
Spatial Distribution of Citrus Pseudocercospora Leaf and Fruit Spot Disease and Shade Effect on Disease Intensity



Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.1.1. Biophysical Characterization of Sites
- ○
- The first one was located in the forest–savanna transition area of Bokito (latitude 4°38′ N, longitude 11°09′ E, altitude 480 m), situated in the western part of the humid forest agroecological zone of Cameroon. The annual mean temperature is 25 °C and relative humidity averaged 75%. The rainfall pattern is bimodal, i.e., there are two rainy seasons, spanning March-June and September-November (average annual rainfall: 1300 mm to 2500 mm). The soils are sandy loam, sandy clay loam, or clay in texture. This site was chosen because it is one of the main citrus production basins of the country, with medium PLFSD incidence.
- ○
- The second site was located in Foumbot in the western highland zone of Cameroon, (latitude 5°30′ N, longitude 10°37′ E, altitude 1010 m). This site was selected because of its situation in an area of high PLFSD incidence [6]. The annual mean temperature is 19 °C and the average relative humidity is >75%, with annual rainfall ranging from 1500 to 2500 mm which occurs in a unimodal pattern (one rainy season from March to November). Soils are predominantly volcanic, loam-like, clay-like in texture.
2.1.2. Bokito Plot
2.1.3. Foumbot Plot
2.2. Disease Incidence and Shade Data Collection
2.2.1. Bokito Plot
2.2.2. Foumbot Plot
2.3. Statistical Analysis
2.3.1. Analysis of Variance
2.3.2. Mapping of PLFSD Incidence and Shade
2.3.3. Residue Analysis
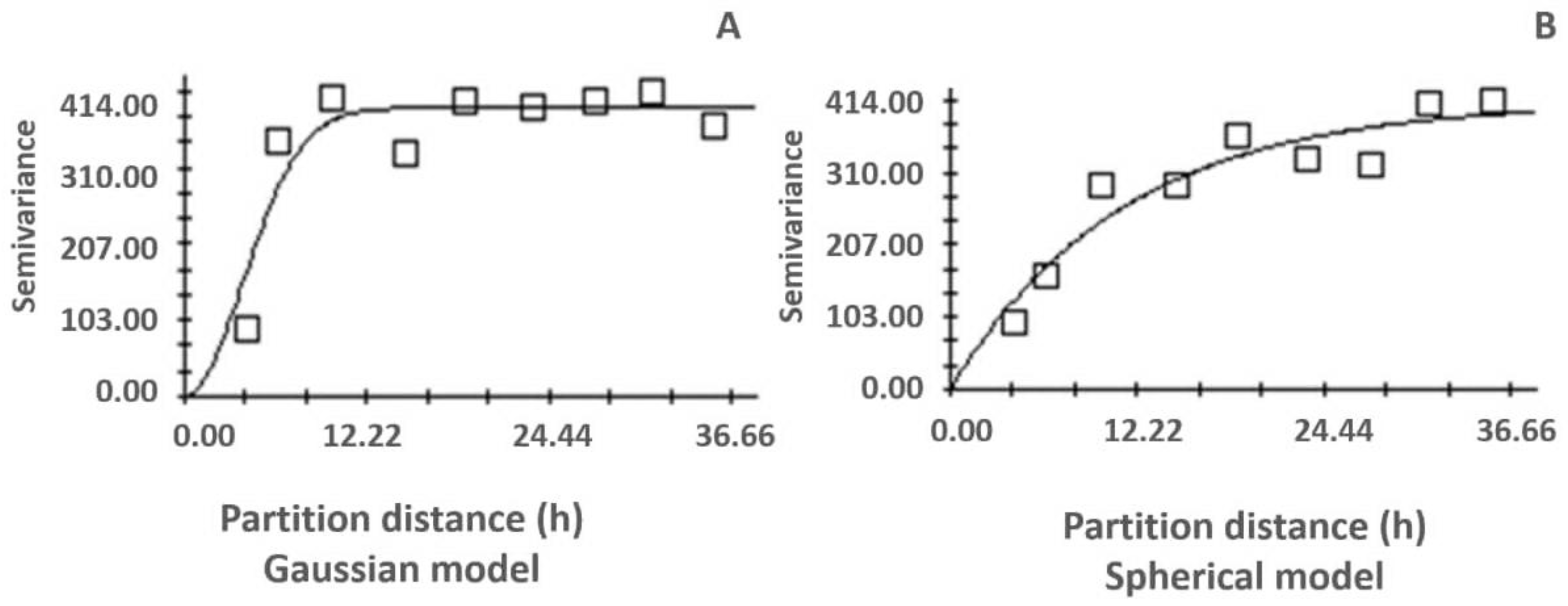
2.3.4. Spatial Analysis
- -
- C0: The nugget effect is the y value at which the curve of the model cuts the y axis;
- -
- a: The semi-variogram may reach a plateau. Reaching a plateau indicates that, from a certain distance, there is no longer spatial dependence between the data. This distance is called the range (a);
- -
- C0+C: The bearing is the variance at which the plateau appears. The bearing is reached by an asymptote (Figure 2).
3. Results
3.1. Variation of Disease Incidence with Shade Intensity
3.1.1. Bokito Site
3.1.2. Foumbot Site
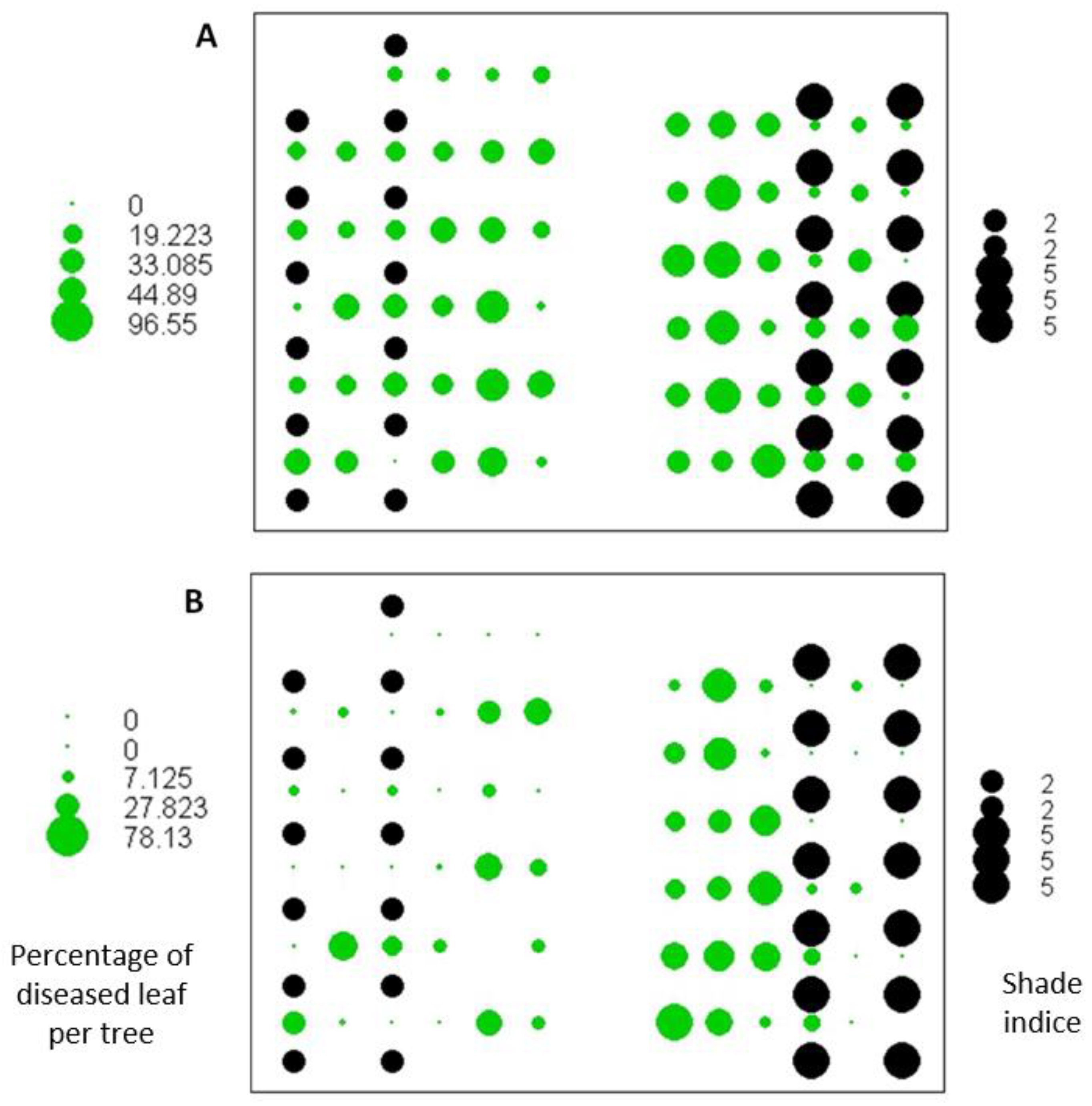
3.2. Representation of Disease Incidence According to Shade Intensity
3.3. Shade Effect and Observation Date on PLFSD Intensity
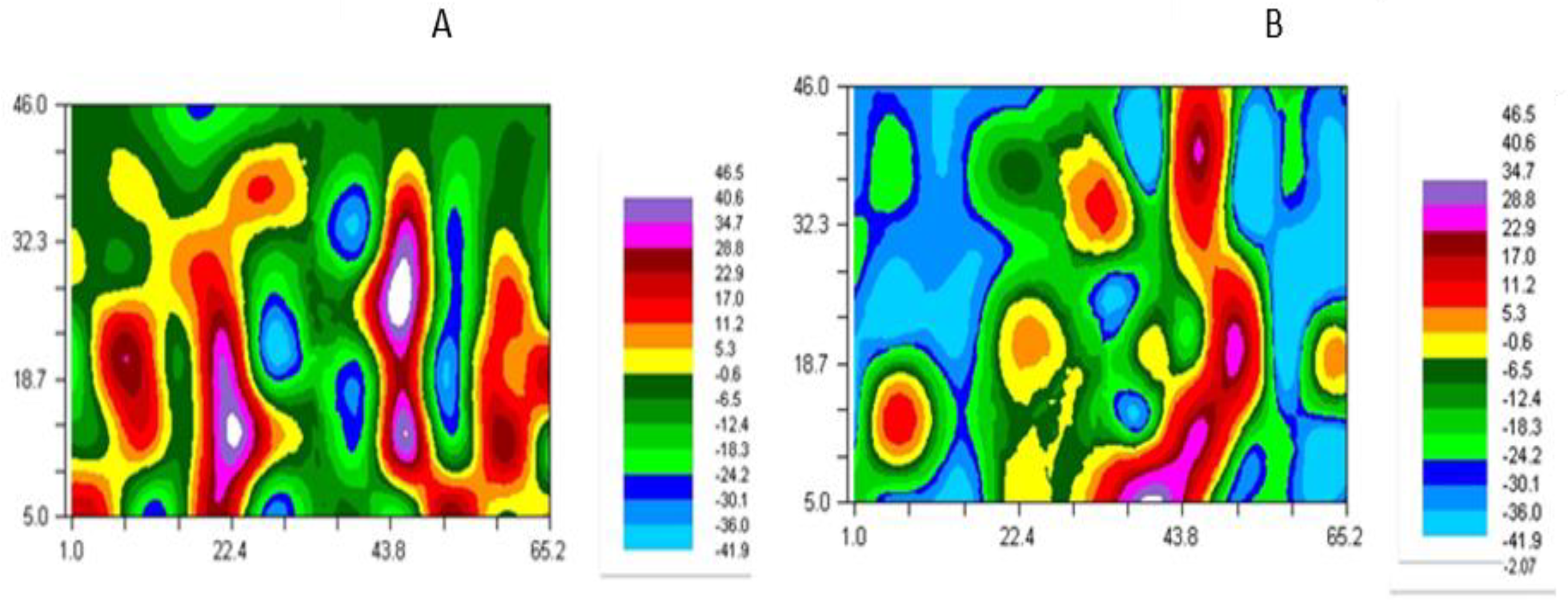
3.4. Spatial Analysis of Residues
4. Discussion
4.1. Shade Trees Effect
4.2. Spatial Distribution of PLFSD
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Driciru, P.; Mugasa, M.C.; Acidri, R.; Adriko, J. Development of Loop-Mediated Isothermal Amplification (LAMP) Assay for Detection of Pseudocercospora Angolensis in Sweet Orange. bioRxiv 2021. [Google Scholar] [CrossRef]
- Yesuf, M. Pseudocercospora Leaf and Fruit Spot Disease of Citrus: Achievements and Challenges in the Citrus Industry: A Review. Agric. Sci. 2013, 04, 324–328. [Google Scholar] [CrossRef] [Green Version]
- Brentu, F.C.; Cornelius, E.W.; Lawson, L.E.V.; Oduro, K.A.; Vicent, A. First Report of Pseudocercospora Angolensis Causing Fruit and Leaf Spot of Citrus in Ghana. Plant Dis. 2013, 97, 1661. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, T.; Mendes, O. Una Cercosporiose Em Citrinos. Notιcia Preliminar. Mocßambique Doc. Trimest. 1952, 72, 5–8. [Google Scholar]
- Ndo, E.G.D.; Kuate, J.; Sidjeu Wonfa, C.S.; Tchio, F.; Ndzana Abanda, F.X.; Mbieji Kemayou, C.; Akoutou Mvondo, E.; Amele Ndjoumoui, C.; Amang, A.; Mbang, J. Tolerance of Citrus Genotypes towards Pseudocercospora Leaf and Fruit Spot Disease in Western Highlands Zone of Cameroon. Crop Prot. 2019, 124, 104828. [Google Scholar] [CrossRef]
- Ndo, E.; Manga, B.; Ndoumbe-Nkeng, M.; Cilas, C. Distribution of Pseudocercospora Fruit and Leaf Spot, Phytophthora Foot Rot and Scab Diseases and Their Effect on Citrus Tree Decline Prevalence in the Humid Zones of Cameroon. Fruits 2019, 74, 249–256. [Google Scholar] [CrossRef]
- Ndo, E.; Bella-Manga, F.; Ndindeng, S.A.; Ndoumbe-Nkeng, M.; Fontem, D.; Cilas, C. Altitude, Tree Species and Soil Type Are the Main Factors Influencing the Severity of Phaeoramularia Leaf and Fruit Spot Disease of Citrus in the Humid Zones of Cameroon. Eur. J. Plant Pathol. 2010, 128, 385–397. [Google Scholar] [CrossRef]
- Dagnew, A.; Belew, D.; Admassu, B.; Yesuf, M. Citrus Production, Constraints and Management Practices in Ethiopia: The Case of Pseudocercospora Leaf and Fruit Spot Disease. Sci. Technol. Arts Res. J. 2014, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Kupagme, J.Y. Epidemiology and Control of Pseudocercospora Fruit and Leaf Spot Disease of Sweet Orange (Citrus sinensis (L.) Osbeck). Master’s Thesis, University of Ghana, Accra, Ghana, 2019. [Google Scholar]
- Pretorius, M.C. Epidemiology and Control of Pseudocercospora Angolensis Fruit and Leaf Spot Disease on Citrus in Zimbabwe. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2005. [Google Scholar]
- Kuate, J.; Fouré, E.; Tchio, F.; Ducelier, D. La Phaeoramulariose Des Agrumes Au Cameroun Due à Phaeoramularia Angolensis: Expression Parasitaire à Différentes Altitudes. Fruits 2002, 57, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Manga, M.; Dubois, C.; Kuaté, J.; Ngbwa, M.M.; Rey, J.-Y.; Développement, I. de la recherche agricole pour le; CIRAD-FLHOR Sensibilité à Phaeoramularia angolensis de divers agrumes cultivés en zone forestière humide au Cameroun. Fruits 1999, 54, 167–176. [Google Scholar]
- Sonwa, D.J.; Nkongmeneck, B.A.; Weise, S.F.; Tchatat, M.; Adesina, A.A.; Janssens, M.J. Diversity of Plants in Cocoa Agroforests in the Humid Forest Zone of Southern Cameroon. Biodivers. Conserv. 2007, 16, 2385–2400. [Google Scholar] [CrossRef]
- Mvondo, E.A.; Danièle Ndo, E.G.; Nomo, L.B.; Ambang, Z.; Manga, F.B.; Cilas, C. Tree Diversity and Shade Rate in Complex Cocoa-Based Agroforests Affect Citrus Foot Rot Disease. Basic Appl. Ecol. 2022, 64, 134–146. [Google Scholar] [CrossRef]
- Akoutou Mvondo, E.; Ndo, E.G.D.; Tsouga Manga, M.L.; Aba’ane, C.L.; Abondo Bitoumou, J.; Manga, B.; Bidzanga Nomo, L.; Ambang, Z.; Cilas, C. Effects of Complex Cocoa-Based Agroforests on Citrus Tree Decline. Crop Prot. 2020, 130, 105051. [Google Scholar] [CrossRef]
- Armengot, L.; Ferrari, L.; Milz, J.; Velásquez, F.; Hohmann, P.; Schneider, M. Cacao Agroforestry Systems Do Not Increase Pest and Disease Incidence Compared with Monocultures under Good Cultural Management Practices. Crop Prot. 2020, 130, 105047. [Google Scholar] [CrossRef]
- Barrios, E.; Valencia, V.; Jonsson, M.; Brauman, A.; Hairiah, K.; Mortimer, P.E.; Okubo, S. Contribution of Trees to the Conservation of Biodiversity and Ecosystem Services in Agricultural Landscapes. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2018, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Asigbaase, M.; Sjogersten, S.; Lomax, B.H.; Dawoe, E. Tree Diversity and Its Ecological Importance Value in Organic and Conventional Cocoa Agroforests in Ghana. PLoS ONE 2019, 14, e0210557. [Google Scholar] [CrossRef] [Green Version]
- Ratnadass, A.; Fernandes, P.; Avelino, J.; Habib, R. Plant Species Diversity for Sustainable Management of Crop Pests and Diseases in Agroecosystems: A Review. Agron. Sustain. Dev. 2012, 32, 273–303. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, M.; Raphael, I.A.; Ekbom, B.; Kyamanywa, S.; Karungi, J. Contrasting Effects of Shade Level and Altitude on Two Important Coffee Pests. J. Pest Sci. 2015, 88, 281–287. [Google Scholar] [CrossRef]
- Seif, A.A.; Hillocks, R.J. Phaeoramularia Fruit and Leaf Spot of Citrus with Special Reference to Kenya. Int. J. Pest Manag. 1993, 39, 44–50. [Google Scholar] [CrossRef]
- Seif, A.A.; Hillocks, R.J. Some Factors Affecting Infection of Citrus by Phaeoramularia angolensis. J. Phytopathol. (1986) 1998, 146, 385–391. [Google Scholar]
- Schroth, G.; Krauss, U.; Gasparotto, L.; Duarte, J.A.; Vohland, K. Pests and Diseases in Agroforestry Systems of the Humid Tropics. Agrofor. Syst. 2000, 50, 199–241. [Google Scholar] [CrossRef]
- Kuaté, J.; Kouodiekong, L.; David, O.; Ndindeng, S.A.; Parrot, L. Les Exploitations Fruitières en Zones Périurbaines de Yaoundé au Cameroun: Une Enquête Diagnostic. Available online: https://agritrop.cirad.fr/529960/ (accessed on 20 October 2020).
- Aka, A.R.; Kouassi, N.K.; Agnéroh, T.A.; Amancho, N.A.; Sangare, A. Distribution et Incidence de La Mosaïque Du Concombre (Cmv) Dans Des Bananeraies Industrielles Au Sud-Est de La Côte d’ivoire. Sci. Nat. 2009, 6. [Google Scholar] [CrossRef]
- Baillargeon, S. Le Krigeage: Revue de La Théorie et Application à l’interpolation Spatiale de Données de Précipitations. Ph.D. Thesis, Université Laval, Québec, QC, Canada, 2005. [Google Scholar]
- Gratton, Y. Le Krigeage: La Méthode Optimale d’interpolation Spatiale. Les Artic. L’Institut D’Analyse Géographique 2022, 1. Available online: https://www.researchgate.net/profile/Yves-Gratton/publication/229009107_Le_krigeage_la_methode_optimale_d%27interpolation_spatiale/links/00b49533adff3e8501000000/Le-krigeage-la-methode-optimale-dinterpolation-spatiale.pdf (accessed on 20 June 2022).
- Mouen Bedimo, J.A.; Bieysse, D.; Njiayouom, I.; Deumeni, J.P.; Cilas, C.; Nottéghem, J.L. Effect of Cultural Practices on the Development of Arabica Coffee Berry Disease, Caused by Colletotrichum Kahawae. Eur. J. Plant Pathol. 2007, 119, 391–400. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Bhagwat, S.A.; Buchori, D.; Faust, H.; Hertel, D.; Hölscher, D.; Juhrbandt, J.; Kessler, M.; Perfecto, I.; et al. Multifunctional Shade-Tree Management in Tropical Agroforestry Landscapes—A Review. J. Appl. Ecol. 2011, 48, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, M.S. Crop Strength through Diversity. Nature 2000, 406, 681–682. [Google Scholar] [CrossRef] [PubMed]
- Ndo, E.G.D.; Mvondo, E.A.; Ambang, Z.; Manga, B.; Cilas, C.; Nomo, L.B.; Gidoin, C.; Bieng, M.-A.N. Spatial Organisation Influences Citrus Pseudocercospora Leaf and Fruit Spot Disease Severity in Cocoa-Based Agroforestry Systems. Available online: /paper/Spatial-organisation-influences-citrus-leaf-and-in-Ndo-Mvondo/5f9aab68d387c331b3af966ab0b93810b7b9e672 (accessed on 21 October 2020).
- Akoutou, E.; Ndo, E.; Bieng, M.-A.; Ambang, Z.; Manga, B.; Cilas, C.; Manga, M.; Bidzanga Nomo, L. Assessment of the Interaction between the Spatial Organization of Citrus Trees Populations in Cocoa Agroforests and Phytophthora Foot Rot Disease of Citrus Severity. Agrofor. Syst. 2017, 93, 493–502. [Google Scholar] [CrossRef]
- Ratnadass, A.; Avelino, J.; Fernandes, P.; Letourmy, P.; Babin, R.; Deberdt, P.; Deguine, J.-P.; Grechi, I.; Naudin, K.; Rhino, B.; et al. Synergies and Tradeoffs in Natural Regulation of Crop Pests and Diseases under Plant Species Diversification. Crop Prot. 2021, 146, 105658. [Google Scholar] [CrossRef]
- Pumariño, L.; Sileshi, G.W.; Gripenberg, S.; Kaartinen, R.; Barrios, E.; Muchane, M.N.; Midega, C.; Jonsson, M. Effects of Agroforestry on Pest, Disease and Weed Control: A Meta-Analysis. Basic Appl. Ecol. 2015, 16, 573–582. [Google Scholar] [CrossRef]
- Hatvani, I.G.; de Barros, V.D.; Tanos, P.; Kovács, J.; Székely Kovács, I.; Clement, A. Spatiotemporal Changes and Drivers of Trophic Status over Three Decades in the Largest Shallow Lake in Central Europe, Lake Balaton. Ecol. Eng. 2020, 151, 105861. [Google Scholar] [CrossRef]
- Ndo, E.G.D. Evaluation des Facteurs de Risque Épidémiologique de la Phaeoramulariose des Agrumes Dans les Zones Humides du Cameroun. Ph.D. Thesis, University of Montpellier, Montpellier, France, 2011. [Google Scholar]
- Merle, I.; Villarreyna-Acuña, R.; Ribeyre, F.; Roupsard, O.; Cilas, C.; Avelino, J. Microclimate Estimation under Different Coffee-Based Agroforestry Systems Using Full-Sun Weather Data and Shade Tree Characteristics. Eur. J. Agron. 2022, 132, 126396. [Google Scholar] [CrossRef]
- Mustafa, M.; Szalai, Z.; Divéky-Ertsey, A.; Gál, I.; Csambalik, L. Conceptualizing Multiple Stressors and Their Consequences in Agroforestry Systems. Stresses 2022, 2, 242–255. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observation Date | Shade Type | Incidence (% ± x ) |
|---|---|---|
| September 2020 | Full sunlight | 3.12 ± 6.19 a |
| Forest trees shade | 0.00 ± 0.00 a | |
| April 2021 | Full sunlight | 6.70 ± 10.57 a |
| Forest trees shade | 0.0 ± 0.00 b | |
| September 2021 | Full sunlight | 22.44 ± 21.77 a |
| Forest trees shade | 0.00 ± 0.00 b | |
| April 2022 | Full sunlight | 2.40 ± 4.65 a |
| Forest trees shade | 0.00 ± 0.00 a |
| Observation Date | Shade Type | Incidence (% ± x ) |
|---|---|---|
| April 2021 | Full sunlight | 1.34 ± 3.54 a |
| Forest trees shade | 0.0 ± 0.00 a | |
| September 2021 | Full sunlight | 23.47 ± 26.18 a |
| Forest trees shade | 0.83 ± 2.04 b | |
| April 2022 | Full sunlight | 6.59 ± 17.44 b |
| Forest trees shade | 0.00 ± 0.00 a |
| Observation Date | Shade Type | Incidence (% ± x) |
|---|---|---|
| October 2021 | Full sunlight | 45.26 ± 23.92 a |
| Avocado trees shade | 21.39 ± 13.70 b | |
| Mango trees shade | 27.76 ± 14.33 b | |
| June 2022 | Full sunlight | 25.20 ± 23.05 a |
| Avocado trees shade | 3.02 ± 5.54 b | |
| Mango trees shade | 8.11± 13.80 b |
| Variables | Parameters | DDl | Sum of Square | Means Square | F Value | Pr (>F) |
|---|---|---|---|---|---|---|
| Incidence | Shade | 3 | 21,292 | 7097.3 | 23.1 | 4.5 × 10−12 *** |
| Observation date | 1 | 12,493 | 12,493.4 | 40.7 | 2.8 × 10−9 *** | |
| Interaction shade x observation date | - | - | - | - | - | |
| Residue | 131 | 40,238 | 307.2 | - | - |
| Variables | Dates | Model | Reach (m) | Degree (C0+C) | C/(C0+C) | RSS | R2 |
|---|---|---|---|---|---|---|---|
| Severity | October 2021 | Gaussian | 11.70 | 0.914 | 0.999 | 0.106 | 0.883 |
| June 2022 | Gaussian | 10.08 | 0.249 | 1.000 | 0.015 | 0.704 | |
| Incidence | October 2021 | Gaussian | 9.59 | 391.10 | 0.997 | 15660 | 0.829 |
| June 2022 | Exponential | 7.49 | 314.60 | 0.999 | 5593 | 0.425 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndo, E.G.D.; Akoutou Mvondo, E.; Bella Manga, F.; Bidzanga Nomo, L.; Cilas, C. Spatial Distribution of Citrus Pseudocercospora Leaf and Fruit Spot Disease and Shade Effect on Disease Intensity. Crops 2023, 3, 11-23. https://doi.org/10.3390/crops3010002
Ndo EGD, Akoutou Mvondo E, Bella Manga F, Bidzanga Nomo L, Cilas C. Spatial Distribution of Citrus Pseudocercospora Leaf and Fruit Spot Disease and Shade Effect on Disease Intensity. Crops. 2023; 3(1):11-23. https://doi.org/10.3390/crops3010002
Chicago/Turabian StyleNdo, E. G. D., E. Akoutou Mvondo, F. Bella Manga, L. Bidzanga Nomo, and C. Cilas. 2023. "Spatial Distribution of Citrus Pseudocercospora Leaf and Fruit Spot Disease and Shade Effect on Disease Intensity" Crops 3, no. 1: 11-23. https://doi.org/10.3390/crops3010002