
Development of Mouse Hepatitis Virus Chimeric Reporter Viruses Expressing the 3CLpro Proteases of Human Coronaviruses HKU1 and OC43 Reveals Susceptibility to Inactivation by Natural Inhibitors Baicalin and Baicalein
 and
and
Abstract
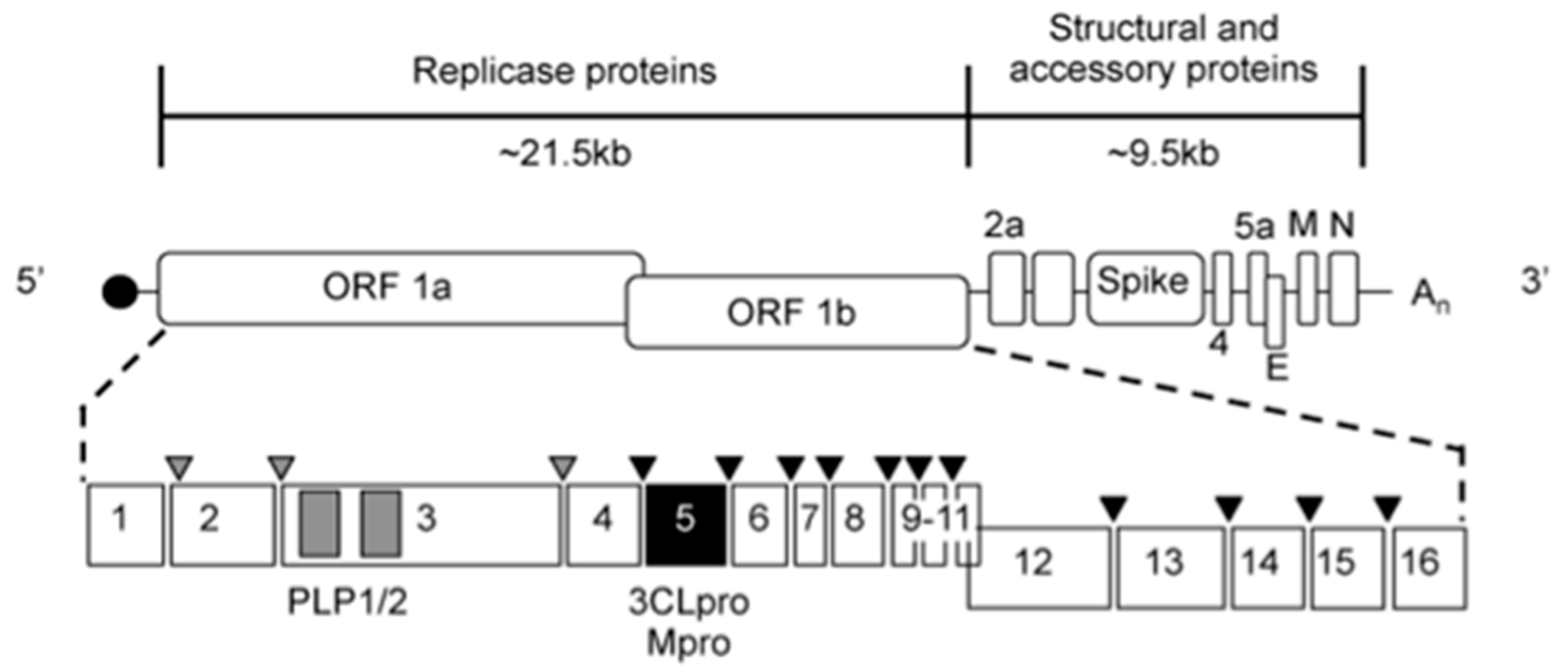
:1. Introduction
2. Materials and Methods
2.1. Viruses, Cells, and Inhibitors
2.2. Virus Recovery of H5-MHV-FFL and O5-MHV-FFL
2.3. RT-PCR and Sequencing
2.4. Viral Replication Assay and Luciferase Assay
2.5. Cytotoxicity Analysis
2.6. Inhibitor Assays
2.7. Analysis of Viral Escape
3. Results
3.1. H5-MHV and O5-MHV Chimeric Reporter Viruses Expressing Firefly Luciferase (FFL) Do Not Exhibit Altered Replication Kinetics
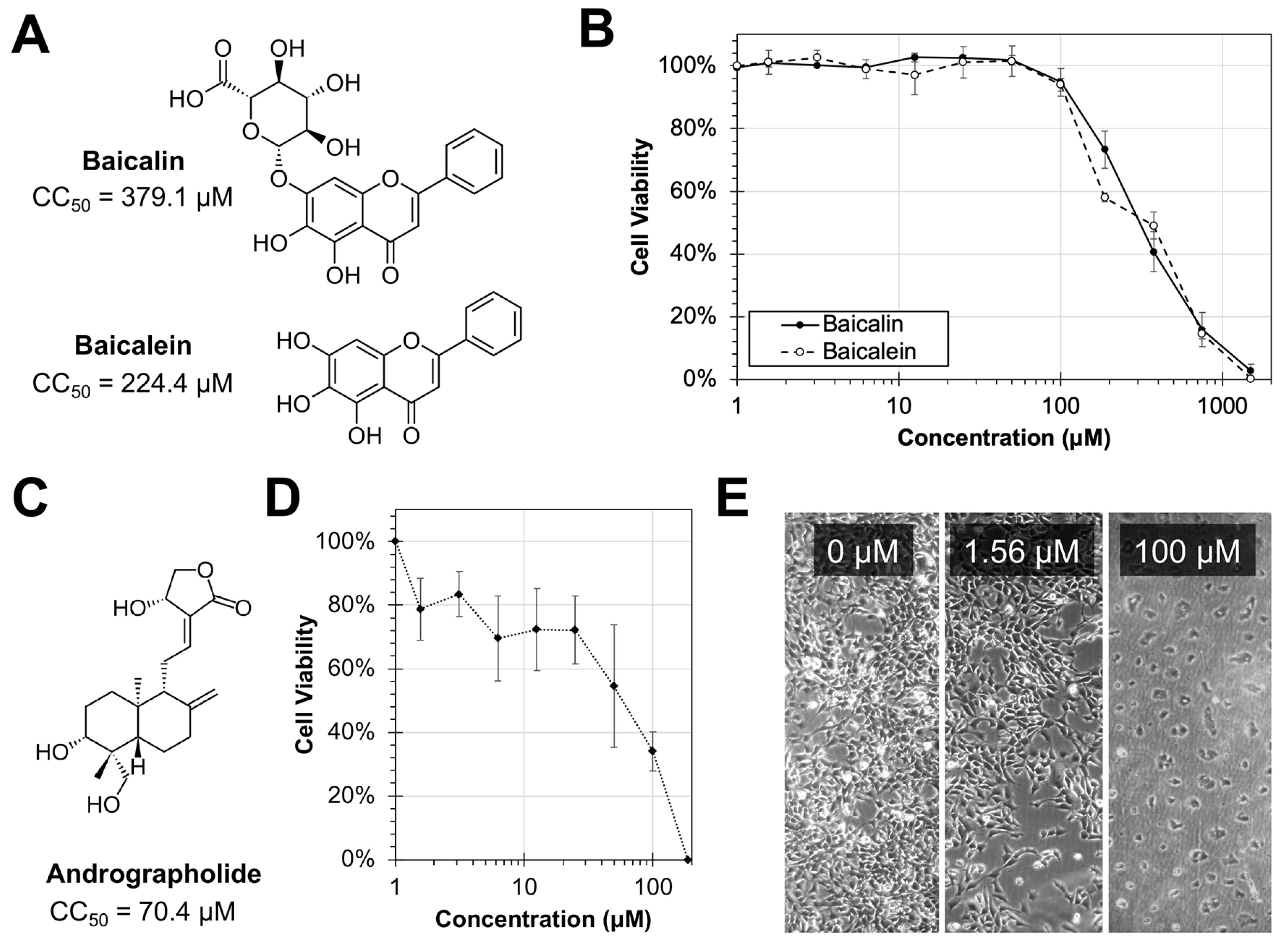
3.2. Analysis of Cytotoxicity of Natural Inhibitors of SARS-CoV-2 3CLpro
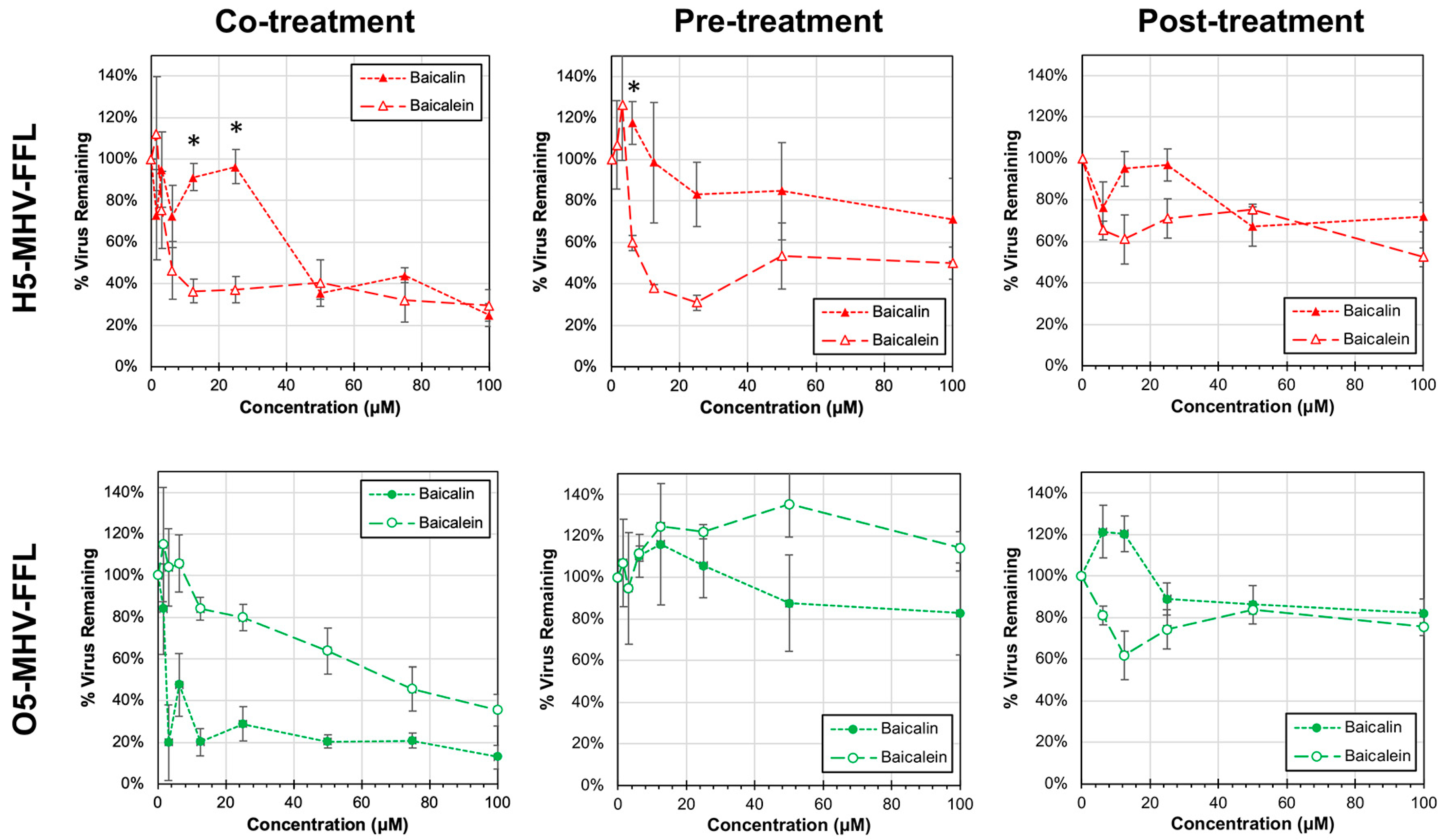
3.3. Antiviral Activity of Baicalin and Baicalein against H5-MHV-FFL and O5-MHV-FFL
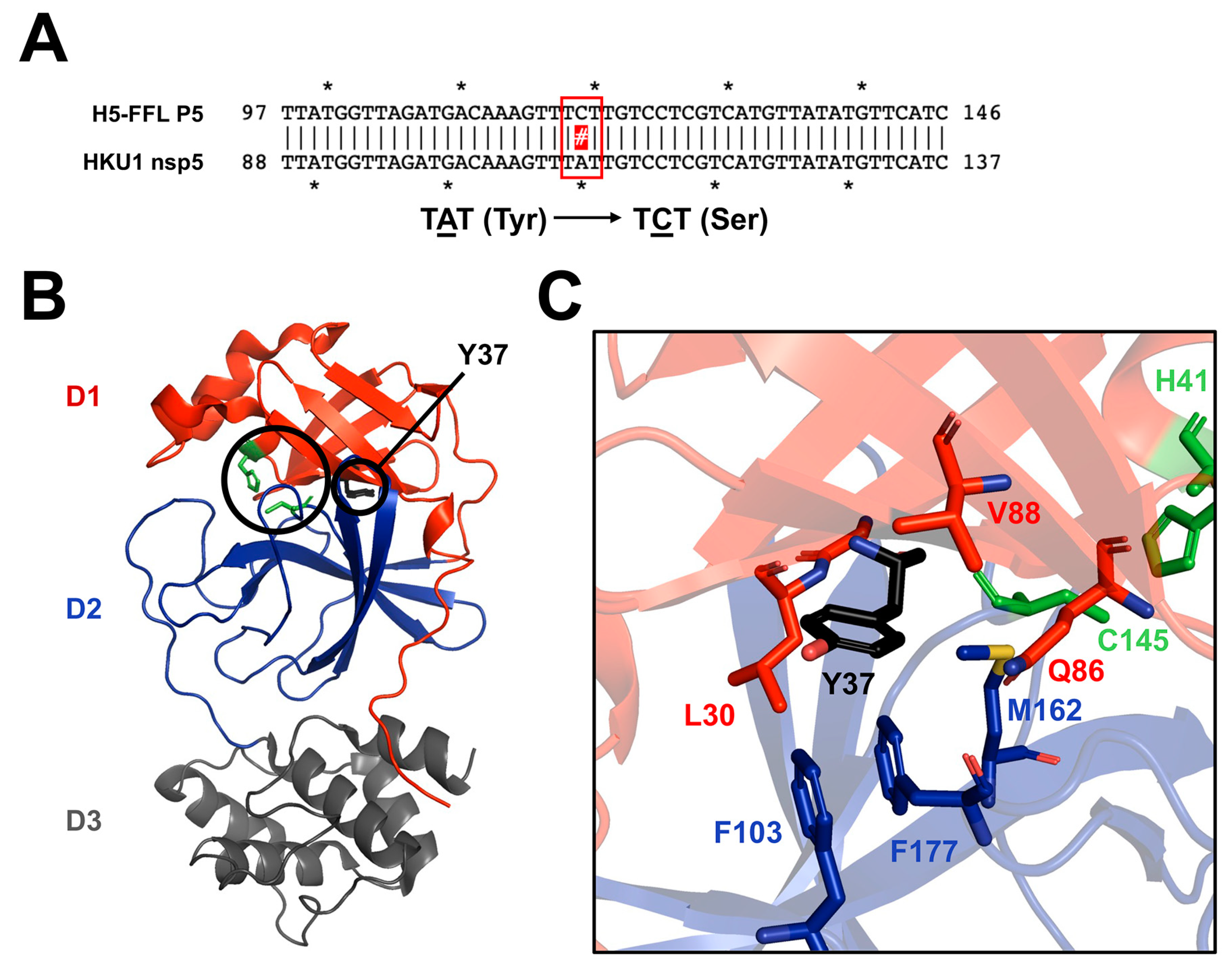
3.4. Identifying Potential Patterns of Viral Escape to Baicalein Treatment
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Grunewald, M.; Perlman, S. Coronaviruses: An Updated Overview of Their Replication and Pathogenesis. Methods Mol. Biol. Clifton NJ 2020, 2203, 1–29. [Google Scholar] [CrossRef]
- Harrison, C.M.; Doster, J.M.; Landwehr, E.H.; Kumar, N.P.; White, E.J.; Beachboard, D.C.; Stobart, C.C. Evaluating the Virology and Evolution of Seasonal Human Coronaviruses Associated with the Common Cold in the COVID-19 Era. Microorganisms 2023, 11, 445. [Google Scholar] [CrossRef]
- Drosten, C.; Günther, S.; Preiser, W.; van der Werf, S.; Brodt, H.-R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.M.; et al. Identification of a Novel Coronavirus in Patients with Severe Acute Respiratory Syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef]
- Perlman, S.; Netland, J. Coronaviruses Post-SARS: Update on Replication and Pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An Overview of Their Replication and Pathogenesis. Methods Mol. Biol. Clifton NJ 2015, 1282, 1–23. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Gong, T.; Wu, D.; Feng, Y.; Gao, Q.; Xing, J.; Zheng, X.; Song, Z.; Liu, X.; Chen, X.; et al. Isolation, Identification, and Pathogenicity of Porcine Epidemic Diarrhea Virus. Front. Microbiol. 2023, 14, 1273589. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Park, B. Porcine Epidemic Diarrhoea Virus: A Comprehensive Review of Molecular Epidemiology, Diagnosis, and Vaccines. Virus Genes 2012, 44, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Yap, C.; Ali, A.; Prabhakar, A.; Prabhakar, A.; Pal, A.; Lim, Y.Y.; Kakodkar, P. Comprehensive Literature Review on COVID-19 Vaccines and Role of SARS-CoV-2 Variants in the Pandemic. Ther. Adv. Vaccines Immunother. 2021, 9, 25151355211059791. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Rehman, S.U.; Yoo, H.H. COVID-19 Challenges and Its Therapeutics. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 142, 112015. [Google Scholar] [CrossRef]
- Ng, T.I.; Correia, I.; Seagal, J.; DeGoey, D.A.; Schrimpf, M.R.; Hardee, D.J.; Noey, E.L.; Kati, W.M. Antiviral Drug Discovery for the Treatment of COVID-19 Infections. Viruses 2022, 14, 961. [Google Scholar] [CrossRef]
- Temmam, S.; Vongphayloth, K.; Baquero, E.; Munier, S.; Bonomi, M.; Regnault, B.; Douangboubpha, B.; Karami, Y.; Chrétien, D.; Sanamxay, D.; et al. Bat Coronaviruses Related to SARS-CoV-2 and Infectious for Human Cells. Nature 2022, 604, 330–336. [Google Scholar] [CrossRef]
- Tao, Y.; Tang, K.; Shi, M.; Conrardy, C.; Li, K.S.M.; Lau, S.K.P.; Anderson, L.J.; Tong, S. Genomic Characterization of Seven Distinct Bat Coronaviruses in Kenya. Virus Res. 2012, 167, 67–73. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and Evolution of Pathogenic Coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef]
- Stobart, C.C.; Sexton, N.R.; Munjal, H.; Lu, X.; Molland, K.L.; Tomar, S.; Mesecar, A.D.; Denison, M.R. Chimeric Exchange of Coronavirus Nsp5 Proteases (3CLpro) Identifies Common and Divergent Regulatory Determinants of Protease Activity. J. Virol. 2013, 87, 12611–12618. [Google Scholar] [CrossRef]
- Anand, K.; Ziebuhr, J.; Wadhwani, P.; Mesters, J.R.; Hilgenfeld, R. Coronavirus Main Proteinase (3CLpro) Structure: Basis for Design of Anti-SARS Drugs. Science 2003, 300, 1763–1767. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, S.; Xue, F.; Zou, Y.; Chen, C.; Bartlam, M.; Rao, Z. Structure of the Main Protease from a Global Infectious Human Coronavirus, HCoV-HKU1. J. Virol. 2008, 82, 8647–8655. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal Structure of SARS-CoV-2 Main Protease Provides a Basis for Design of Improved α-Ketoamide Inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef]
- Ng, C.S.; Stobart, C.C.; Luo, H. Innate Immune Evasion Mediated by Picornaviral 3C Protease: Possible Lessons for Coronaviral 3C-like Protease? Rev. Med. Virol. 2021, 31, 1–22. [Google Scholar] [CrossRef]
- Stobart, C.C.; Lee, A.S.; Lu, X.; Denison, M.R. Temperature-Sensitive Mutants and Revertants in the Coronavirus Nonstructural Protein 5 Protease (3CLpro) Define Residues Involved in Long-Distance Communication and Regulation of Protease Activity. J. Virol. 2012, 86, 4801–4810. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Tiwari, V.; Sowdhamini, R. Computational Search for Potential COVID-19 Drugs from FDA-Approved Drugs and Small Molecules of Natural Origin Identifies Several Anti-Virals and Plant Products. J. Biosci. 2020, 45, 100. [Google Scholar] [CrossRef]
- Su, H.-X.; Yao, S.; Zhao, W.-F.; Li, M.-J.; Liu, J.; Shang, W.-J.; Xie, H.; Ke, C.-Q.; Hu, H.-C.; Gao, M.-N.; et al. Anti-SARS-CoV-2 Activities in Vitro of Shuanghuanglian Preparations and Bioactive Ingredients. Acta Pharmacol. Sin. 2020, 41, 1167–1177. [Google Scholar] [CrossRef]
- Enmozhi, S.K.; Raja, K.; Sebastine, I.; Joseph, J. Andrographolide as a Potential Inhibitor of SARS-CoV-2 Main Protease: An in Silico Approach. J. Biomol. Struct. Dyn. 2021, 39, 3092–3098. [Google Scholar] [CrossRef] [PubMed]
- Sa-Ngiamsuntorn, K.; Suksatu, A.; Pewkliang, Y.; Thongsri, P.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Pitiporn, S.; Chaopreecha, J.; et al. Anti-SARS-CoV-2 Activity of Andrographis Paniculata Extract and Its Major Component Andrographolide in Human Lung Epithelial Cells and Cytotoxicity Evaluation in Major Organ Cell Representatives. J. Nat. Prod. 2021, 84, 1261–1270. [Google Scholar] [CrossRef]
- Freeman, M.C.; Graham, R.L.; Lu, X.; Peek, C.T.; Denison, M.R. Coronavirus Replicase-Reporter Fusions Provide Quantitative Analysis of Replication and Replication Complex Formation. J. Virol. 2014, 88, 5319–5327. [Google Scholar] [CrossRef] [PubMed]
- Yount, B.; Denison, M.R.; Weiss, S.R.; Baric, R.S. Systematic Assembly of a Full-Length Infectious cDNA of Mouse Hepatitis Virus Strain A59. J. Virol. 2002, 76, 11065–11078. [Google Scholar] [CrossRef]
- Su, H.; Yao, S.; Zhao, W.; Li, M.; Liu, J.; Shang, W.; Xie, H.; Ke, C.; Gao, M.; Yu, K.; et al. Discovery of Baicalin and Baicalein as Novel, Natural Product Inhibitors of SARS-CoV-2 3CL Protease in Vitro. BioRxiv, 2020; Preprint. [Google Scholar] [CrossRef]
- Feng, J.; Li, D.; Zhang, J.; Yin, X.; Li, J. Crystal Structure of SARS-CoV 3C-like Protease with Baicalein. Biochem. Biophys. Res. Commun. 2022, 611, 190–194. [Google Scholar] [CrossRef]
- St John, S.E.; Tomar, S.; Stauffer, S.R.; Mesecar, A.D. Targeting Zoonotic Viruses: Structure-Based Inhibition of the 3C-like Protease from Bat Coronavirus HKU4–The Likely Reservoir Host to the Human Coronavirus That Causes Middle East Respiratory Syndrome (MERS). Bioorganic Med. Chem. 2015, 23, 6036–6048. [Google Scholar] [CrossRef]
- Huynh, J.; Li, S.; Yount, B.; Smith, A.; Sturges, L.; Olsen, J.C.; Nagel, J.; Johnson, J.B.; Agnihothram, S.; Gates, J.E.; et al. Evidence Supporting a Zoonotic Origin of Human Coronavirus Strain NL63. J. Virol. 2012, 86, 12816–12825. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Andrographolide 1 | Baicalin | Baicalein | |
|---|---|---|---|
| Cytotoxicity (CC50) | 70.4 μM | 379.1 μM | 224.4 μM |
| H5-MHV-FFL Inhibition | |||
| Co-treatment EC50 | - | 48.0 μM | 8.0 μM |
| Pre-treatment EC50 | - | 183.3 μM | 11.5 μM |
| Post-treatment EC50 | - | 203.3 μM | 111.2 μM |
| O5-MHV-FFL Inhibition | |||
| Co-treatment EC50 | - | 3.4 μM | 68.9 μM |
| Pre-treatment EC50 | - | >300 μM | >300 μM |
| Post-treatment EC50 | - | 224.0 μM | >300 μM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huffman, E.R.; Franges, J.X.; Doster, J.M.; Armstrong, A.R.; Batista, Y.S.; Harrison, C.M.; Brooks, J.D.; Thomas, M.N.; Butler Student Virology Group; Tomar, S.; et al. Development of Mouse Hepatitis Virus Chimeric Reporter Viruses Expressing the 3CLpro Proteases of Human Coronaviruses HKU1 and OC43 Reveals Susceptibility to Inactivation by Natural Inhibitors Baicalin and Baicalein. COVID 2024, 4, 208-220. https://doi.org/10.3390/covid4020016
Huffman ER, Franges JX, Doster JM, Armstrong AR, Batista YS, Harrison CM, Brooks JD, Thomas MN, Butler Student Virology Group, Tomar S, et al. Development of Mouse Hepatitis Virus Chimeric Reporter Viruses Expressing the 3CLpro Proteases of Human Coronaviruses HKU1 and OC43 Reveals Susceptibility to Inactivation by Natural Inhibitors Baicalin and Baicalein. COVID. 2024; 4(2):208-220. https://doi.org/10.3390/covid4020016
Chicago/Turabian StyleHuffman, Elise R., Jared X. Franges, Jayden M. Doster, Alexis R. Armstrong, Yara S. Batista, Cameron M. Harrison, Jon D. Brooks, Morgan N. Thomas, Butler Student Virology Group, Sakshi Tomar, and et al. 2024. "Development of Mouse Hepatitis Virus Chimeric Reporter Viruses Expressing the 3CLpro Proteases of Human Coronaviruses HKU1 and OC43 Reveals Susceptibility to Inactivation by Natural Inhibitors Baicalin and Baicalein" COVID 4, no. 2: 208-220. https://doi.org/10.3390/covid4020016