
Spatial Computational Hepatic Molecular Biomarker Reveals LSEC Role in Midlobular Liver Zonation Fibrosis in DILI and NASH Liver Injury

Abstract
:1. Introduction
2. Methods and Materials
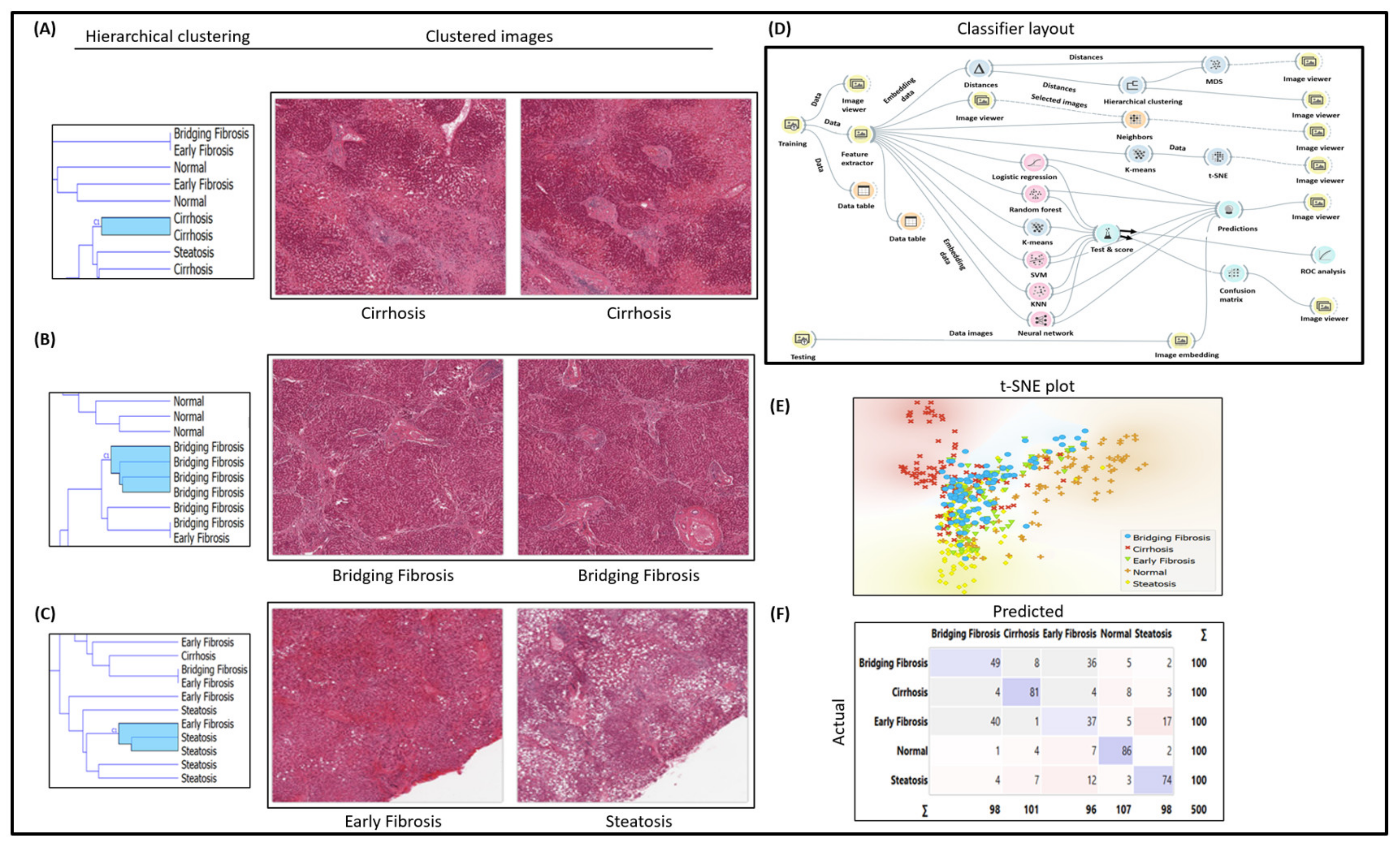
2.1. H&E Histopathology Image Classifications
2.2. Spatial Transcriptomics Data Analysis
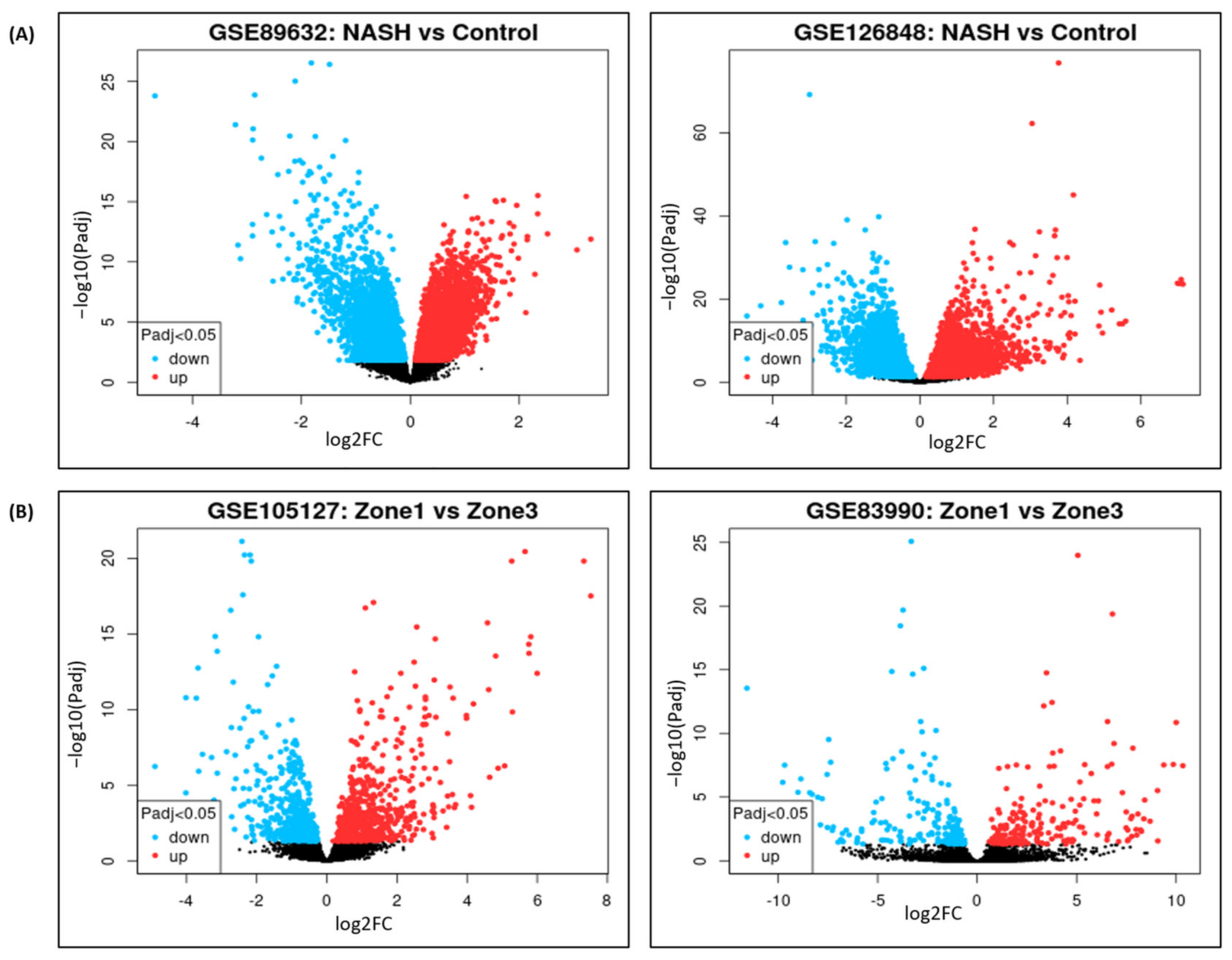
2.2.1. DGE Analysis for NASH and Zonation
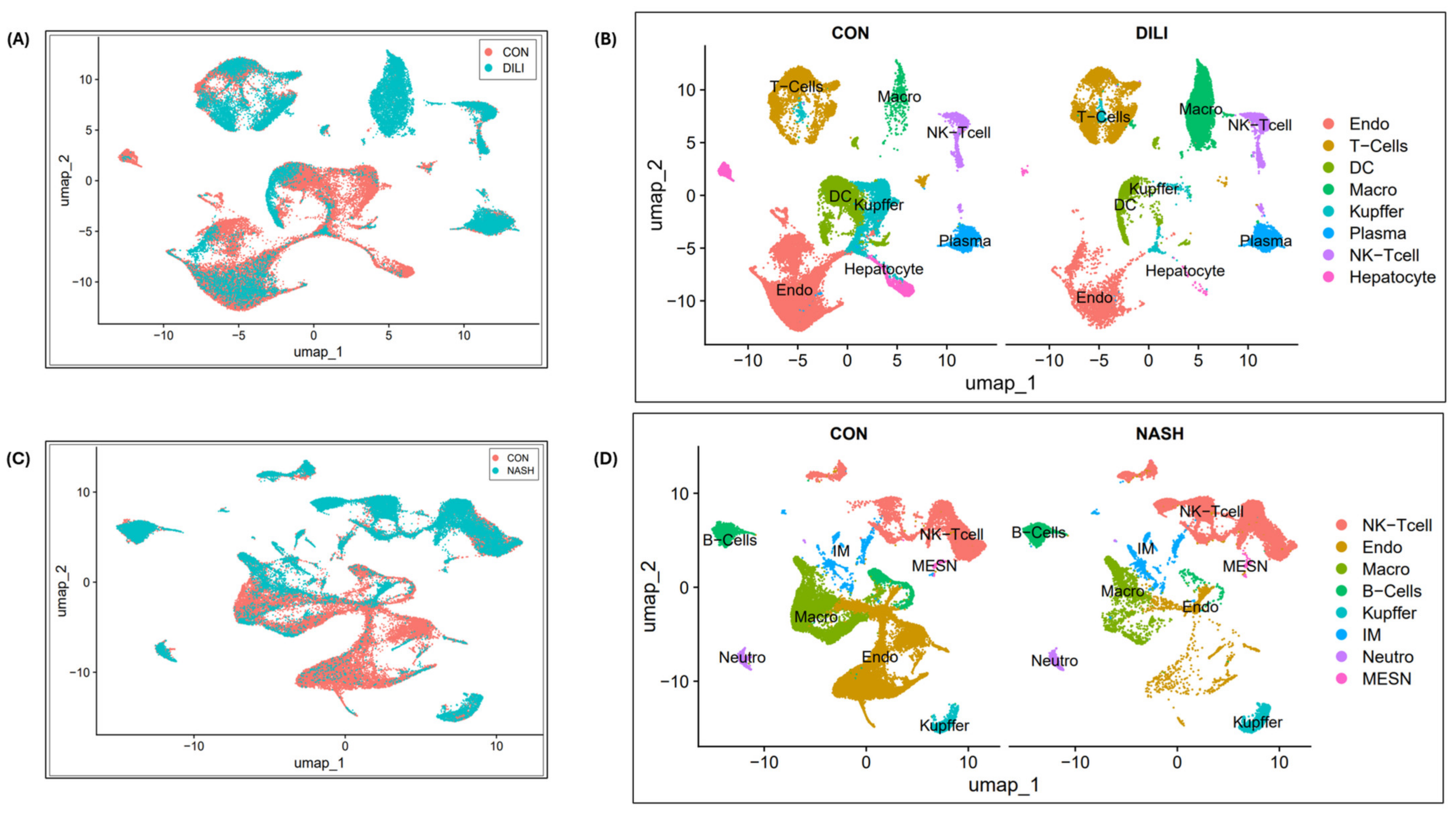
2.2.2. Single-Cell Clustering for NASH and DILI
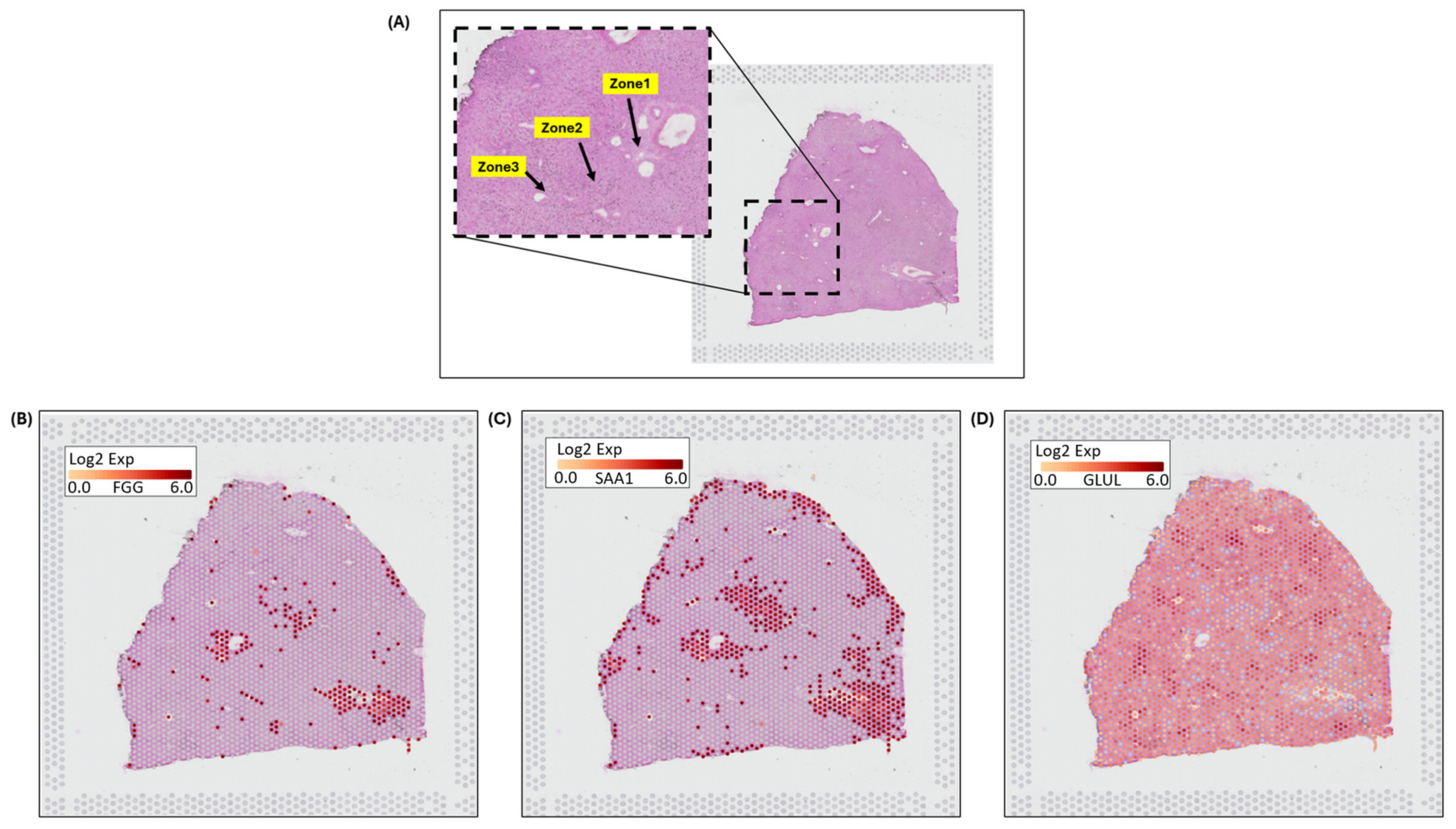
2.2.3. Visium Data Analysis and Visualization
2.2.4. Spatial Molecular Imaging
2.3. Liver Cell Clustering and Analysis Tool
2.4. LSEC Markers
3. Results
3.1. Histopathology H&E Image Classification Experimental Workflow for Early and Bridging Fibrosis
3.2. DGE Analysis for NASH and Zonation Expression
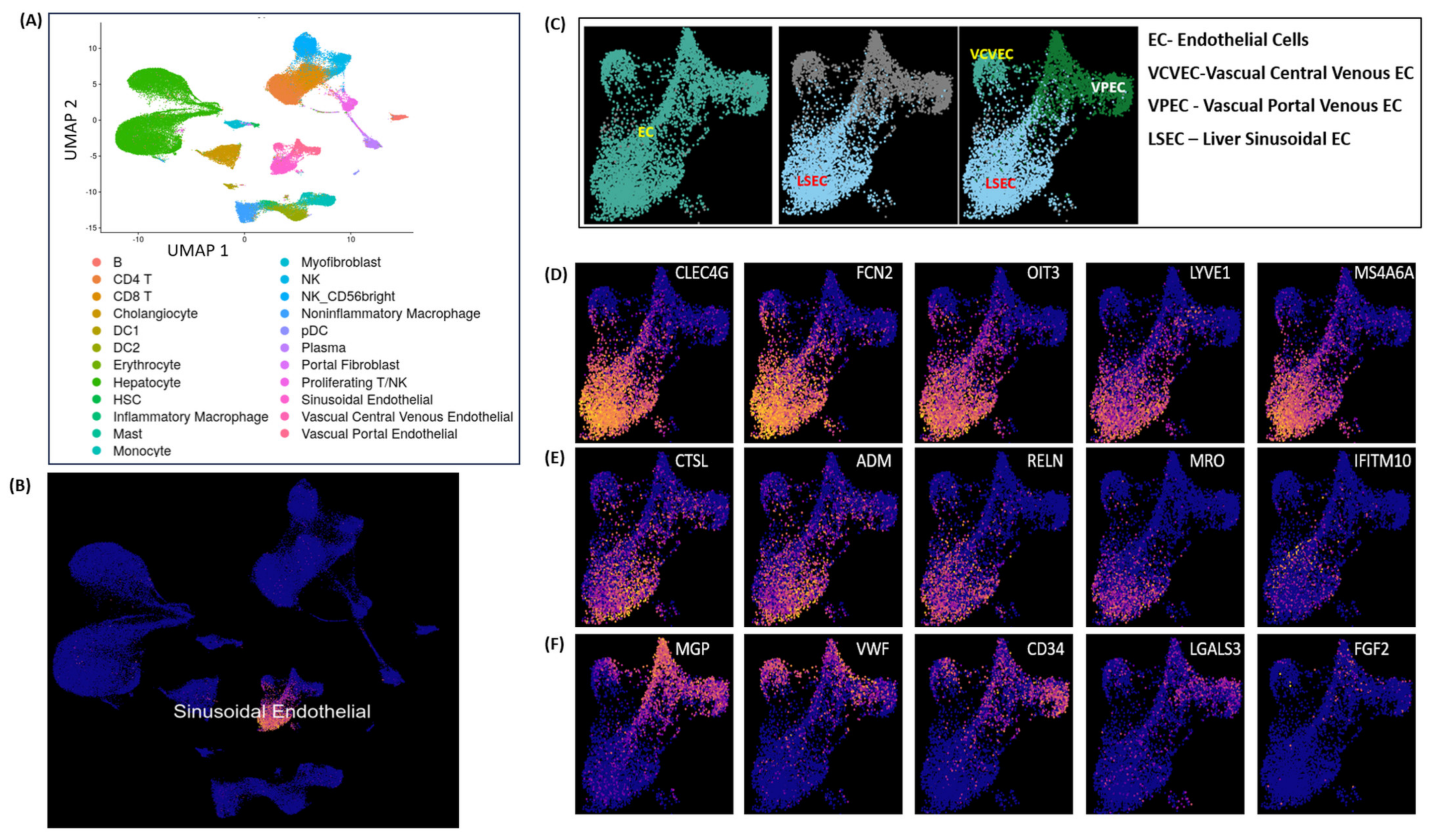
3.3. Single-Cell Clustering and DGE Expression Profiles in DILI and NASH ECs
3.4. Spatial Transcriptomics Data Analysis for Zonation Marker Genes
3.4.1. 10x Genomics Visium Image Analysis for Spatial Distribution of Zonation Expression Markers
3.4.2. Spatial Molecular Imaging to Demonstrate Liver Zonation Architecture
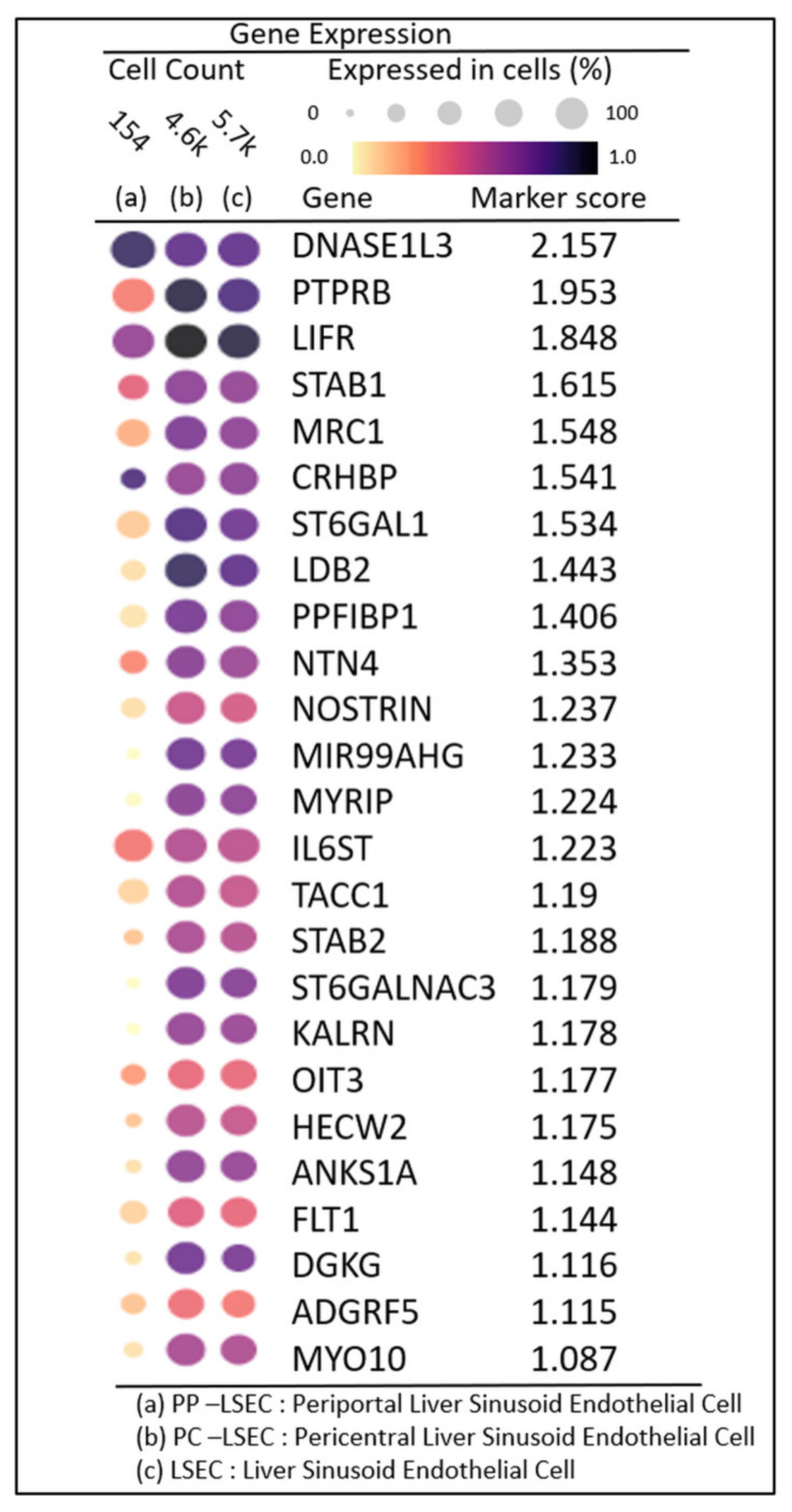
3.5. Liver Cell Clustering and LSEC Markers
3.6. DGE Profiles in Zonated LSECs
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global Burden of Liver Disease: 2023 Update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef]
- Ben-Moshe, S.; Itzkovitz, S. Spatial Heterogeneity in the Mammalian Liver. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T. The Truth Lies Somewhere in the Middle: The Cells Responsible for Liver Tissue Maintenance Finally Identified. Cell Regen. 2021, 10, 28. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, Y.G.; Jia, Y.; Li, L.; Yoon, J.; Zhang, S.; Wang, Z.; Zhang, Y.; Zhu, M.; Sharma, T.; et al. Liver Homeostasis Is Maintained by Midlobular Zone 2 Hepatocytes. Science 2021, 371, eabb1625. [Google Scholar] [CrossRef] [PubMed]
- Panday, R.; Monckton, C.P.; Khetani, S.R. The Role of Liver Zonation in Physiology, Regeneration, and Disease. Semin. Liver Dis. 2022, 42, 001–016. [Google Scholar] [CrossRef] [PubMed]
- Verhulst, S.; van Os, E.A.; De Smet, V.; Eysackers, N.; Mannaerts, I.; van Grunsven, L.A. Gene Signatures Detect Damaged Liver Sinusoidal Endothelial Cells in Chronic Liver Diseases. Front. Med. 2021, 8, 750044. [Google Scholar] [CrossRef]
- Su, T.; Yang, Y.; Lai, S.; Jeong, J.; Jung, Y.; McConnell, M.; Utsumi, T.; Iwakiri, Y. Single-Cell Transcriptomics Reveals Zone-Specific Alterations of Liver Sinusoidal Endothelial Cells in Cirrhosis. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1139–1161. [Google Scholar] [CrossRef]
- DeLeve, L.D. Liver Sinusoidal Endothelial Cells in Hepatic Fibrosis. Hepatology 2015, 61, 1740–1746. [Google Scholar] [CrossRef]
- Lafoz, E.; Ruart, M.; Anton, A.; Oncins, A.; Hernández-Gea, V. The Endothelium as a Driver of Liver Fibrosis and Regeneration. Cells 2020, 9, 929. [Google Scholar] [CrossRef]
- Gao, J.; Zuo, B.; He, Y. Liver Sinusoidal Endothelial Cells as Potential Drivers of Liver Fibrosis (Review). Mol. Med. Rep. 2024, 29, 40. [Google Scholar] [CrossRef]
- Velliou, R.-I.; Legaki, A.-I.; Nikolakopoulou, P.; Vlachogiannis, N.I.; Chatzigeorgiou, A. Liver Endothelial Cells in NAFLD and Transition to NASH and HCC. Cell. Mol. Life Sci. 2023, 80, 314. [Google Scholar] [CrossRef]
- Nasiri-Ansari, N.; Androutsakos, T.; Flessa, C.-M.; Kyrou, I.; Siasos, G.; Randeva, H.S.; Kassi, E.; Papavassiliou, A.G. Endothelial Cell Dysfunction and Nonalcoholic Fatty Liver Disease (NAFLD): A Concise Review. Cells 2022, 11, 2511. [Google Scholar] [CrossRef]
- Ghallab, A.; Myllys, M.; Holland, C.H.; Zaza, A.; Murad, W.; Hassan, R.; Ahmed, Y.A.; Abbas, T.; Abdelrahim, E.A.; Schneider, K.M.; et al. Influence of Liver Fibrosis on Lobular Zonation. Cells 2019, 8, 1556. [Google Scholar] [CrossRef]
- Nagy, D.; Maude, H.; Birdsey, G.M.; Randi, A.M.; Cebola, I. RISING STARS: Liver Sinusoidal Endothelial Transcription Factors in Metabolic Homeostasis and Disease. J. Mol. Endocrinol. 2023, 71, e230026. [Google Scholar] [CrossRef]
- Ben-Moshe, S.; Veg, T.; Manco, R.; Dan, S.; Papinutti, D.; Lifshitz, A.; Kolodziejczyk, A.A.; Halpern, K.B.; Elinav, E.; Itzkovitz, S. The Spatiotemporal Program of Zonal Liver Regeneration Following Acute Injury. Cell Stem Cell 2022, 29, 973–989.e10. [Google Scholar] [CrossRef]
- Demšar, J.; Curk, T.; Erjavec, A.; Gorup, Č.; Hočevar, T.; Milutinovič, M.; Možina, M.; Polajnar, M.; Toplak, M.; Starič, A.; et al. Orange: Data Mining Toolbox in Python. J. Mach. Learn. Res. 2013, 14, 2349–2353. [Google Scholar]
- Godec, P.; Pančur, M.; Ilenič, N.; Čopar, A.; Stražar, M.; Erjavec, A.; Pretnar, A.; Demšar, J.; Starič, A.; Toplak, M.; et al. Democratized Image Analytics by Visual Programming through Integration of Deep Models and Small-Scale Machine Learning. Nat. Commun. 2019, 10, 4551. [Google Scholar] [CrossRef]
- Wang, Z.; Qian, J.; Lu, X.; Zhang, P.; Guo, R.; Lou, H.; Zhang, S.; Yang, J.; Fan, X. A Single-Cell Transcriptomic Atlas Characterizes Liver Non-Parenchymal Cells in Healthy and Diseased Mice. bioRxiv 2021. [Google Scholar] [CrossRef]
- MacParland, S.A.; Liu, J.C.; Ma, X.-Z.; Innes, B.T.; Bartczak, A.M.; Gage, B.K.; Manuel, J.; Khuu, N.; Echeverri, J.; Linares, I.; et al. Single Cell RNA Sequencing of Human Liver Reveals Distinct Intrahepatic Macrophage Populations. Nat. Commun. 2018, 9, 4383. [Google Scholar] [CrossRef]
- Ramachandran, P.; Dobie, R.; Wilson-Kanamori, J.R.; Dora, E.F.; Henderson, B.E.P.; Luu, N.T.; Portman, J.R.; Matchett, K.P.; Brice, M.; Marwick, J.A.; et al. Resolving the Fibrotic Niche of Human Liver Cirrhosis at Single-Cell Level. Nature 2019, 575, 512–518. [Google Scholar] [CrossRef]
- Aizarani, N.; Saviano, A.; Sagar, N.; Mailly, L.; Durand, S.; Herman, J.S.; Pessaux, P.; Baumert, T.F.; Grün, D. A Human Liver Cell Atlas Reveals Heterogeneity and Epithelial Progenitors. Nature 2019, 572, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M.; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated Analysis of Multimodal Single-Cell Data. Cell 2021, 184, 3573–3587.e29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, H.; Wan, L.; Wang, Z.; Wang, H.; Ge, C.; Liu, Y.; Hao, Y.; Zhang, D.; Shi, G.; et al. Single-Cell Transcriptomic Architecture and Intercellular Crosstalk of Human Intrahepatic Cholangiocarcinoma. J. Hepatol. 2020, 73, 1118–1130. [Google Scholar] [CrossRef] [PubMed]
- Brancale, J.; Vilarinho, S. A Single Cell Gene Expression Atlas of 28 Human Livers. J. Hepatol. 2021, 75, 219–220. [Google Scholar] [CrossRef] [PubMed]
- Halpern, K.B.; Shenhav, R.; Matcovitch-Natan, O.; Tóth, B.; Lemze, D.; Golan, M.; Massasa, E.E.; Baydatch, S.; Landen, S.; Moor, A.E.; et al. Single-Cell Spatial Reconstruction Reveals Global Division of Labour in the Mammalian Liver. Nature 2017, 542, 352–356. [Google Scholar] [CrossRef]
- Hildebrandt, F.; Andersson, A.; Saarenpää, S.; Larsson, L.; Van Hul, N.; Kanatani, S.; Masek, J.; Ellis, E.; Barragan, A.; Mollbrink, A.; et al. Spatial Transcriptomics to Define Transcriptional Patterns of Zonation and Structural Components in the Mouse Liver. Nat. Commun. 2021, 12, 7046. [Google Scholar] [CrossRef] [PubMed]
- Segal, J.M.; Kent, D.; Wesche, D.J.; Ng, S.S.; Serra, M.; Oulès, B.; Kar, G.; Emerton, G.; Blackford, S.J.I.; Darmanis, S.; et al. Single Cell Analysis of Human Foetal Liver Captures the Transcriptional Profile of Hepatobiliary Hybrid Progenitors. Nat. Commun. 2019, 10, 3350. [Google Scholar] [CrossRef] [PubMed]
- Program, C.S.-C.B.; Abdulla, S.; Aevermann, B.; Assis, P.; Badajoz, S.; Bell, S.M.; Bezzi, E.; Cakir, B.; Chaffer, J.; Chambers, S.; et al. CZ CELL×GENE Discover: A Single-Cell Data Platform for Scalable Exploration, Analysis and Modeling of Aggregated Data. bioRxiv 2023. [Google Scholar] [CrossRef]
- Massalha, H.; Bahar Halpern, K.; Abu-Gazala, S.; Jana, T.; Massasa, E.E.; Moor, A.E.; Buchauer, L.; Rozenberg, M.; Pikarsky, E.; Amit, I.; et al. A Single Cell Atlas of the Human Liver Tumor Microenvironment. Mol. Syst. Biol. 2020, 16, e9682. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, V.; Harris, E.N.; Kidambi, S. SECs (Sinusoidal Endothelial Cells), Liver Microenvironment, and Fibrosis. BioMed Res. Int. 2017, 2017, 4097205. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-W.; Ruan, B.; Yang, P.-J.; Liu, J.-J.; Song, P.; Duan, J.-L.; Wang, L. Oit3, a Promising Hallmark Gene for Targeting Liver Sinusoidal Endothelial Cells. Signal Transduct. Target. Ther. 2023, 8, 344. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.A.; Singer, H.; Al-Rifai, R.; Nüsgen, N.; Rath, M.; Strauss, S.; Andreou, I.; Oldenburg, J.; El-Maarri, O. Molecular Analysis of Fetal and Adult Primary Human Liver Sinusoidal Endothelial Cells: A Comparison to Other Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 7776. [Google Scholar] [CrossRef]
- Olsavszky, V.; Sticht, C.; Schmid, C.D.; Winkler, M.; Wohlfeil, S.A.; Olsavszky, A.; Schledzewski, K.; Géraud, C.; Goerdt, S.; Leibing, T.; et al. Exploring the Transcriptomic Network of Multi-Ligand Scavenger Receptor Stabilin-1- and Stabilin-2-Deficient Liver Sinusoidal Endothelial Cells. Gene 2021, 768, 145284. [Google Scholar] [CrossRef]
- Maeda-Smithies, N.; Hiller, S.; Dong, S.; Kim, H.-S.; Bennett, B.J.; Kayashima, Y. Ectopic Expression of the Stabilin2 Gene Triggered by an Intracisternal A Particle (IAP) Element in DBA/2J Strain of Mice. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 2020, 31, 2–16. [Google Scholar] [CrossRef]
- Huebert, R.C.; Jagavelu, K.; Hendrickson, H.I.; Vasdev, M.M.; Arab, J.P.; Splinter, P.L.; Trussoni, C.E.; LaRusso, N.F.; Shah, V.H. Aquaporin-1 Promotes Angiogenesis, Fibrosis, and Portal Hypertension Through Mechanisms Dependent on Osmotically Sensitive MicroRNAs. Am. J. Pathol. 2011, 179, 1851–1860. [Google Scholar] [CrossRef]
- Gerussi, A.; Natalini, A.; Antonangeli, F.; Mancuso, C.; Agostinetto, E.; Barisani, D.; Di Rosa, F.; Andrade, R.; Invernizzi, P. Immune-Mediated Drug-Induced Liver Injury: Immunogenetics and Experimental Models. Int. J. Mol. Sci. 2021, 22, 4557. [Google Scholar] [CrossRef] [PubMed]
- Puri, M. Automated Machine Learning Diagnostic Support System as a Computational Biomarker for Detecting Drug-Induced Liver Injury Patterns in Whole Slide Liver Pathology Images. Assay Drug Dev. Technol. 2020, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-C.; Chang, J.; Lee, H.S.; Kwon, H.J. Mitochondrial UQCRB as a New Molecular Prognostic Biomarker of Human Colorectal Cancer. Exp. Mol. Med. 2017, 49, e391. [Google Scholar] [CrossRef] [PubMed]
- Schwenger, K.J.; Clermont-Dejean, N.; Allard, J.P. The Role of the Gut Microbiome in Chronic Liver Disease: The Clinical Evidence Revised. JHEP Rep. 2019, 1, 214–226. [Google Scholar] [CrossRef]
- Tilg, H.; Cani, P.D.; Mayer, E.A. Gut Microbiome and Liver Diseases. Gut 2016, 65, 2035–2044. [Google Scholar] [CrossRef]
- Gurcan, M.N.; Boucheron, L.; Can, A.; Madabhushi, A.; Rajpoot, N.; Yener, B. Histopathological Image Analysis: A Review. IEEE Rev. Biomed. Eng. 2009, 2, 147. [Google Scholar] [CrossRef]
- Clifton, K.; Anant, M.; Aihara, G.; Atta, L.; Aimiuwu, O.K.; Kebschull, J.M.; Miller, M.I.; Tward, D.; Fan, J. STalign: Alignment of Spatial Transcriptomics Data Using Diffeomorphic Metric Mapping. Nat. Commun. 2023, 14, 8123. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | AUC | CA | F1 | Precision | Recall |
|---|---|---|---|---|---|
| KNN | 0.876 | 0.592 | 0.609 | 0.645 | 0.592 |
| Tree | 0.659 | 0.460 | 0.467 | 0.477 | 0.460 |
| SVM | 0.896 | 0.660 | 0.692 | 0.667 | 0.660 |
| Random Forest | 0.774 | 0.518 | 0.520 | 0.525 | 0.518 |
| Neural Network | 0.877 | 0.644 | 0.642 | 0.640 | 0.644 |
| Naïve Bayes | 0.847 | 0.600 | 0.597 | 0.598 | 0.600 |
| Logistic Regression | 0.892 | 0.656 | 0.655 | 0.654 | 0.656 |
| Constant | 0.500 | 0.200 | 0.067 | 0.040 | 0.582 |
| AdaBoost | 0.748 | 0.582 | 0.580 | 0.582 | 0.582 |
| NCBI GEO Study ID | Publication PMID | Year | Patients (N) | Differentially Expressed Genes | Top UP/Down Expressed Genes |
|---|---|---|---|---|---|
| GSE48452 | 23931760 | 2013 | NASH vs. Healthy Control(N = 46) | NASH vs. Cntrl = 47 Steatosis vs. Cntrl = 32 NASH vs. Steatosis = 1 | Up = H2AFY2, GALNT18; Down = APOF, C8B Up = RPS13, UBE2N; Down = CCDC82, NCAM2 Up = ZMAT3; Down = C8B |
| GSE89632 | 35166723 | 2016 | NASH vs. Healthy Control (N = 63) | NASH vs. Cntrl = 2641 Steatosis vs. Cntrl = 3627 NASH vs. Steatosis = 11 | Up = TYMS, FMO1; Down = MIR21, AXUD1 Up = FOSB, MYC; Down = RFXDC2, WNT5A Up = AKR1B10, CDC2; Down = IL6, CCL2 |
| GSE126848 | 30653341 | 2019 | NASH vs. Healthy Control (N = 45) | NASH vs. Cntrl = 1906 NAFLD vs. Cntrl = 1045 NASH vs. NAFLD = 5 | Up = UQCRBP1, SNORD140; Down = FNBP1, GLUD1P2 Up = FNBP1, GLUD1P2; Down = UQCRBP1, SNORD140 Up = MRC2, GALNT18; Down = ST3GAL6, MAT1A |
| GSE83990 | 29244788 | 2018 | Liver Zonation (N = 12) | Zone1 vs. Zone2 = 27 Zone2 vs. Zone 3 = 4 Zone1 vs. Zone 3 = 323 | Up = DPT, STAB1; Down = OAT, SLCO1B3 Up = HAL, OIT3; Down = GLUL, SRPX Up = HAL, AQP1; Down = OAT, CXCL6 |
| GSE105127 | 30297808 | 2018 | Liver Zonation (N = 57) | Zone1 vs. Zone2 = 63 Zone2 vs. Zone 3 = 37 Zone1 vs. Zone 3 = 1010 | Up = MGP, FGFR2; Down = TBX15, SLCO1B7 Up = SPRYD4, D9; Down = GLUL, PTGDS Up = KRT19, AQP1; Down = RSPO3, GLUL |
| zone 1 | zone 2 | zone 3 |
|---|---|---|
| MRC1 | DNASE1L3 | SELE |
| HAL | CRHBP | APOB |
| TIMP1 | C9 | GLUL |
| SERPINE1 | CDH5 | FGF2 |
| SAA1 | IGFBP7 | PLG |
| ID1 | APOF | ITGA5 |
| CLDN10 | C8B | ICOS |
| CRP | LYVE1 | PLPP3 |
| SLPI | NOSTRIN | CTSS |
| CHI3L1 | TTR | FGF2 |
| FST | BTNL9 | PLG |
| TIE1 | ENG | LGR5 |
| LGALS3 | LIFR | NOTUM |
| TRAT1 | FGG | SLC13A3 |
| SDS | TEK | OAT |
| PDPN | KRT7 | GPAM |
| ADAM23 | CXCL6 | SP5 |
| FGFR2 | LEPR | CYP2E1 |
| H2AFY2 | EDN1 | SLCO1B3 |
| RPL3 | CD34 | MTMR11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puri, M. Spatial Computational Hepatic Molecular Biomarker Reveals LSEC Role in Midlobular Liver Zonation Fibrosis in DILI and NASH Liver Injury. Int. J. Transl. Med. 2024, 4, 208-223. https://doi.org/10.3390/ijtm4020012
Puri M. Spatial Computational Hepatic Molecular Biomarker Reveals LSEC Role in Midlobular Liver Zonation Fibrosis in DILI and NASH Liver Injury. International Journal of Translational Medicine. 2024; 4(2):208-223. https://doi.org/10.3390/ijtm4020012
Chicago/Turabian StylePuri, Munish. 2024. "Spatial Computational Hepatic Molecular Biomarker Reveals LSEC Role in Midlobular Liver Zonation Fibrosis in DILI and NASH Liver Injury" International Journal of Translational Medicine 4, no. 2: 208-223. https://doi.org/10.3390/ijtm4020012