight points on the equatorial surface to obtain enough tissue to analyze. Thirty microliters of the presented treatments was applied to each wound. Samples for treatments had the compositions indicated in Table 1.
An ethanolic solution of chlorophyll a was chosen as a standard since this pigment is naturally present in the extract. The oranges were randomly arranged into five lots for tissue excision at different time intervals with a step of 12 h (0, 12, 24, 48, and 72 h). The first measurements were performed at the beginning of incubation. Each lot consisted of three replicates of four oranges. Experiments were repeated twice. Oranges were put into plastic boxes, then individually positioned in plastic bags (90–95% relative humidity) and stored at 20 °C for 3 days. At the determined time intervals, 8 tissue cylinders of 3 mm wide by 5 mm deep were taken from each lot from the inoculation site. The excised tissues were mixed and ground into a fine powder using a commercial blender and stored at −80 °C before assays of enzyme activities.
2.5. Enzyme Assays
According to Youssef et al. [20], for each sampling timepoint, 10 g of fine tissue powder (treated or nontreated) was homogenized in 50 mmol sodium acetate buffer at pH 5.6 (1:1, w/v). The mixture was centrifuged (15 min at 10,000× g and 4 °C) and the supernatant then filtered through filter paper using a Buchner funnel. Proteins were precipitated in 60% acetone (v/v) at −20 °C and the resulting pellets, following centrifugation (30 min at 10,000× g and 4 °C), were washed three times with 60% cold acetone. The pellets were dried, re-suspended in 2 mL 50 mmol sodium acetate buffer (pH 5.6) and stored at −20 °C until use. The protein concentration was determined according to the Bradford method [24] using the Bio-Rad protein assay kit (Bio-Rad Laboratories Ltd., Hercules, CA, USA).
2.5.1. The Activity of β-1,3-glucanase
The activity of β-1,3-glucanase was assessed following the method of Abeles et al. [25]. The mixture of 65 µL protein extract with 65 µL 4% (w/v) laminarin was incubated for 2 h at 37 °C in. The reaction was stopped by adding 300 µL 3,5-dinitrosalicyclic acid (DNS). The mixture was then heated and stirred at 70 °C. Then, 30 g sodium potassium tartrate (MW = 282.2 g·mol–1) was added, followed by cooling at ambient temperature. After the addition of water to 100 mL, the sample was heated in boiling water for 10 min and then rapidly cooled on ice. The absorbance was measured at 492 nm. The enzymatic activity was expressed as µmol glucose per µg of total protein min–1.
2.5.2. The Activity of Peroxidase Enzyme
The activity of peroxidase was evaluated using guaiacol as a substrate [26]. The reaction mixture was produced by adding 100 µL crude extract to 100 µL 50 mmol sodium acetate buffer pH 5.6 (10 mmol of guaiacol and 10 mmol H2O2). The mixture was then incubated for 60 s at ambient temperature. The rise in absorbance at 470 nm was evaluated through spectrophotometry (Beckman DU 640 Spectrophotometer, Corona, CA, USA) and the activity of the enzyme was expressed in U µg−1protein·s−1.
2.6. Statistical Analysis
An ANOVA analysis was carried out using the statistical software package Statistics for Windows (GraphPad prism). Means comparison was performed using Duncan’s test at the p < 0.05 probability level in order to investigate the significant differences. The in vivo experiments were performed using three repetitions of four oranges each. In the other experiments with artificially inoculated fruit, each of the three repetitions consisted of nine oranges. Each enzyme activity assay was performed three times.
3. Results and Discussion
Table 1 shows the characteristics of the two extracts selected for the treatment. As expected, the contents of total polyphenols and chlorophyll a were higher in the ethanolic extract. Ulvans made of water-soluble sulfated polysaccharides were identified through infrared spectroscopy in the aqueous extract, with the S=O stretching at 1258 cm−1 and the C=O stretchings (υas = 1640 cm−1 and υs = 1436 cm−1) of uronic acid (component of the sulfated heteropolysaccharide).
3.1. Effect of Extracts on Green Mold (Artificial Infection)
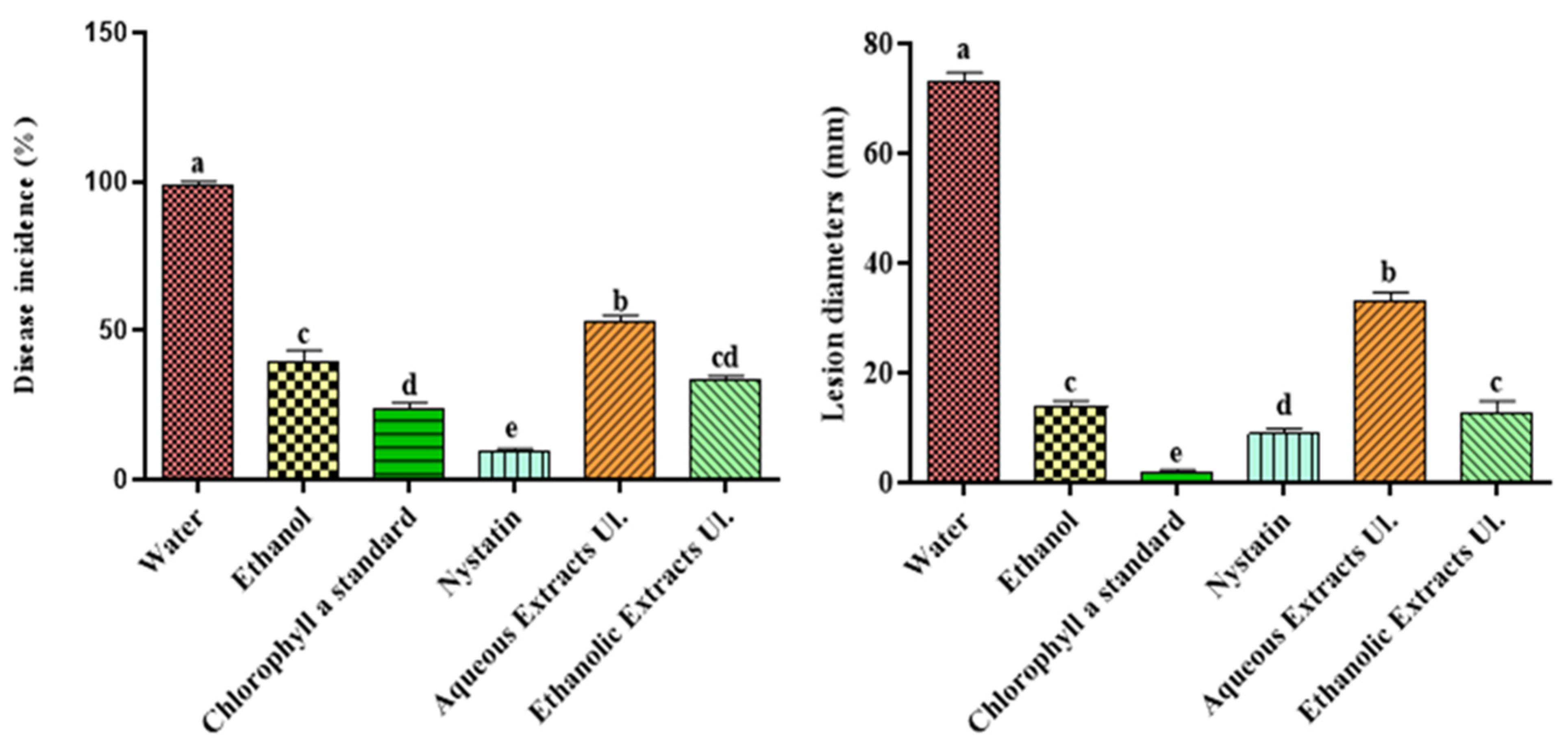
After inoculation of a wound (5 mm depth × 3 mm wide) with P. digitatum, the green mold incidence (as percentage of wounds presenting disease symptoms) and severity on oranges treated with ethanolic and aqueous extracts of sea lettuce was evaluated (Figure 1). We observed that the treatment based on chlorophyll a ethanolic solution lowered the disease incidence by 42% compared to the ethanolic standard. The aqueous extract also led to a decrease in disease incidence of 47% compared to the standard (water). With regard to disease severity, reductions in lesion diameters were recorded with the chlorophyll a solution (84%) and with the aqueous extract (55%) compared to the solvent standards. The ethanolic extract exhibited smaller reductions of about 15% compared to ethanol.
The measurements of disease incidence and lesion severity after the treatment with U. lactuca extracts indicated a limitation of contamination by P. digitatum, compared to water. The ethanolic solution of chlorophyll a divided the disease incidence by 2 and the severity by 5 compared to the effect due to ethanol. These results are consistent with the previous in vitro tests performed for the inhibitions of spore adhesion and germination by the U. lactuca extract [14]. Chlorophyll a was already identified by Bhagavathy et al. [27] as an effective microbial growth inhibitor. Its inhibiting action towards P. digitatum is explained by its tetrapyrrole structure, which is a chelating ligand for magnesium cation. Moreover, Li Destri Nicosia et al. [28] proved that phenolic compounds from pomegranate peel extracts (ethanol/water) contributed to a broad range of antifungal activity, including for P. digitatum on citrus fruit.
We suspected that peroxidase and β-1,3-glucanase were the key enzymes associated with the disease response to pathogens in plants by improving their resistance to abiotic and biotic stresses [29,30]. β-1,3-glucanase acts as an extracellular hydrolytic enzyme that contributes to altering the permeability of the fungal cell membrane [26] and inhibiting progression of the contamination. The role of peroxidase in the defense of plant cells is to serve as a catalyst of lignin formation and polyphenol accumulation to limit pathogen infection [18].
3.2. Protein Quantification
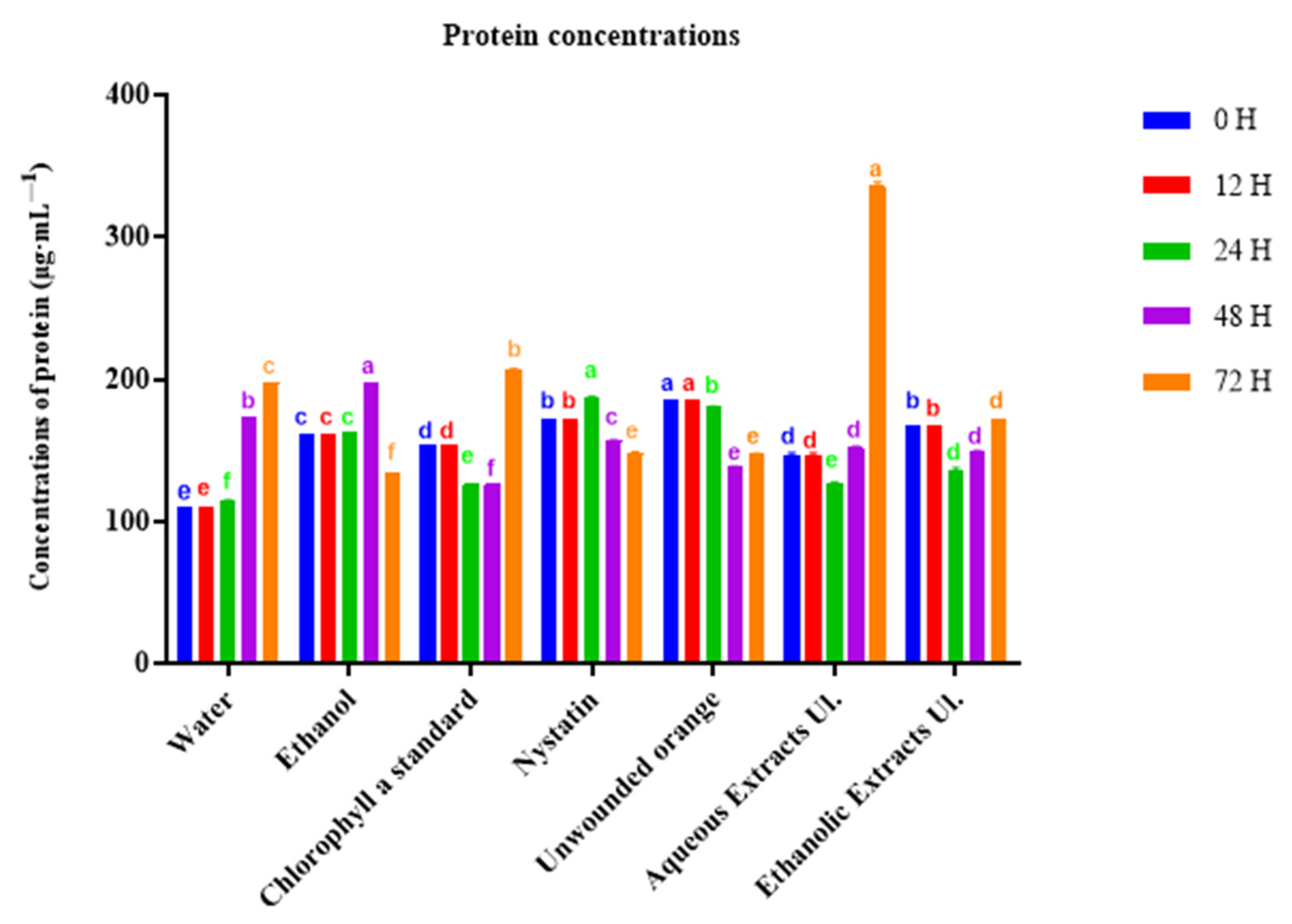
According to a global approach, we determined the concentrations of total proteins in inoculated tissues. Unwounded oranges and those that received nystatin exhibited a protein concentration diminution after incubation for 48 h (Figure 2). In contrast, oranges treated with chlorophyll a and aqueous extracts showed an increase in protein concentration at 72 h, reaching 337 µg·mL–1 with the aqueous extract.
3.3. Enzyme Activity Assays
3.3.1. β-1,3-glucanase Activity
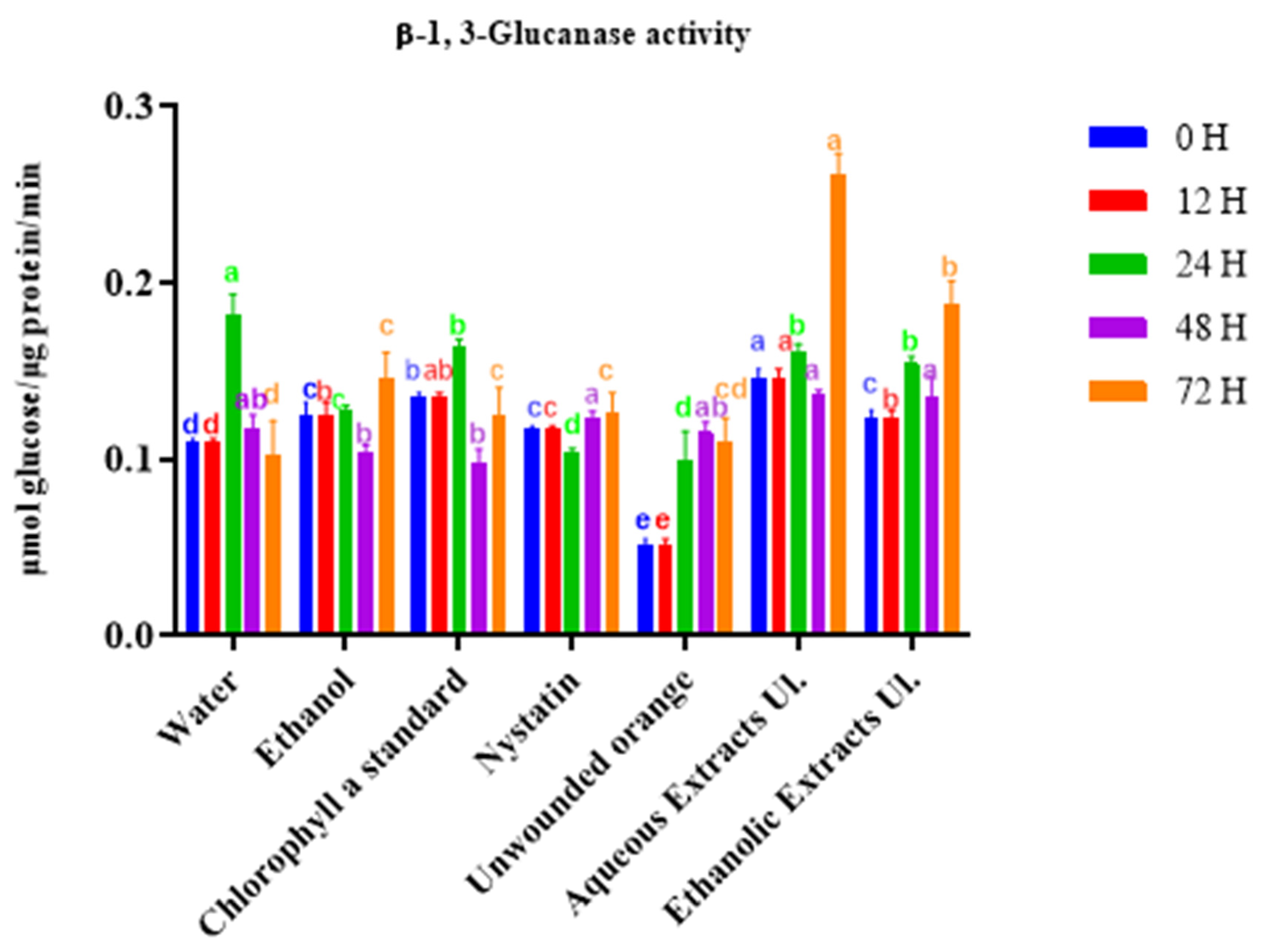
We next measured the kinetics of induction of β-1,3-glucanase activity in orange tissues. The ethanolic extract, and even more so the aqueous extract, induced β-1,3-glucanase activity relative to untreated oranges, reaching a maximum level above 0.18 μmol glucose·μg−1 protein·min−1 at 72 h (Figure 3). The contribution of chlorophyll a was mainly observed for 24 h, but remained moderate, indicating that other substances are involved in the enzyme activation.
3.3.2. Peroxidase Activity
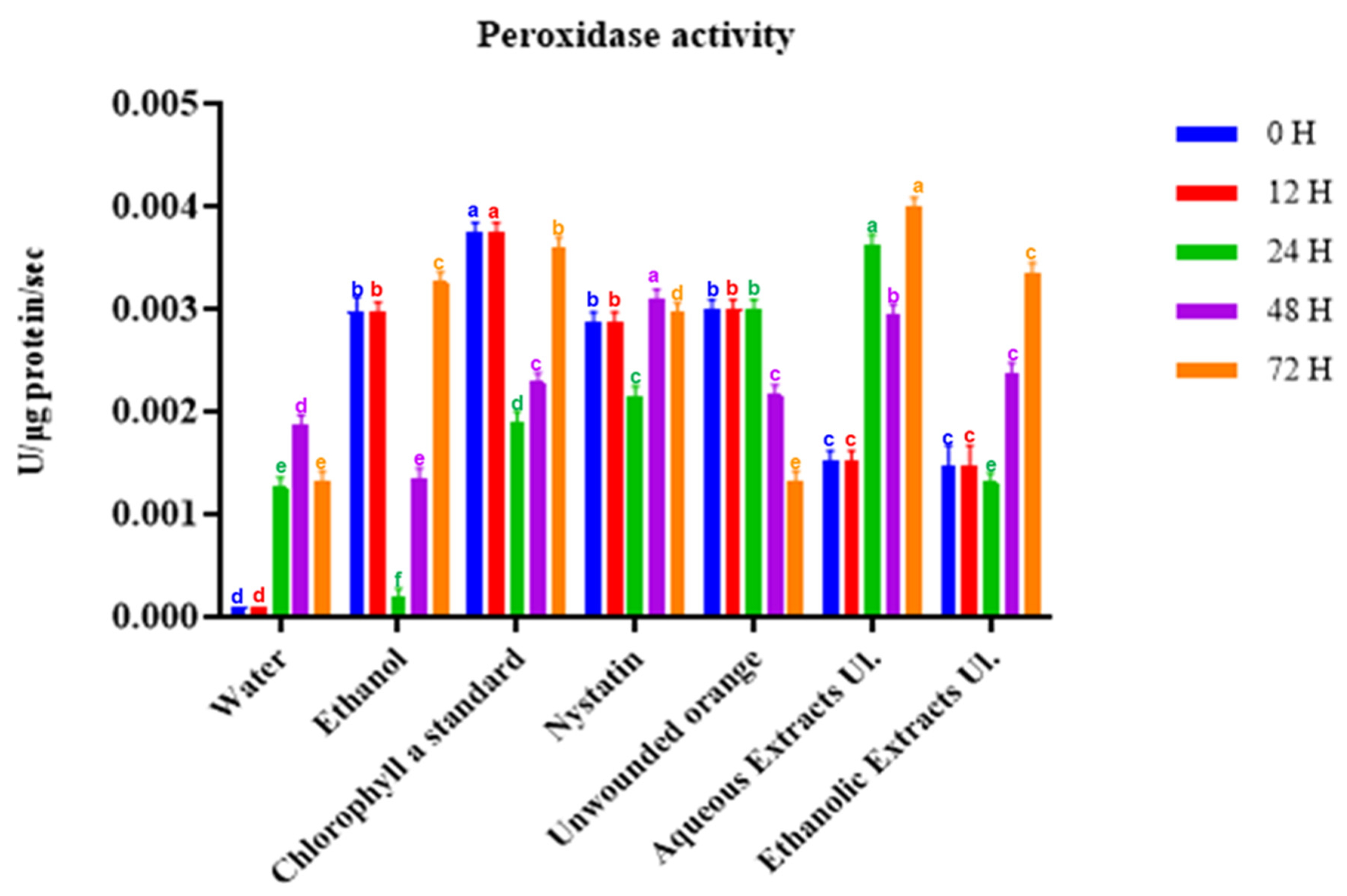
We also assessed the inducted kinetics of peroxidase activity in orange tissues (Figure 4). The enzymatic activity decreased for the unwounded oranges and remained stable for those treated with nystatin. Beyond 24 h, the ethanolic and aqueous extracts induced an increase in peroxidase activity (above 0.003 U·µg protein–1·s–1) in the treated tissues. These measured peroxidase activities were higher than the maximum value (0.0016 U·µg protein–1·s–1) observed after the treatment of citrus fruits with salt solutions [20].
During the incubation period, we detected an augmentation in peroxidase and β-1,3-glucanase activities in the orange tissues treated with aqueous and ethanolic extracts from U. lactuca. This suggested the role of enzyme activities in the beneficial effects observed after post-harvest treatment with the Ulva extracts. Chlorophyll a, ulvans and organic acids seem to be involved in the mechanism of resistance initiated in oranges. Llorens et al. [31] reported the role of polysaccharides, such as ulvans, in triggering defense responses in plants, enhancing their protection against pathogens. Moreover, Wang et al. [32] showed that organic acids such as succinic acid took part in the mechanism of defense response against rot in citrus fruit. Therefore, we can confirm that U. lactuca extracts contained active molecules able to initiate host defense mechanisms in oranges’ albedo.
Among the identified resistance inducers, some natural substances from green macroalgae have shown their potential for the development of alternative methods for the safe protection of fruit, as promoted by European directive 128/2009 and regulation 79/117/EEC.
4. Conclusions
The properties of Ulva lactuca extracts (both aqueous and ethanolic extracts), were investigated to identify their impacts on the control of green mold on citrus fruit. The measurement of disease incidence and lesion severity on treated wounded oranges highlighted some positive effects of U. lactuca liquid extracts against the studied pathogen. Moreover, an increase in enzyme activities in orange tissues was detected over 72 h following the treatment with these extracts. Several polar molecules such as chlorophyll a, polyphenols, organic acids and ulvans are likely responsible for these effects.
The direct inhibition of P. digitatum and the induction of fruit resistance represent two modes of action for Ulva lactuca extracts. On the basis of this activity, a new strategy of biocontrol could be a solution to preserve the harvested oranges without synthetic fungicides.
Author Contributions
Conceptualization: A.C. and P.d.C.; Methodology: D.S., J.A.-A., P.d.C. and A.C.; Formal analyses and investigation: D.S. and P.d.C.; Statistical analyses: D.S. and O.M.; Writing—preparation of the original draft: D.S., P.d.C. and O.M.; Review and editing: P.d.C. and O.M.; Project administration: P.d.C. and A.C. All authors have read and agreed to the published version of the manuscript.
Funding
A PhD fellowship for author DS was granted by the Laboratory of Applied Biotechnology (Azm Center) and the Doctoral School of Science and Technology (Lebanese University). The work was funded by Laboratoire de Chimie Agro-industrielle (LCA) in Toulouse INP (France).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caserta, R.; Teixeira-Silva, N.S.; Granato, L.M.; Dorta, S.O.; Rodrigues, C.M.; Mitre, L.K. Citrus biotechnology: What has been done to improve disease resistance in such an important crop? Biotechnol. Res. Innov. 2020, 3, 95–109. [Google Scholar] [CrossRef]
- Dillard, C.J.; German, J.B. Phytochemicals: Nutraceuticals and human health. J. Sci. Food Agric. 2000, 80, 1744–1756. [Google Scholar] [CrossRef]
- Kanetis, L.; Förster, H.; Adaskaveg, J.E. Comparative efficacy of the new postharvest fungicides azoxystrobin, fludioxonil and pyrimethanil for managing citrus green mold. Plant Dis. 2017, 91, 1502–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macarisin, D.; Cohen, L.; Eick, A.; Rafael, G.; Belausov, E.; Wisniewski, M.; Droby, S. Penicillium digitatum suppresses production of hydrogen peroxide in host tissue during infection of citrus fruit. Phytopathology 2007, 97, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Palou, L.; Smilanick, J.L.; Droby, S. Alternatives to conventional fungicides for the control of citrus postharvest green and blue moulds. Stewart Postharvest Rev. 2008, 2, 1–16. [Google Scholar]
- Moraes Bazioli, J.; Belinato, J.R.; Costa, J.H.; Akiyama, D.Y.; de Moraes Pontes, J.-G.; Kupper, K.C.; Augusto, F.; de Carvalho, J.E.; Fill, T.P. Biological Control of Citrus Postharvest Phytopathogens. Toxins 2019, 11, 460. [Google Scholar] [CrossRef] [Green Version]
- Kharchoufi, S.; Parafati, L.; Licciardello, F.; Muratore, G.; Hamdi, M.; Cirvilleri, G.; Restuccia, C. Edible coatings incorporating pomegranate peel extract and biocontrol yeast to reduce Penicillium digitatum postharvest decay of oranges. Food Microbiol. 2018, 74, 107–112. [Google Scholar] [CrossRef]
- Li, J.; Li, H.; Ji, S.; Chen, T.; Tian, S.; Qin, G. Enhancement of biocontrol efficacy of Cryptococcus laurentii by cinnamic acid against Penicillium italicum in citrus fruit. Postharvest Biol. Technol. 2019, 149, 42–49. [Google Scholar] [CrossRef]
- Carmona-Hernandez, S.; Reyes-Pérez, J.J.; Chiquito-Contreras, G.; Rincon-Enriquez, G.; Cerdan-Cabrera, C.R.; Hernandez-Montiel, L.G. Biocontrol of postharvest fruit fungal diseases by bacterial antagonists: A review. Agronomy 2019, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. The effect of locust bean gum (LBG)-based edible coatings carrying biocontrol yeasts against Penicillium digitatum and Penicillium italicum causal agents of postharvest decay of mandarin fruit. Food Microbiol. 2016, 58, 87–94. [Google Scholar] [CrossRef]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials—A review. Plants 2016, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Hammet, S.M.; El-Rhman, A.A.; Abdel-Raouf, N.; Ibraheem, I.B.M. Role of marine macroalgae in plant protection & improvement for sustainable agriculture technology. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 104–110. [Google Scholar] [CrossRef]
- Suthin Raj, T.; Vignesh, S.; Nishanthi, P.; Hane graff, K.; Ann Suji, H. Induction of defence enzymes activities in grape plant treated by seaweed algae against Plasmopara viticola and Uncinula necator causing downy and powdery mildews of grapes. Nov. Res. Microbiol. J. 2018, 2, 122–137. [Google Scholar] [CrossRef] [Green Version]
- Salim, D.; de Caro, P.; Merah, O.; Chbani, A. Control of post-harvest citrus green mold using Ulva lactuca extracts as a source of active substances. Int. J. Biores. Stress Manag. 2020, 11, 287–296. [Google Scholar] [CrossRef]
- Salim, D.; de Caro, P.; Chasseray, X.; Sing, A.S.C. Development of biobased emulsions for postharvest citrus fruit preservation. Sustain. Chem. Pharm. 2022, 25, 100583. [Google Scholar] [CrossRef]
- Romanazzi, G.; Feliziani, E.; Santini, M.; Landi, L. Effectiveness of postharvest treatment with chitosan and other resistance inducers in the control of storage decay of strawberry. Postharvest Biol. Technol. 2013, 75, 24–27. [Google Scholar] [CrossRef]
- Asimakis, E.; Shehata, A.A.; Eisenreich, W.; Acheuk, F. Algae and their metabolites as potential bio-pesticides. Microorganisms 2022, 10, 307. [Google Scholar] [CrossRef]
- Costa, J.H.; Bazioli, J.M.; de Moraes Pontes, J.G.; Fill, T.P. Penicillium digitatum infection mechanisms in citrus: What do we know so far? Fungal. Biol. 2019, 123, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.R.; Lafuente, M.T.; González-Candelas, L. Spatial study of antioxidant enzymes, peroxidase and phenylalanine ammonia-lyase in the citrus fruit–Penicillium digitatum interaction. Postharvest Biol. Technol. 2010, 56, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Youssef, K.; Sanzani, S.M.; Ligorio, A.; Ippolito, A.; Terry, L.A. Sodium carbonate and bicarbonate treatments induce resistance to postharvest green mould on citrus fruit. Postharvest Biol. Technol. 2014, 87, 61–69. [Google Scholar] [CrossRef]
- Fallanaj, F.; Ippolito, A.; Ligorio, A.; Garganese, F.; Zavanella, C.; Sanzani, S.M. Electrolyzed sodium bicarbonate inhibits Penicillium digitatum and induces defence responses against green mould in citrus fruit. Postharvest Biol. Technol. 2016, 115, 18–29. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, F.; Lu, Y.; Deng, J. Combination of chitosan and salicylic acid to control postharvest green mold caused by Penicillium digitatum in grapefruit fruit. Sci. Hortic. 2018, 233, 54–60. [Google Scholar] [CrossRef]
- Abouraïcha, E.; El Alaoui-Talibi, Z.; El Boutachfaiti, R.; Petit, E.; Courtois, B.; Courtois, J.; El Modafar, C. Induction of natural defense and protection against Penicillium expansum and Botrytis cinerea in apple fruit in response to bioelicitors isolated from green algae. Sci. Hortic. 2015, 181, 121–128. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford Method for Protein Quantitation. In The Protein Protocols Handbook, 2nd ed.; Walker, J.M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2002; pp. 15–21. [Google Scholar]
- Abeles, F.B.; Forrence, L.E. Temporal and hormonal control of β-1,3-glucanase in Phaseolus Vulgaris L. Plant Physiol. 1979, 45, 395–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer-Schmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Bhagavathy, S.; Sumathi, P.; Jancy Sherene Bell, I. Green algae Chlorococcum humicola-a new source of bioactive compounds with antimicrobial activity. Asian Pac. J. Trop. Biomed. 2011, 1, S1–S7. [Google Scholar] [CrossRef]
- Li Destri Nicosia, G.M.; Pangallo, S.; Raphael, G.; Romeo, F.V.; Strano, M.C.; Rapisarda, P.; Droby, S.; Schena, L. Control of postharvest fungal rots on citrus fruit and sweet cherries using a pomegranate peel extract. Postharvest Biol. Technol. 2016, 114, 54–61. [Google Scholar] [CrossRef]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Martínez, P.G.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Papoutsis, K.; Mathioudakis, M.M.; Hasperué, J.H.; Ziogas, V. Non-chemical treatments for preventing the postharvest fungal rotting of citrus caused by Penicillium digitatum (green mold) and Penicillium italicum (blue mold). Trends Food Sci. Technol. 2019, 86, 479–491. [Google Scholar] [CrossRef]
- Llorens, E.; García-Agustín, P.; Lapeña, I. Advances in induced resistance by natural compounds: Towards new options for woody plant protection. Sci. Agric. 2018, 74, 90–100. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Y.; Luo, W.; Deng, L.; Yao, S.; Zeng, K. Primary metabolites analysis of induced citrus fruit disease resistance upon treatment with oligochitosan, salicylic acid and Pichia membranaefaciens. Biol. Control. 2020, 148, 104289. [Google Scholar] [CrossRef]
Figure 1.
Disease incidence and severity (lesion diameter) caused by P. digitatum (suspension of 104 spores·mL–1) for oranges treated with aqueous and ethanolic extracts of Ulva lactuca and with an ethanolic solution of chlorophyll a, 14 days after inoculation. Columns (means) with different letters are significantly different: p < 0.05 (Duncan’s test).
Figure 1.
Disease incidence and severity (lesion diameter) caused by P. digitatum (suspension of 104 spores·mL–1) for oranges treated with aqueous and ethanolic extracts of Ulva lactuca and with an ethanolic solution of chlorophyll a, 14 days after inoculation. Columns (means) with different letters are significantly different: p < 0.05 (Duncan’s test).

Figure 2.
Protein concentration (µg·mL−1) of “Valencia late” oranges treated with the aqueous or ethanolic extract of Ulva lactuca (Ul.) measured during incubation. For each time point, columns (treatments) with different letters are significantly different: p < 0.05 (Duncan’s test).
Figure 2.
Protein concentration (µg·mL−1) of “Valencia late” oranges treated with the aqueous or ethanolic extract of Ulva lactuca (Ul.) measured during incubation. For each time point, columns (treatments) with different letters are significantly different: p < 0.05 (Duncan’s test).

Figure 3.
Enzymatic activity of β-1,3-glucanase in orange tissues treated with ethanolic or aqueous extracts of Ulva lactuca. Tissues were sampled up to 72 h. For each timepoint, columns (treatments) with different letters are significantly different: p < 0.05 (Duncan’s test).
Figure 3.
Enzymatic activity of β-1,3-glucanase in orange tissues treated with ethanolic or aqueous extracts of Ulva lactuca. Tissues were sampled up to 72 h. For each timepoint, columns (treatments) with different letters are significantly different: p < 0.05 (Duncan’s test).

Figure 4.
Enzymatic activity of peroxidase in orange tissues treated with ethanolic or aqueous extracts and chlorophyll a of U. lactuca. Tissues were sampled for up to 72 h. For each timepoint, columns (treatments) with different letters are significantly different: p < 0.05 (Duncan’s test).
Figure 4.
Enzymatic activity of peroxidase in orange tissues treated with ethanolic or aqueous extracts and chlorophyll a of U. lactuca. Tissues were sampled for up to 72 h. For each timepoint, columns (treatments) with different letters are significantly different: p < 0.05 (Duncan’s test).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Designed samples used for treatment with seaweed liquid extracts (SLEs).
| Treatment (Liquid Extracts) | Total Polyphenol Content (mg GAE.g–1 Dry Extract) * | |
|---|---|---|
| Chlorophyll a standard | - | 0.5 mg·mL–1 ethanol |
| U. l. aqueous extract: 12 mg DM·mL–1 | 0.45 ± 0.01 | 0.5 ± 0.1 mg·g–1 DE |
| U. l. ethanolic extract: 2.9 mg DM·mL–1 | 1.66 ± 0.05 | 1.1 ± 0.1 mg·g–1 DE |
| Unwounded fruit: untreated |
DM: dry matter in liquid extract, DE: dry extract; * according to the Folin–Ciocalteu method (λ = 760 nm), expressed in grams of gallic acid equivalents per gram of extract on a dry weight basis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Salim, D.; Al-Alam, J.; Merah, O.; Chbani, A.; de Caro, P. Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges. Phycology 2023, 3, 202-210. https://doi.org/10.3390/phycology3010013
AMA Style
Salim D, Al-Alam J, Merah O, Chbani A, de Caro P. Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges. Phycology. 2023; 3(1):202-210. https://doi.org/10.3390/phycology3010013
Chicago/Turabian StyleSalim, Douaa, Josephine Al-Alam, Othmane Merah, Asma Chbani, and Pascale de Caro. 2023. "Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges" Phycology 3, no. 1: 202-210. https://doi.org/10.3390/phycology3010013