
Habitat Mapping and Spatiotemporal Overlap of the Amazon River Dolphin, Fishers, and Tourism in the Central Region of the Brazilian Amazon

Abstract
:1. Introduction
1.1. Human–Boto Interactions and Conflict
1.2. Fisher, BIP, and Boto Spaces of Conflict
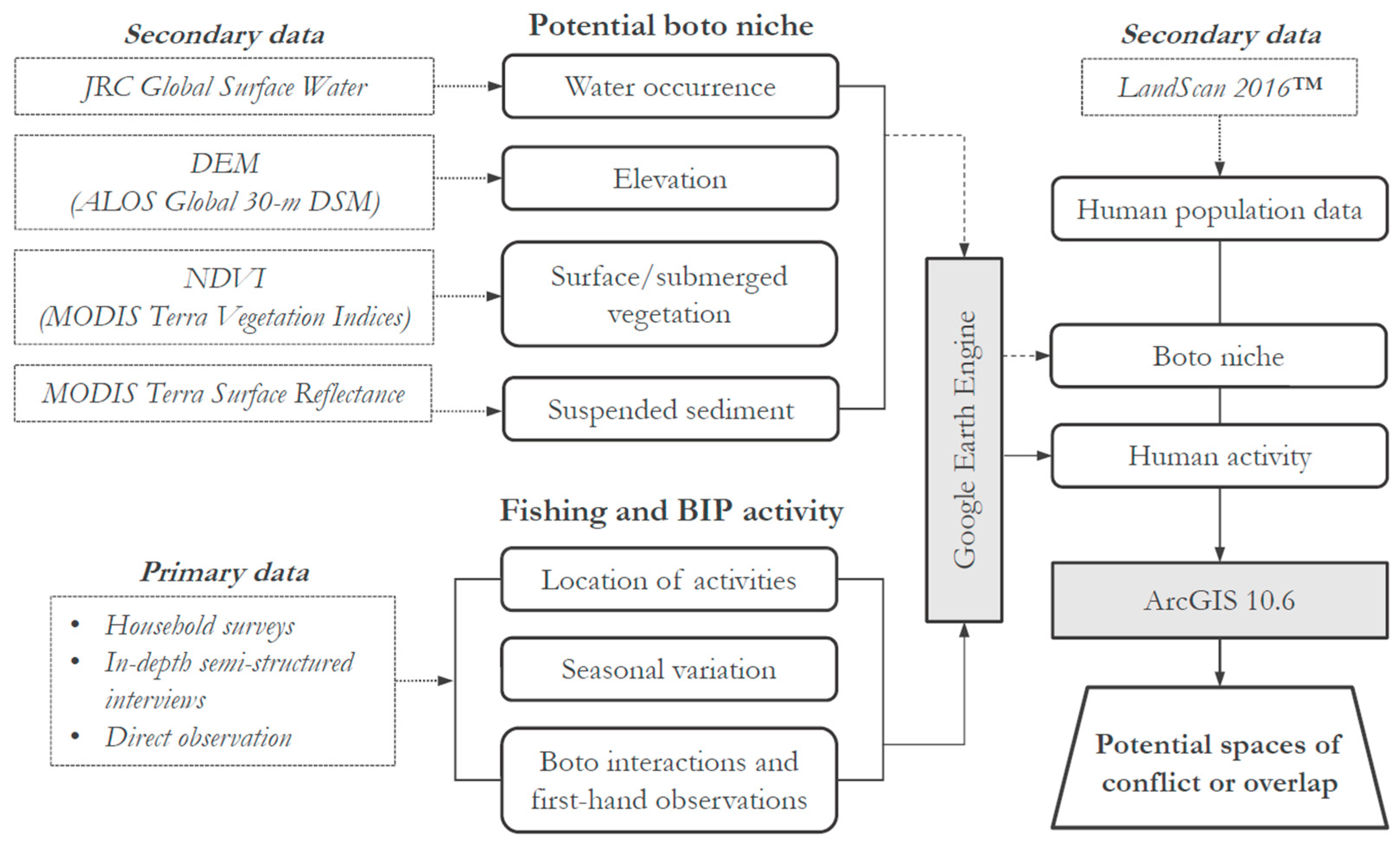
2. Materials and Methods
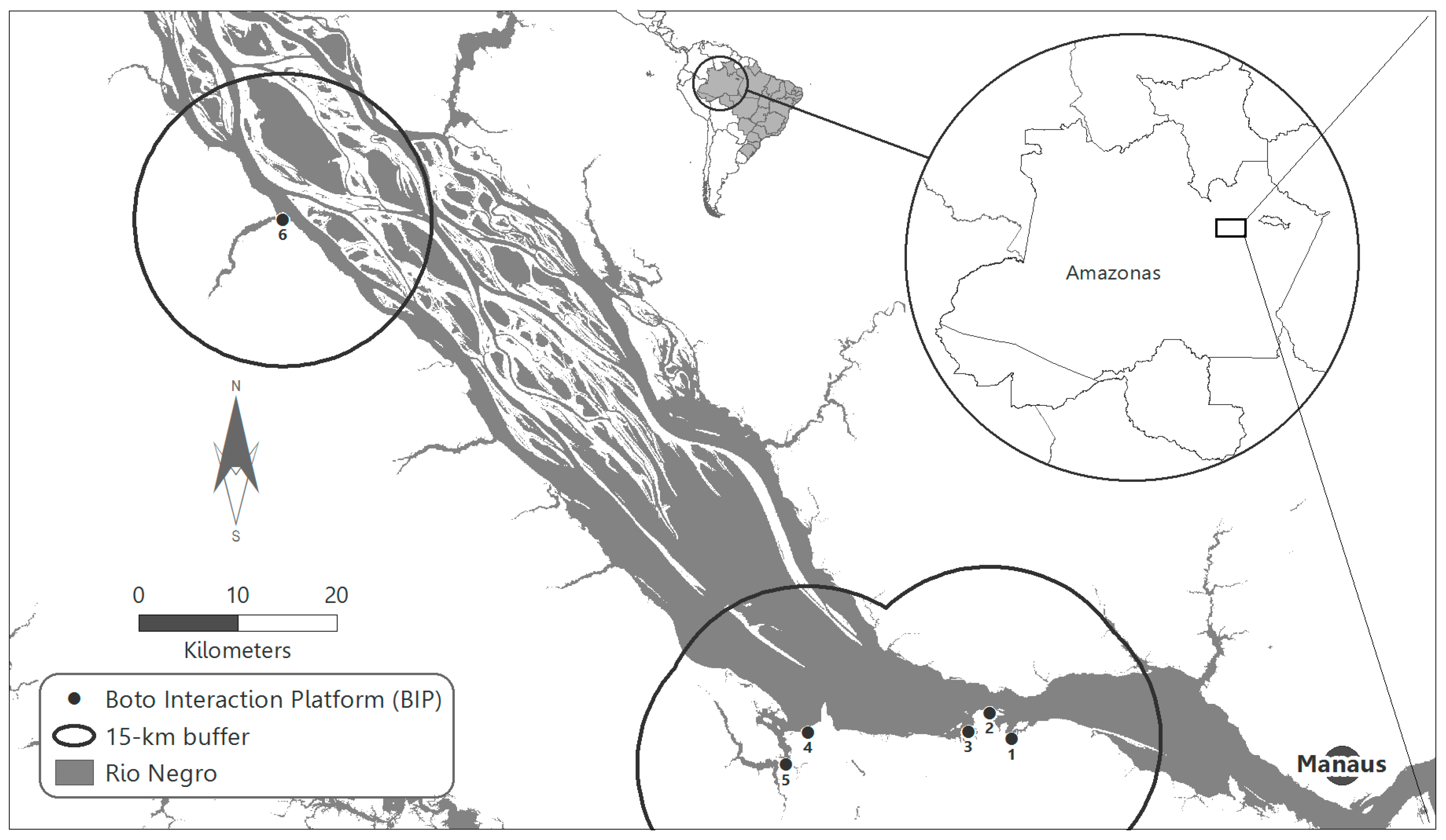
2.1. Field Data Collection and Study Area
2.2. Fisher and BIP Spatiotemporal Data
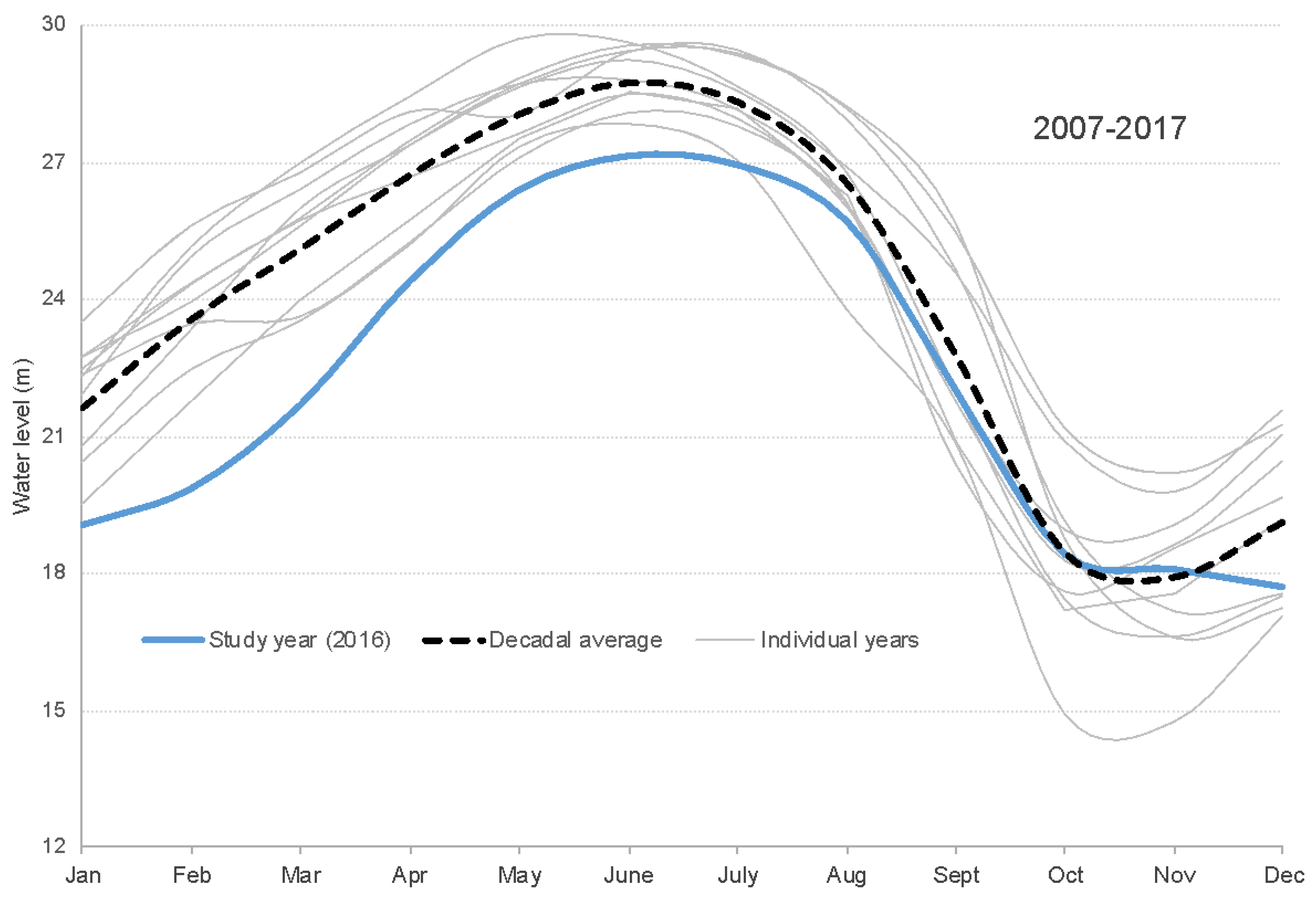
2.3. Seasonal Boto Habitat and Temporal Parameterization
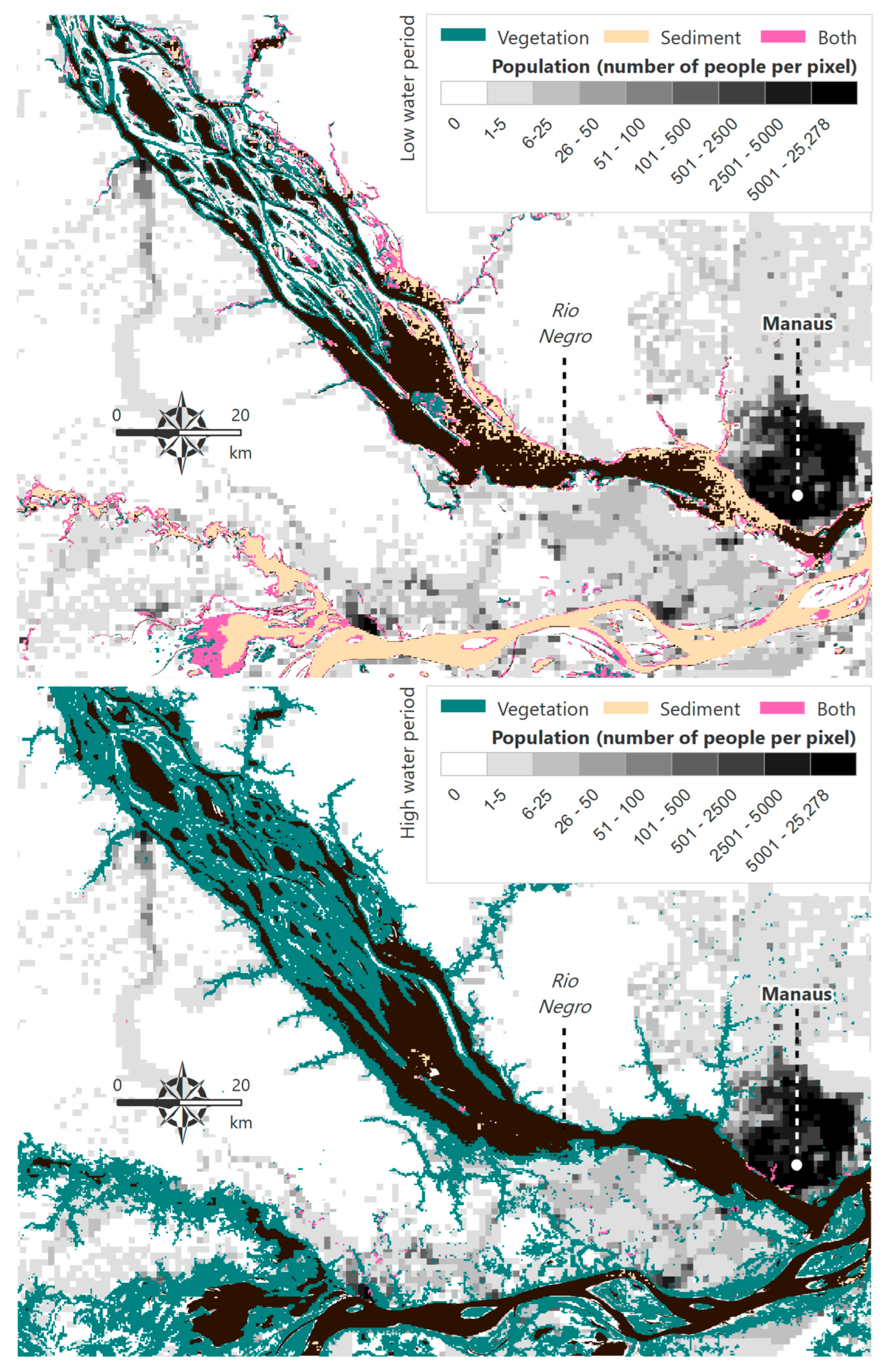
2.4. Surface and Submerged Vegetation
2.5. Suspended Sediment
2.6. Identifying Boto Niche
3. Results
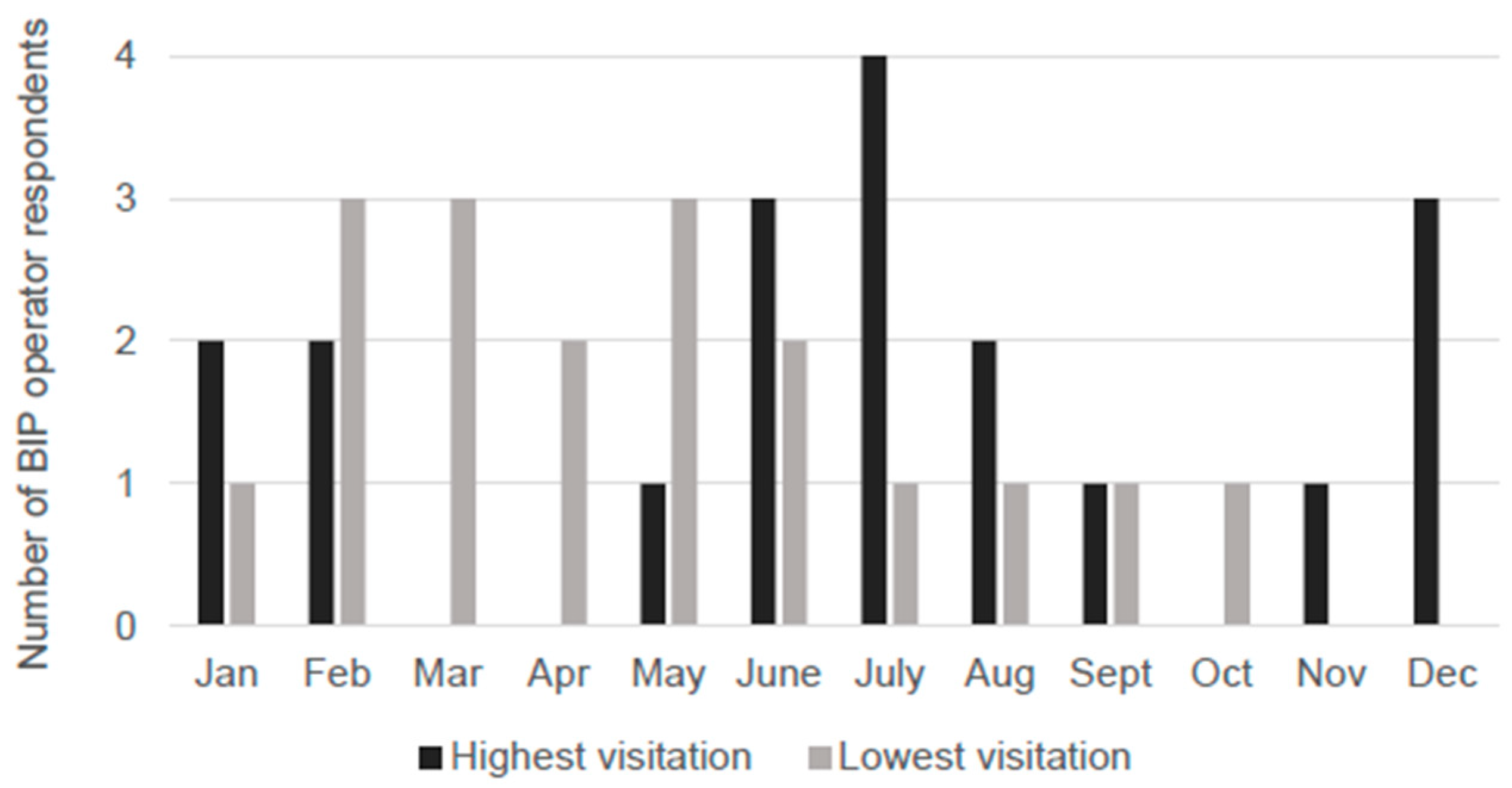
3.1. Spatial and Temporal Variation of BIP Activity
3.2. Spatial and Temporal Variation of Fishing Activity
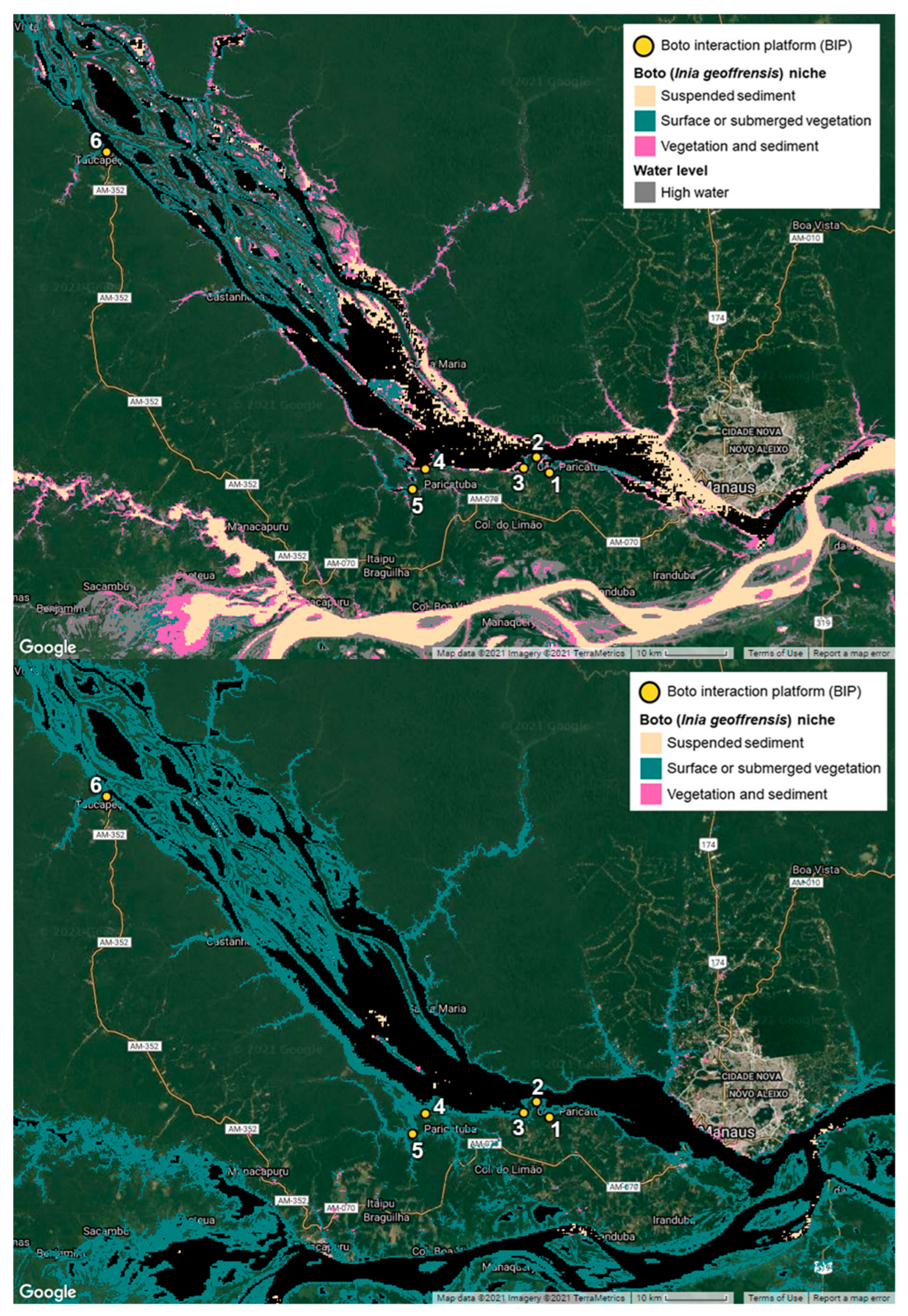
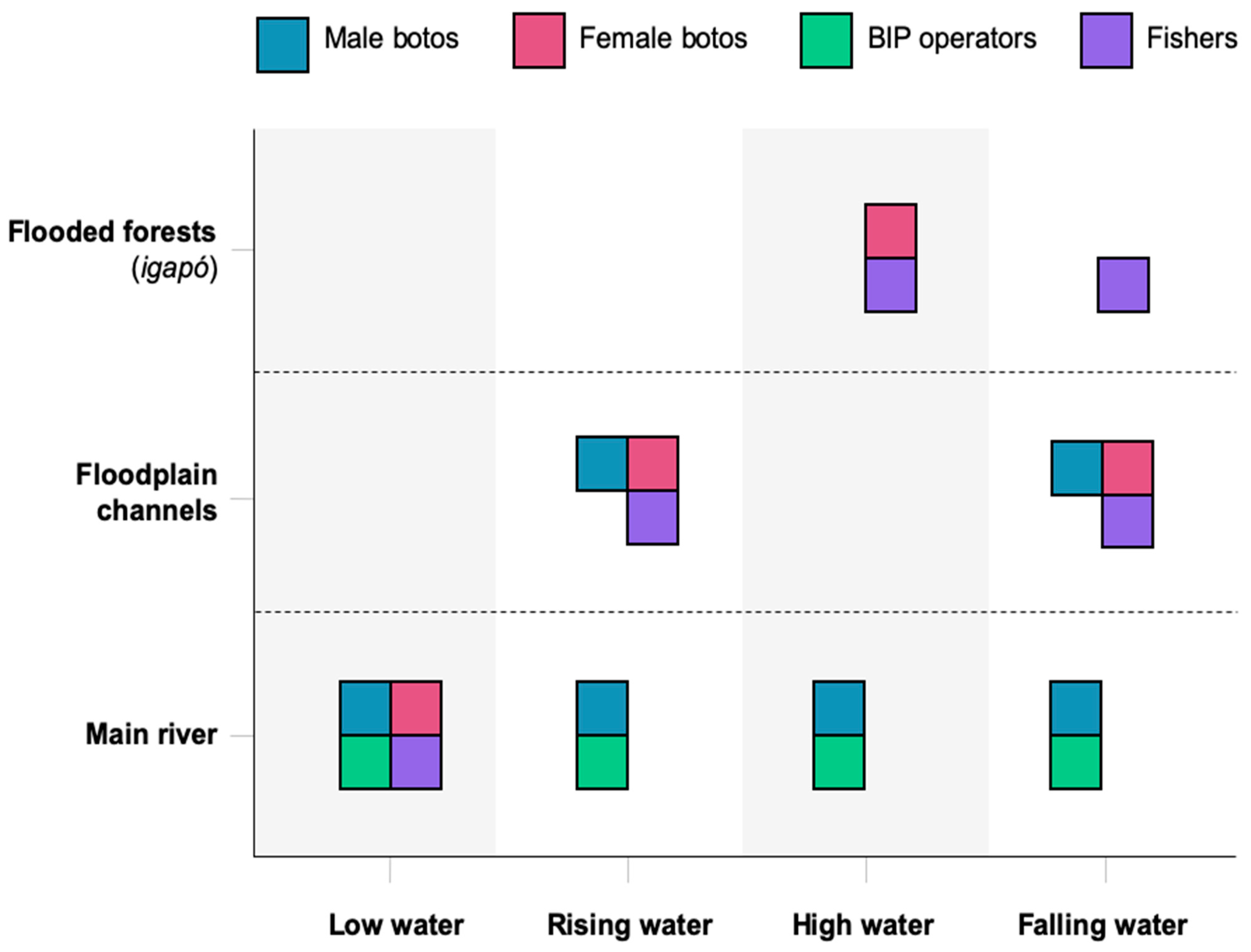
3.3. Boto Niche and Spaces of Contention
4. Discussion
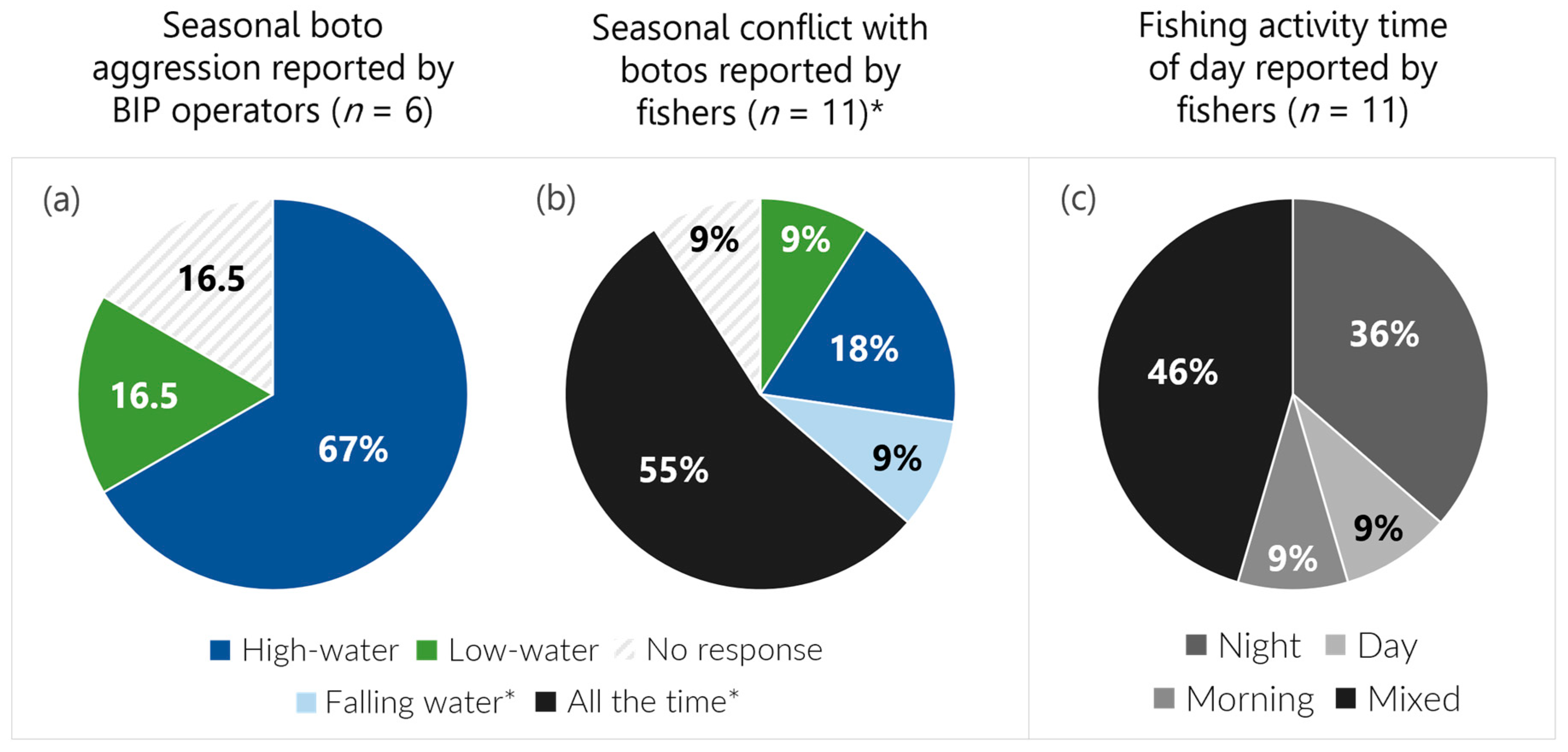
4.1. Seasonality of BIP Activities and Observed Boto Behavior
4.2. Spatial and Temporal Fishing Considerations
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- da Silva, V.M.F.; Freitas, C.E.C.; Dias, R.L.; Martin, A.R. Both cetaceans in the Brazilian Amazon show sustained, profound population declines over two decades. PLoS ONE 2018, 13, e0191304. [Google Scholar] [CrossRef] [PubMed]
- McGuire, T.L.; Henningsen, T. Movement Patterns and Site Fidelity of River Dolphins (Inia geoffrensis and Sotalia fluviatilis) in the Peruvian Amazon as Determined by Photo-Identification. Aquat. Mamm. 2007, 33, 359–367. [Google Scholar] [CrossRef]
- Araujo, C.C.; da Silva, V.M.F. Spatial distribution of river dolphins, Inia geoffrensis (Iniidae), in the Araguaia River (central Brazil). Mammalia 2014, 78, 481–486. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Akamatsu, T.; da Silva, V.M.F.; Kohshima, S. Local habitat use by botos (Amazon river dolphins, Inia geoffrensis) using passive acoustic methods. Mar. Mammal Sci. 2016, 32, 220–240. [Google Scholar] [CrossRef]
- Flores, P.A.C. Espécies-alvo do Pan Pequenos Cetáceos. In Plano de Ação Nacional para a Conservação dos Mamíferos Aquáticos-Pequenos Cetáceos; Rocha-Campos, C.C., Câmara, I.d.G., Pretto, D.J., Eds.; Instituto Chico Mendes de Conservação da Biodiversidade, Icmbio: Brasília, Brazil, 2010. [Google Scholar]
- Gomez-Salazar, C.; Trujillo, F.; Portocarrero-Aya, M.; Whitehead, H. Population, density estimates, and conservation of river dolphins (Inia and Sotalia) in the Amazon and Orinoco river basins. Mar. Mammal Sci. 2012, 28, 124–153. [Google Scholar] [CrossRef]
- Oliveira-da-Costa, M.; Marmontel, M.; Da-Rosa, D.S.X.; Coelho, A.; Wich, S.; Mosquera-Guerra, F.; Trujillo, F. Effectiveness of unmanned aerial vehicles to detect Amazon dolphins. Oryx 2019, 54, 696–698. [Google Scholar] [CrossRef]
- Watts, S.M.; McCarthy, T.M.; Namgail, T. Modelling potential habitat for snow leopards (Panthera uncia) in Ladakh, India. PLoS ONE 2019, 14, e0211509. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.S.F.; Georgiadis, G.; Campello, S.; Brandão, R.A.; Ciuti, S. Improving river dolphin monitoring using aerial surveys. Ecosphere 2017, 8, e01912. [Google Scholar] [CrossRef]
- Dawson, S.; Wade, P.; Slooten, E.; Barlow, J. Design and field methods for sighting surveys of cetaceans in coastal and riverine habitats. Mammal Rev. 2008, 38, 19–49. [Google Scholar] [CrossRef]
- Richman, N.I.; Gibbons, J.M.; Turvey, S.T.; Akamatsu, T.; Ahmed, B.; Mahabub, E.; Smith, B.D.; Jones, J.P.G. To see or not to see: Investigating detectability of Ganges river dolphins using a combined visual-acoustic survey. PLoS ONE 2014, 9, e96811. [Google Scholar] [CrossRef]
- Briscoe, D.K.; Fossette, S.; Scales, K.L.; Hazen, E.L.; Bograd, S.J.; Maxwell, S.M.; McHuron, E.A.; Robinson, P.W.; Kuhn, C.; Costa, D.P.; et al. Characterizing habitat suitability for a central-place forager in a dynamic marine environment. Ecol. Evol. 2018, 8, 2788–2801. [Google Scholar] [CrossRef] [PubMed]
- Derville, S.; Torres, L.G.; Iovan, C.; Garrigue, C. Finding the right fit: Comparative cetacean distribution models using multiple data sources and statistical approaches. Divers. Distrib. 2018, 24, 1657–1673. [Google Scholar] [CrossRef]
- Huang, S.L.; Peng, C.; Chen, M.; Wang, X.; Jefferson, T.A.; Xu, Y.; Yu, X.; Lao, Y.; Li, J.; Huang, H.; et al. Habitat configuration for an obligate shallow-water delphinid: The Indo-Pacific humpback dolphin, Sousa chinensis, in the Beibu Gulf (Gulf of Tonkin). Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 472–485. [Google Scholar] [CrossRef]
- Ruda, A.; Kolejka, J.; Silwal, T. GIS-assisted prediction and risk zonation of wildlife attacks in the Chitwan National Park in Nepal. ISPRS Int. J. Geo-Inf. 2018, 7, 369. [Google Scholar] [CrossRef]
- da Silva, V.M.F.; Best, R.C. Freshwater dolphin/fisheries interaction in the Central Amazon (Brazil). Amazoniana 1996, 14, 165–175. [Google Scholar]
- Loch, C.; Marmontel, M.; Simões-Lopes, P.C. Conflicts with fisheries and intentional killing of freshwater dolphins (Cetacea: Odontoceti) in the Western Brazilian Amazon. Biodivers. Conserv. 2009, 18, 3979–3988. [Google Scholar] [CrossRef]
- Alves, L.C.P.D.S.; Zappes, C.A.; Andriolo, A. Conflicts between river dolphins (Cetacea: Odontoceti) and fisheries in the Central Amazon: A path toward tragedy? Zoologia 2012, 29, 420–429. [Google Scholar] [CrossRef]
- Iriarte, V.; Marmontel, M. River dolphin (Inia geoffrensis, Sotalia fluviatilis) mortality events attributed to artisanal fisheries in the Western Brazilian Amazon. Aquat. Mamm. 2013, 39, 116–124. [Google Scholar] [CrossRef]
- Brum, S.M.; da Silva, V.M.F.; Rossoni, F.; Castello, L. Use of dolphins and caimans as bait for Calophysus macropterus (Lichtenstein, 1819) (Siluriforme: Pimelodidae) in the Amazon. J. Appl. Ichthyol. 2015, 31, 675–680. [Google Scholar] [CrossRef]
- da Silva, V.M.F.; Martin, A.R. Status, threats, conservation initiatives and possible solutions for Inia geoffrensis and Sotalia fluviatilis in Brazil. In The Action Plan for South American River Dolphins 2010–2020; Trujillo, F., Crespo, E., Van Damme, P.A., Usma, J.S., Eds.; WWF, Fundación Omacha, WDS, WDCS, and SOLAMAC: Bogota, Colombia, 2010; pp. 123–143. [Google Scholar]
- Mintzer, V.J.; Martin, A.R.; da Silva, V.M.F.; Barbour, A.B.; Lorenzen, K.; Frazer, T.K. Effect of illegal harvest on apparent survival of Amazon River dolphins (Inia geoffrensis). Biol. Conserv. 2013, 158, 280–286. [Google Scholar] [CrossRef]
- Alves, L.C.P.S.; Andriolo, A.; Orams, M.B.; Azevedo, A.F. The growth of “botos feeding tourism”, a new tourism industry based on the boto (Amazon river dolphin) Inia geoffrensis in the Amazonas State, Brazil. Sitientibus Ser. Ciências Biológicas 2011, 11, 8–15. [Google Scholar] [CrossRef]
- Baker, C.S.; Perry, A.; Vequist, G. Humpback whales of Glacier Bay, Alaska. Whalewatcher 1988, 21, 13–17. [Google Scholar]
- MacGibbon, J. Responses of Sperm Whales (Physeter macrocephalus) to Commercial Whale Watching Boats off the Coast of Kaikoura; Department of Conservation, University of Canterbury: Christchurch, New Zealand, 1991. [Google Scholar]
- Blane, J.M.; Jaakson, R. The impact of ecotourism boats on the Saint Lawrence beluga whales. Environ. Conserv. 1995, 21, 267–269. [Google Scholar] [CrossRef]
- Wursig, B. Swim-with-dolphin activities in nature: Weighing the pros and cons. Whalewatcher 1996, 30, 11–15. [Google Scholar]
- Constantine, R. Increased avoidance of swimmers by wild bottlenose dolphins (Tursiops truncatus) due to long-term exposure to swim-with-dolphin tourism. Mar. Mammal Sci. 2001, 17, 689–702. [Google Scholar] [CrossRef]
- Nowacek, S.M.; Wells, R.S. Short-term effects of boat traffic on bottlenose dolphins, Turciops truncatus, in Sarasota bay, Florida. Mar. Mammal Sci. 2001, 17, 673–688. [Google Scholar] [CrossRef]
- Williams, R.; Trites, A.W.; Bain, D.E. Behavioural responses of killer whales (Orcinus orca) to whale-watching boats: Opportunistic observations and experimental approaches. J. Zool. 2002, 256, 255–270. [Google Scholar] [CrossRef]
- Orams, M.B. Feeding wildlife as a tourism attraction: A review of issues and impacts. Tour. Manag. 2002, 23, 281–293. [Google Scholar] [CrossRef]
- Lusseau, D.; Higham, J.E.S. Managing the impacts of dolphin-based tourism through the definition of critical habitats: The case of bottlenose dolphins (Tursiops spp.) in Doubtful Sound, New Zealand. Tour. Manag. 2004, 25, 657–667. [Google Scholar] [CrossRef]
- Peters, K.J.; Parra, G.J.; Skuza, P.P.; Möller, L.M. First insights into the effects of swim-with-dolphin tourism on the behavior, response, and group structure of southern Australian bottlenose dolphins. Mar. Mammal Sci. 2013, 29, 484–497. [Google Scholar] [CrossRef]
- Christiansen, F.; Mchugh, K.A.; Bejder, L.; Siegal, E.M.; Lusseau, D.; Mccabe, E.B.; Lovewell, G.; Wells, R.S. Food provisioning increases the risk of injury in a long-lived marine top predator. R. Soc. Open Sci. 2016, 3, 160560. [Google Scholar] [CrossRef] [PubMed]
- Hazelkorn, R.A.; Schulte, B.A.; Cox, T.M. Persistent Effects of Begging on Common Bottlenose Dolphin (Tursiops truncatus) Behavior in an Estuarine Population. Aquat. Mamm. 2016, 42, 531–541. [Google Scholar] [CrossRef]
- Alves, L.C.P.S.; Andriolo, A.; Orams, M.B.; Azevedo, A.F. Resource defence and dominance hierarchy in the boto (Inia geoffrensis) during a provisioning program. Acta Ethologica 2013, 16, 9–19. [Google Scholar] [CrossRef]
- Hoefle, S.W. Multi-functionality, juxtaposition and conflict in the Central Amazon: Will tourism contribute to rural livelihoods and save the rainforest? J. Rural. Stud. 2016, 44, 24–36. [Google Scholar] [CrossRef]
- Pinaya, W.H.D.; Lobon-Cervia, F.J.; Pita, P.; De Souza, R.B.; Freire, J.; Isaac, V.J. Multispecies fisheries in the Lower Amazon River and its relationship with the regional and global climate variability. PLoS ONE 2016, 11, e0157050. [Google Scholar] [CrossRef] [PubMed]
- Vidal, O.; Barlow, J.; Hurtado, L.A.; Torre, J.; Cendon, P.; Ojeda, Z. Distribution and abundance of the Amazon River dolphin (Inia geoffrensis) and the tucuxi (Sotalia fluviatilis) in the upper Amazon River. Mar. Mammal Sci. 1997, 13, 427–445. [Google Scholar] [CrossRef]
- Martin, A.R.; da Silva, V.M.F. Number, seasonal movements, and residency characteristics of river dolphins in an Amazonian floodplain lake system. Can. J. Zool. 2004, 82, 1307–1315. [Google Scholar] [CrossRef]
- Martin, A.R.; da Silva, V.M.F. River dolphins and flooded forest: Seasonal habitat use and sexual segregation of botos (Inia geoffrensis) in an extreme cetacean environment. J. Zool. 2004, 263, 295–305. [Google Scholar] [CrossRef]
- Martin, A.R.; da Silva, V.M.F.; Salmon, D.L. Riverine habitat preferences of botos (Inia geoffrensis) and tucuxis (Sotalia fluviatilis) in the central Amazon. Mar. Mammal Sci. 2004, 20, 189–200. [Google Scholar] [CrossRef]
- Mintzer, V.J.; Lorenzen, K.; Frazer, T.K.; da Silva, V.M.F.; Martin, A.R. Seasonal movements of river dolphins (Inia geoffrensis) in a protected Amazonian floodplain. Mar. Mammal Sci. 2016, 32, 664–681. [Google Scholar] [CrossRef]
- Bright, E.A.; Rose, A.N.; Urban, M.L.; McKee, J.J. LandScan 2016; Oak Ridge National Laboratory SE: Oak Ridge, TN, USA, 2017. Available online: https://landscan.ornl.gov/ (accessed on 10 June 2018).
- da Silva, V.M.F. Ecologia Alimentar Dos Golfinhos Da Amazônia. Master’s Thesis, University of Amazonas, Manaus, Brazil, 1983. [Google Scholar]
- Crampton, W.G.R. Os peixes da Reserva Mamirauá: Diversidade e história natural na planície alagável da Amazônia. In Estratégias para Manejo de Recursos Pesqueiros em Mamirauá; Queiroz, H.L., Crampton, W.G.R., Eds.; Sociedade Civil Mamirauá/CNPq: Brasilia, Brazil, 1999; pp. 10–36. [Google Scholar]
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Fusilli, L.; Collins, M.O.; Laneve, G.; Palombo, A.; Pignatti, S.; Santini, F. Assessment of the abnormal growth of floating macrophytes in Winam Gulf (Kenya) by using MODIS imagery time series. Int. J. Appl. Earth Obs. Geoinf. 2012, 20, 33–41. [Google Scholar] [CrossRef]
- Marchetti, Z.Y.; Minotti, P.G.; Ramonell, C.G.; Schivo, F.; Kandus, P. NDVI patterns as indicator of morphodynamic activity in the middle Paraná River floodplain. Geomorphology 2016, 253, 146–158. [Google Scholar] [CrossRef]
- Didan, K. MOD13Q1 MODIS/Terra Vegetation Indices 16-Day L3 Global 250m SIN Grid V006 [Data set]. NASA EOSDIS Land Processes DAAC. NASA EOSDIS Land Process. DAAC 2015, 10, 415. [Google Scholar]
- Pekel, J.-F.; ACottam; Gorelick, N.; Belward, A.S. High-resolution mapping of global surface water and its long-term changes. Nature 2016, 540, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Junk, W.J.; Wittmann, F.; Schöngart, J.; Piedade, M.T.F. A classification of the major habitats of Amazonian black-water river floodplains and a comparison with their white-water counterparts. Wetl. Ecol. Manag. 2015, 23, 677–693. [Google Scholar] [CrossRef]
- Park, E.; Latrubesse, E.M. Surface water types and sediment distribution patterns at the confluence of mega rivers: The Solimões-Amazon and Negro Rivers junction. Water Resour. Res. 2015, 51, 6197–6213. [Google Scholar] [CrossRef]
- Alves, L.C.P.d.S.; Machado, C.J.S.; Vilani, R.M.; Vidal, M.D.; Andriolo, A.; Azevedo, A.D.F. As atividades turísticas baseadas na alimentação artificial de botos-da-Amazônia (Inia geoffrensis) e a legislação ambiental brasileira. Desenvolv. Meio Ambiente 2013, 28, 89–106. [Google Scholar] [CrossRef]
- Vidal, M.D.; Santos, P.M.D.C.; Chaves, M.D.P.S.R.; Swett, R. Challenges and advances in the planning of tourism with Amazon river dolphins in the Brazilian Amazon. In Tourism; Khan, S.A.R., Ed.; IntechOpen: London, UK, 2021; pp. 1–16. [Google Scholar]
- Gravena, W.; Hrbek, T.; da Silva, V.M.F.; Farias, I.P. Boto (Inia geoffrensis—Cetacea: Iniidae) aggregations in two provisioning sites in the lower Negro River—Amazonas, Brazil: Are they related? PeerJ 2019, 7, e6692. [Google Scholar] [CrossRef]
- Vidal, M.D.; Santos, P.M.D.C.; Parise, M.; Chaves, M.D.P.S.R. From food supply to contemplation: Proposition of areas for dolphin-watching tourism in the Anavilhanas National Park, Brazil. Tour. Plan. Dev. 2021, 20, 1121–1139. [Google Scholar] [CrossRef]
- Monteiro-Neto, C.; Avila, F.J.C.; Alves, T.T., Jr.; Araujo, D.S.; Campos, A.A.; Martins, A.M.A.; Parente, C.L.; Furtado-Neto, M.A.A.; Lien, J. Behavioral responses of Sotalia fluviatilis (Cetacea, Delphinidae) to acoustic pingers, Fortaleza, Brazil. Mar. Mammal Sci. 2004, 20, 145–151. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A Priori Expectations | |
| Spatial overlap |
|
| |
| Temporal overlap | High-water season: |
| |
| |
| |
| Low-water season: | |
|
| Source | Product | Band | Metric | Spatial Resolution | Temporal Resolution | Threshold |
|---|---|---|---|---|---|---|
| NASA MODIS | MOD13Q1 V6 | NDVI | Surface and Submerged Vegetation | 250-m | 16-day | 0.2 |
| NASA MODIS | MOD09Q1 V6 | Surface reflectance band 1 (red) | Suspended Sediment Concentration | 250-m | 8-day | 0.0965 |
| JRC | Global Surface Water Mapping Layers V1.2 | Occurrence | Water Body Extent | 30-m | >0.73 low-water; >0 high-water | |
| Landscan | Landscan 2016 | Population density | 1000-m |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fung, C.Y.; Peter, B.G.; Simmons, C.S. Habitat Mapping and Spatiotemporal Overlap of the Amazon River Dolphin, Fishers, and Tourism in the Central Region of the Brazilian Amazon. Conservation 2023, 3, 523-542. https://doi.org/10.3390/conservation3040034
Fung CY, Peter BG, Simmons CS. Habitat Mapping and Spatiotemporal Overlap of the Amazon River Dolphin, Fishers, and Tourism in the Central Region of the Brazilian Amazon. Conservation. 2023; 3(4):523-542. https://doi.org/10.3390/conservation3040034
Chicago/Turabian StyleFung, Cadi Y., Brad G. Peter, and Cynthia S. Simmons. 2023. "Habitat Mapping and Spatiotemporal Overlap of the Amazon River Dolphin, Fishers, and Tourism in the Central Region of the Brazilian Amazon" Conservation 3, no. 4: 523-542. https://doi.org/10.3390/conservation3040034