
Assisted Migration of Sequoiadendron Genotypes for Conservation and Timber: Performance and Morphology in a Warmer Climate Outside of Their Range

Abstract
:1. Introduction
2. Materials and Methods
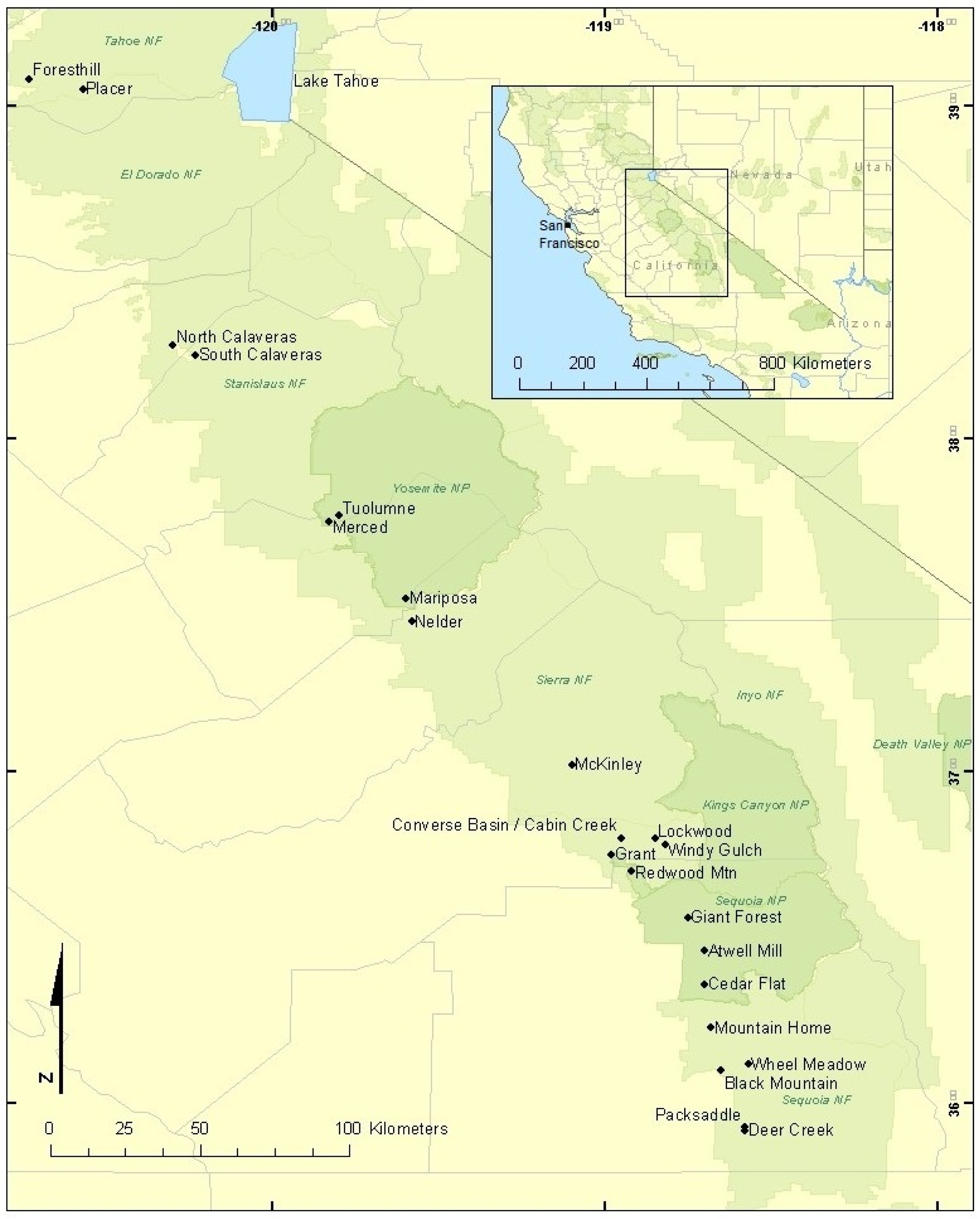
2.1. Study Site
2.2. Seed Collection and Creation of Cloned Sets
2.3. Experimental Design and Planting History
2.4. Nursery Effect and Thinning of the Trials
2.5. Data Collection and Calculations
2.6. Analysis
2.6.1. Comparison of Giant Sequoia Seedlings and Stecklings
2.6.2. Giant Sequoia Seedlings in a Mixed-Species Planting
2.6.3. Heartwood Decay Resistance
3. Results
3.1. Performance and Morphology of Steckling Grove Samples
3.1.1. Trial 1 Steckling Tree-Size Traits after 29 Growing Seasons
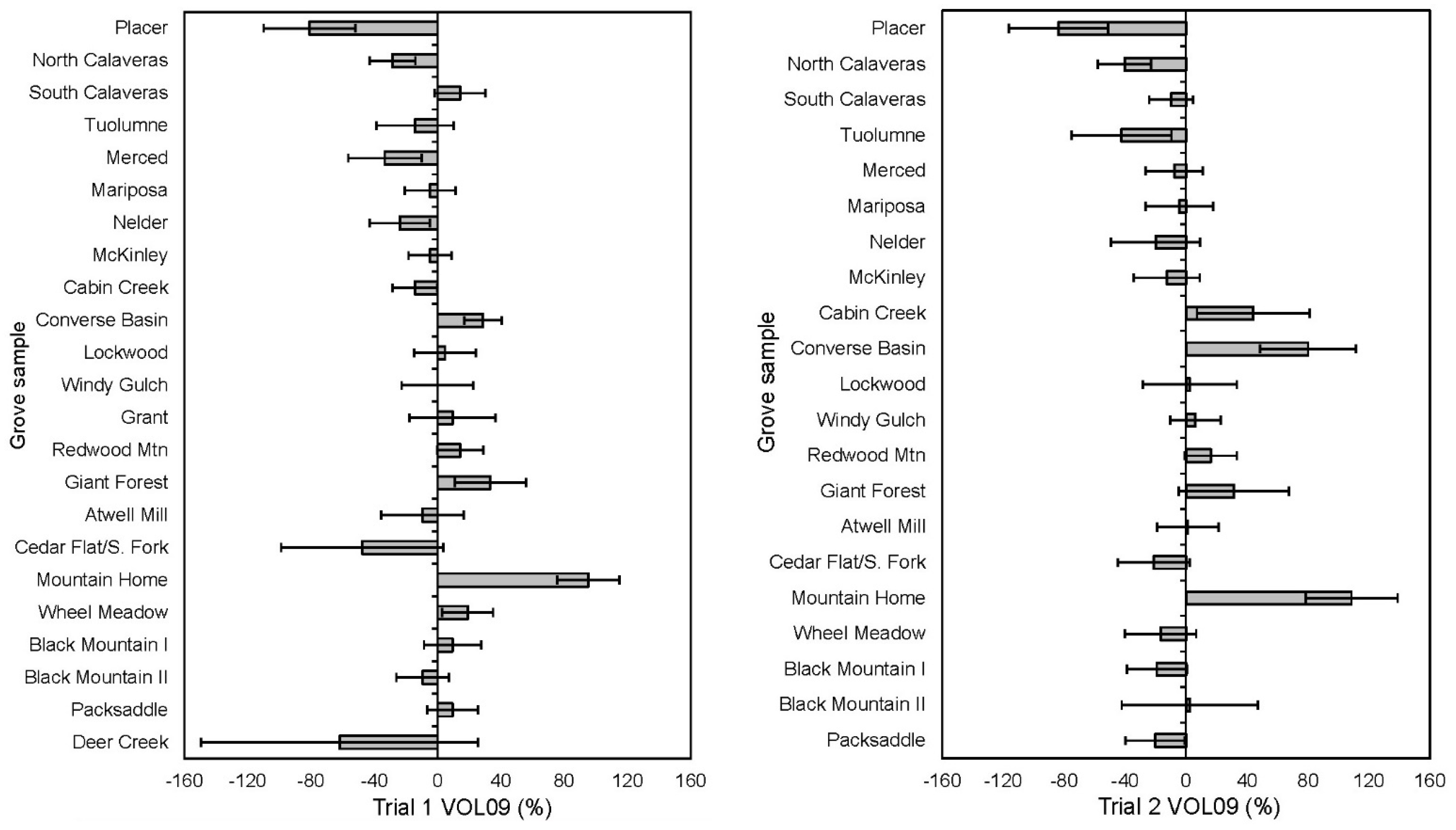
3.1.2. Trial 1 Steckling Growth over 29 Growing Seasons from 1981 to 2009
3.1.3. Trial 1 Steckling Height–Diameter Ratio from 1991 to 2009
3.1.4. Trial 1 Steckling Form Traits after 29 Growing Seasons
3.1.5. Trial 2 Steckling Tree-Size Traits after 29 Growing Seasons
3.1.6. Trial 2 Steckling Growth over Time 1981–2009
3.1.7. Trial 2 Steckling Height–Diameter Ratios from 1991 to 2009
3.1.8. Trial 2 Steckling Form Traits after 29 Growing Seasons
3.2. Comparison of Steckling Performance and Morphology in Pure vs. Mixed Planting
3.3. Seedling Performance and Morphology
3.3.1. Trial 1 Seedling Grove Sample Tree-Size Traits
3.3.2. Trial 1 Seedling Grove Sample Form Traits
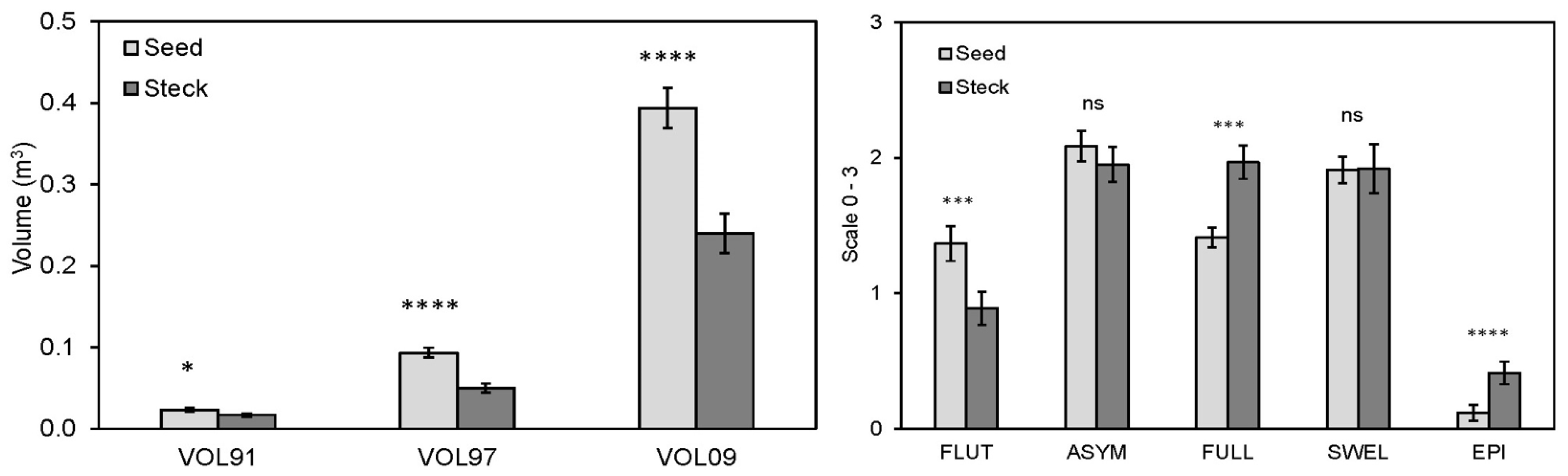
3.3.3. Trial 1 Comparative Performance and Morphology of Seedlings and Stecklings
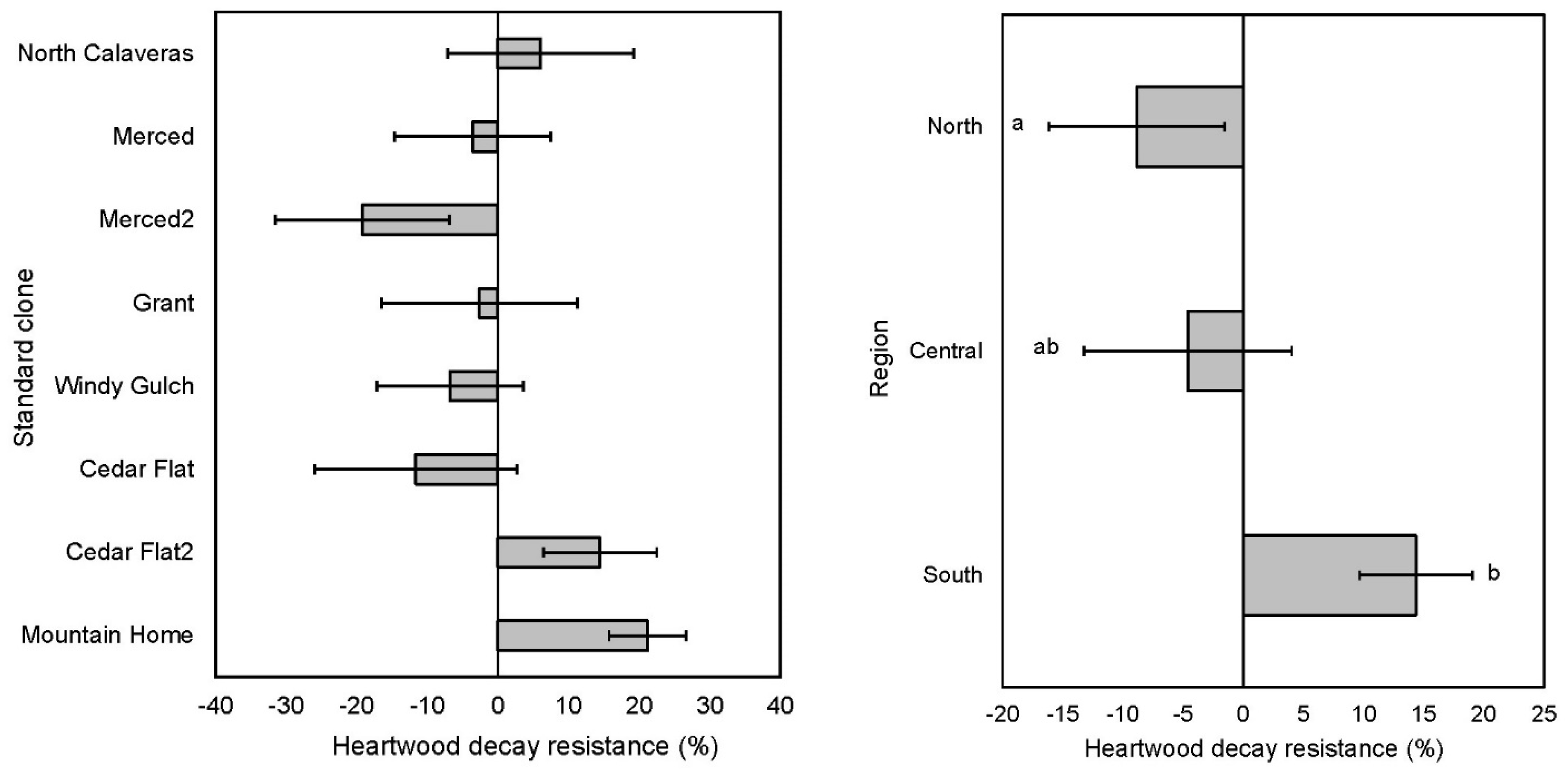
3.4. Heartwood Decay Resistance in Stumps 11 Years after Thinning
4. Discussion
4.1. Performance and Morphology Varied by Grove Origins
4.2. Growth and Tree Size Differed between Pure vs. Mixed Planting
4.3. Differences among Steckling and Seedling Tree-Size and Form Traits
4.4. Heartwood Decay Resistance of Thinned Stumps
4.5. Limitations and Recommendations for Future Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dirnböck, T.; Essl, F.; Rabitsch, W. Disproportional risk for habitat loss of high-altitude endemic species under climate change. Glob. Change Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; O’Neill, G.; Aitken, S.N.; Lindig-Cisneros, R. Assisted migration field tests in Canada and Mexico: Lessons, limitations, and challenges. Forests 2021, 12, 9. [Google Scholar] [CrossRef]
- Burdon, R.D.; Libby, W.J.; Brown, A. Domestication of Radiata Pine; Springer International: Cham, Switzerland, 2017; 480p. [Google Scholar]
- Rundel, P.W. An Annotated Check List of the Groves of Sequoiadendron giganteum in the Sierra Nevada, California. Madroño 1972, 21, 319–328. [Google Scholar]
- Harvey, H.T.; Shellhammer, H.S.; Stecker, R.E. Giant Sequoia Ecology; Scientific Monograph Series, 12; U.S. Department of Interior, National Park Service: Washington, DC, USA, 1980; 182p.
- Willard, D.A. Guide to the Sequoia Groves of California; Yosemite Association: El Portal, CA, USA, 2000; 124p. [Google Scholar]
- Hartesveldt, R.J.; Harvey, H.T.; Shellhammer, H.S.; Stecker, R.E. The Giant Sequoia of the Sierra Nevada; U.S. Department of Interior: Washington, DC, USA, 1975; 180p.
- Fins, L.; Libby, W.J. Population variation in Sequoiadendron: Seed and seedling studies, vegetative propagation, and isozyme variation. Silvae Genet. 1982, 31, 102–110. [Google Scholar]
- Libby, W.J. Genetic Variation and Early Performance of Giant Sequoia in Plantation; General Technical Report PSW-95; Pacific Southwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Berkeley, CA, USA, 1986.
- Rundel, P.W. Community structure and stability in the giant sequoia groves of the Sierra Nevada, California. Am. Midl. Nat. 1971, 85, 478–492. [Google Scholar] [CrossRef]
- DeSilva, R.; Dodd, R.S. Fragmented and isolated: Limited gene flow coupled with weak isolation by environment in the paleoendemic giant sequoia (Sequoiadendron giganteum). Am. J. Bot. 2020, 107, 45–55. [Google Scholar] [CrossRef]
- Rundel, P.W. Habitat restriction in giant sequoia: The environmental control of grove boundaries. Am. Midl. Nat. 1972, 87, 81–99. [Google Scholar] [CrossRef]
- Stephenson, N.L. Long-term dynamics of giant sequoia populations: Implications for managing a pioneer species. In Proceedings of the Symposium on Giant Sequoias: Their Place in the Ecosystem and Society; General Technical Report PSW-151; Aune, P., Ed.; Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture: Albany, CA, USA, 1994; pp. 56–63. [Google Scholar]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fule, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread increase of tree mortality rates in the western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef]
- Pile, L.S.; Meyer, M.D.; Rojas, R.; Roe, O.; Smith, M.T. Drought impacts and compounding mortality on forest trees in the southern Sierra Nevada. Forests 2019, 10, 237. [Google Scholar] [CrossRef]
- Harvey, H.T. Evolution and History of Giant Sequoia; General Technical Report PSW-95; Pacific Southwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Berkeley, CA, USA, 1986.
- Millar, C.I.; Woolfenden, W.B. Sierra Nevada Forests: Where did they come from? Where are they going? What does it mean? In Transcripts of 64th North American Wildlands and Natural Resources Conference; Wildlife Management Institute: Washington, DC, USA, 1999; pp. 206–236. [Google Scholar]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- York, R.A.; Stephenson, N.L.; Meyer, M.; Hanna, S.; Moody, T.; Carrio, T.; Battles, J.J. Giant Sequoia. In A Natural Resource Condition Assessment for Sequoia and Kings Canyon National Parks; Sydoriak, C., Panek, J.A., Battles, J.J., Nydick, K.R., Eds.; Natural Resource Report NPS/SEKI/NRR-2013/665.11a; National Park Service: Fort Collins, CO, USA, 2013; 81p. [Google Scholar]
- Harris, A.S.; Farr, W.A. The Forest Ecosystem of Alaska: Forest Ecology and Management; General Technical Report PNW-25; Pacific Northwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Portland, OR, USA, 1974; 116p.
- De La Mare, P. Sequoiadendron. In The Proceedings of a Farm Forestry Seminar on Special Purpose Tree Plantings; New Zealand Farm Forestry Association: Telford Polytechnic: Waitepeka, New Zealand, 2004; pp. 11–14. [Google Scholar]
- Weatherspoon, C. Sequoiadendron giganteum (Lindl.) Buchholz. In Silvics of North America. Vol. 1. Conifers; Burns, R.M., Honkala, B.H., Eds.; Agriculture Handbook 654; USDA Forest Service: Washington, DC, USA, 1990; pp. 552–562. Available online: http://www.na.fs.fed.us/spfo/pubs/silvics_manual/table_of_contents.htm (accessed on 9 May 2016).
- Kozlowski, T.T.; Pallardy, S.G. Physiology of Woody Plants, 2nd ed.; Academic Press, Inc.: San Diego, CA, USA, 1997; 411p. [Google Scholar]
- O’Hara, K.L.; York, R.A.; Heald, R.C. Effect of pruning severity and timing of treatment on epicormic sprout development in giant sequoia. Forestry 2008, 81, 103–110. [Google Scholar] [CrossRef]
- Kitzmiller, J.H.; Lunak, G. Growth of giant sequoia compared to ponderosa pine and other mixed-conifers in California plantations. West. J. Appl. For. 2012, 27, 196–204. [Google Scholar] [CrossRef]
- Piirto, D.D. Wood of Giant Sequoia: Properties and Unique Characteristics; General Technical Report PSW-95; Pacific Southwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Berkeley, CA, USA, 1986.
- Ukrainetz, N.K.; Kang, K.-Y.; Aitken, S.N.; Stoehr, M.; Mansfield, S.D. Heritability and phenotypic and genetic correlations of coastal Douglas-fir (Pseudotsuga menziesii) wood quality traits. Can. J. For. Res. 2008, 38, 1536–1546. [Google Scholar] [CrossRef]
- WRCC. Western Regional Climate Center: Cooperative Climatological Data Summaries. 2014. Available online: http://www.wrcc.dri.edu/climatedata/climsum/ (accessed on 23 February 2014).
- Thornton, P.E.; Thornton, M.M.; Mayer, B.W.; Wilhelmi, N.; Wei, Y.; Cooke, R.B. Daymet: Daily Surface Weather on a 1 km Grid for North America 1980–2011; Oak Ridge National Laboratory Distributed Active Archive Center: Oak Ridge, TN, USA, 2013. [CrossRef]
- Fins, L. Genetic Architecture of Giant Sequoia. Ph.D. Dissertation, University of California, Berkeley, CA, USA, 1979; 237p. [Google Scholar]
- Mahalovich, M.F. A Genetic Architecture Study of Giant Sequoia: Early Growth Characteristics. Master’s Thesis, University of California, Berkeley, CA, USA, 1985; 98p. [Google Scholar]
- Libby, W.J.; Cockerham, C.C. Random non-contiguous plots in interlocking field layouts. Silvae Genet. 1980, 29, 183–190. [Google Scholar]
- Harry, D.E. Shoot elongation and growth plasticity in incense-cedar. Can. J. For. Res. 1987, 17, 484–489. [Google Scholar] [CrossRef]
- Conover, W.J.; Johnson, M.E.; Johnson, M.M. A comparative study of homogeneity of variances, with application to the outer continental shelf bidding data. Technometrics 1981, 23, 351–361. [Google Scholar] [CrossRef]
- York, R.A.; O’Hara, K.L.; Battles, J.J. Density effects on giant sequoia (Sequoiadendron giganteum) growth through 22 years: Implications for restoration and plantation management. West. J. Appl. For. 2013, 28, 30–36. [Google Scholar] [CrossRef]
- Cox, L.E.; York, R.A.; Battles, J.J. Growth and form of giant sequoia (Sequoiadendron giganteum) in a plantation spacing trial after 28 years. For. Ecol. Manag. 2021, 488, 119033. [Google Scholar] [CrossRef]
- Sexton, J.P.; Hangartner, S.B.; Hoffmann, A.A. Genetic isolation by environment or distance: Which pattern of gene flow is most common? Evolution 2014, 68, 1–15. [Google Scholar] [CrossRef]
- Pretzsch, H.; Grams, T.; Häberle, K.H.; Pritsch, K.; Bauerle, T.; Rötzer, T. Growth and mortality of Norway spruce and European beech in monospecific and mixed-species stands under natural episodic and experimentally extended drought. Results of the KROOF throughfall exclusion experiment. Trees 2020, 34, 957–970. [Google Scholar] [CrossRef]
- Rydelius, J.A.; Libby, W.J. Arguments for clonal redwood forestry. In Clonal Forestry II; Ahuja, M.A., Libby, W.J., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; pp. 158–168. [Google Scholar]
- Deal, R.L.; Barbour, R.J.; McClellan, M.H.; Parry, D.L. Development of epicormic sprouts in Sitka spruce following thinning and pruning in south-east Alaska. West. J. Appl. For. 2003, 76, 401–412. [Google Scholar] [CrossRef]
- DeBell, J.D.; Gartner, B.L. Stem characteristics on the lower log of 35-year-old western redcedar grown at several spacings. West. J. Appl. For. 1997, 12, 9–15. [Google Scholar] [CrossRef]
- Julin, K.R.; Shaw, C.G., III; Farr, W.A.; Hinckley, T.M. The fluted western hemlock of Alaska. I. Morphological studies and experiments. For. Ecol. Manag. 1993, 60, 119–132. [Google Scholar] [CrossRef]
- Julin, K.R.; Shaw III, C.G.; Farr, W.A.; Hinckley, T.M. The fluted western hemlock of Alaska. II. Stand observations and synthesis. For. Ecol. Manag. 1993, 60, 133–141. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Berrill, J.-P. Epicormic sprout development in pruned coast redwood: Pruning severity, genotype, and sprouting characteristics. Ann. For. Sci. 2009, 66, 409. [Google Scholar] [CrossRef]
- Waring, K.M.; O’Hara, K.L. Ten-year growth and epicormic sprouting response of western larch to pruning in western Montana. West. J. Appl. For. 2005, 20, 228–232. [Google Scholar] [CrossRef]
- Kellogg, R.M.; Barber, F.J. Stem eccentricity in coastal western hemlock. Can. J. For. Res. 1981, 11, 715–718. [Google Scholar] [CrossRef]
- Paques, L.E. Genetic control of heartwood content in larch. Silvae Genet. 2001, 50, 69–75. [Google Scholar]
- Ahuja, M.R. Genetic constitution and diversity in four narrow endemic redwoods from the family Cupressaceae. Euphytica 2008, 165, 5–19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height (m) | Diameter (cm) | Volume (m3) | HDR | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grove | n | Mean | s.d. | Max | Min | R | Mean | s.d. | Max | Min | R | Mean | s.d. | Max | Min | R | Mean | s.d. | Max | Min | R |
| Placer | 8 | 6.2 | 2.1 | 9.9 | 3.6 | 22 | 14.8 | 3.9 | 20.2 | 9.4 | 23 | 0.04 | 0.03 | 0.11 | 0.01 | 23 | 42 | 4.2 | 49 | 38 | 2 |
| N. Calaveras | 19 | 8.9 | 2.6 | 15.9 | 3.8 | 19 | 23.3 | 5.2 | 32.8 | 9.7 | 20 | 0.15 | 0.09 | 0.45 | 0.01 | 19 | 38 | 3.7 | 49 | 33 | 10 |
| S. Calaveras | 13 | 10.1 | 2.4 | 13.6 | 6.4 | 11 | 28.4 | 5.7 | 38.2 | 21.1 | 4 | 0.24 | 0.14 | 0.47 | 0.07 | 5 | 35 | 4.1 | 43 | 30 | 16 |
| Tuolumne | 12 | 9.3 | 3.1 | 15.2 | 5.8 | 16 | 24.6 | 6.9 | 35.2 | 14.8 | 18 | 0.18 | 0.15 | 0.49 | 0.03 | 16 | 38 | 4.9 | 44 | 31 | 12 |
| Merced | 10 | 7.6 | 2.3 | 11.3 | 4.2 | 20 | 24.2 | 6.3 | 33.1 | 13.5 | 19 | 0.14 | 0.10 | 0.32 | 0.02 | 20 | 31 | 3.4 | 36 | 26 | 23 |
| Mariposa | 8 | 9.7 | 1.6 | 11.8 | 7.3 | 13 | 26.9 | 4.7 | 34.4 | 18.4 | 11 | 0.20 | 0.09 | 0.37 | 0.07 | 12 | 36 | 4.6 | 42 | 26 | 15 |
| Nelder | 6 | 9.0 | 1.8 | 10.5 | 6.3 | 18 | 25.3 | 4.0 | 31.2 | 20.2 | 16 | 0.16 | 0.07 | 0.27 | 0.07 | 18 | 35 | 3.7 | 40 | 31 | 16 |
| McKinley | 13 | 10.0 | 2.6 | 15.3 | 5.8 | 12 | 26.7 | 3.5 | 33.2 | 20.3 | 13 | 0.20 | 0.10 | 0.44 | 0.06 | 12 | 37 | 6.8 | 52 | 29 | 14 |
| Cabin Creek | 10 | 9.4 | 1.8 | 11.8 | 5.4 | 15 | 26.6 | 3.7 | 32.8 | 20.5 | 14 | 0.18 | 0.08 | 0.33 | 0.06 | 16 | 35 | 4.2 | 40 | 26 | 16 |
| Converse Basin | 14 | 11.7 | 1.8 | 15.5 | 9.3 | 2 | 29.0 | 4.1 | 37.4 | 23.7 | 2 | 0.27 | 0.12 | 0.57 | 0.14 | 3 | 41 | 3.5 | 46 | 34 | 3 |
| Lockwood | 11 | 10.4 | 2.9 | 14.4 | 6.8 | 9 | 26.7 | 6.3 | 36.2 | 18.3 | 12 | 0.22 | 0.14 | 0.48 | 0.06 | 10 | 39 | 4.8 | 48 | 32 | 6 |
| Windy Gulch | 5 | 10.3 | 1.7 | 12.8 | 8.4 | 10 | 27.1 | 4.4 | 34.0 | 22.2 | 10 | 0.21 | 0.11 | 0.39 | 0.11 | 11 | 38 | 2.9 | 41 | 34 | 8 |
| Grant | 7 | 9.2 | 2.6 | 14.2 | 6.6 | 17 | 28.8 | 6.5 | 38.9 | 18.3 | 3 | 0.23 | 0.17 | 0.56 | 0.06 | 7 | 32 | 4.9 | 39 | 26 | 21 |
| Redwood Mtn | 19 | 10.6 | 3.3 | 15.9 | 4.6 | 6 | 27.5 | 6.0 | 35.5 | 14.8 | 8 | 0.24 | 0.15 | 0.52 | 0.03 | 5 | 38 | 5.9 | 46 | 30 | 9 |
| Giant Forest | 11 | 11.1 | 3.1 | 16.3 | 6.3 | 3 | 28.4 | 8.0 | 39.1 | 16.0 | 5 | 0.28 | 0.21 | 0.65 | 0.04 | 2 | 39 | 4.0 | 45 | 33 | 5 |
| Atwell Mill | 10 | 9.6 | 3.2 | 15.0 | 4.1 | 14 | 24.7 | 7.8 | 37.0 | 10.9 | 17 | 0.19 | 0.16 | 0.47 | 0.01 | 14 | 39 | 4.8 | 45 | 32 | 7 |
| Cedar Flat | 3 | 7.2 | 3.2 | 10.6 | 4.4 | 21 | 22.2 | 6.9 | 28.0 | 14.6 | 21 | 0.11 | 0.10 | 0.22 | 0.02 | 21 | 32 | 5.5 | 38 | 27 | 22 |
| Mtn. Home | 11 | 12.0 | 3.1 | 17.1 | 8.0 | 1 | 33.8 | 6.8 | 47.9 | 25.9 | 1 | 0.41 | 0.27 | 1.03 | 0.14 | 1 | 35 | 3.8 | 42 | 30 | 16 |
| Wheel Meadow | 13 | 10.6 | 3.1 | 15.3 | 5.6 | 6 | 27.9 | 5.9 | 37.0 | 16.5 | 6 | 0.25 | 0.14 | 0.48 | 0.04 | 4 | 38 | 6.0 | 51 | 28 | 13 |
| Black Mtn. I | 13 | 10.8 | 2.8 | 16.7 | 7.6 | 4 | 27.4 | 4.9 | 36.7 | 20.9 | 9 | 0.23 | 0.15 | 0.59 | 0.09 | 7 | 40 | 7.2 | 51 | 28 | 4 |
| Black Mtn. II | 8 | 10.7 | 2.1 | 12.8 | 7.4 | 5 | 25.3 | 4.0 | 30.3 | 20.4 | 15 | 0.19 | 0.09 | 0.31 | 0.09 | 14 | 42 | 3.5 | 47 | 35 | 1 |
| Packsaddle | 11 | 10.5 | 2.4 | 15.1 | 6.3 | 8 | 27.7 | 4.6 | 36.3 | 17.9 | 7 | 0.23 | 0.12 | 0.52 | 0.05 | 7 | 38 | 3.9 | 43 | 33 | 10 |
| Deer Creek | 2 | 6.2 | 3.4 | 8.6 | 3.8 | 22 | 18.9 | 10.3 | 26.1 | 11.6 | 22 | 0.08 | 0.10 | 0.15 | 0.01 | 22 | 33 | 0.1 | 33 | 33 | 20 |
| All | 237 | 9.9 | 2.8 | 17.1 | 3.6 | 26.5 | 6.3 | 47.9 | 9.4 | 0.21 | 0.15 | 1.03 | 0.01 | 37 | 5.3 | 52 | 26 | ||||
| Decay | DBH97 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Grove | n | Mean | s.d. | Max | Min | %mean | Mean | s.d. | Max | Min | %mean |
| Seedlings | |||||||||||
| N. Calaveras | 25 | 2.1 | 0.6 | 3 | 1 | 101 | 18.3 | 3.5 | 25 | 12 | 91 |
| Redwood Mtn. | 21 | 2.2 | 0.7 | 3 | 1 | 106 | 21.0 | 2.5 | 26 | 17 | 104 |
| Giant Forest | 22 | 1.9 | 0.8 | 3 | 0 | 94 | 20.0 | 5.0 | 29 | 11 | 99 |
| Cedar Flat | 17 | 2.1 | 0.8 | 3 | 0 | 102 | 20.8 | 4.2 | 29 | 10 | 103 |
| Garfield | 26 | 2.1 | 0.7 | 3 | 1 | 104 | 19.9 | 3.9 | 30 | 13 | 98 |
| Mountain Home | 28 | 1.9 | 0.7 | 3 | 1 | 93 | 21.2 | 4.1 | 31 | 14 | 105 |
| All | 2.1 | 0.7 | 3 | 0 | 20.3 | 4.0 | 31 | 10 | 100 | ||
| Clones | |||||||||||
| N. Calaveras | 10 | 2.1 | 0.9 | 3 | 1 | 106 | 16.0 | 1.6 | 19 | 13 | 98 |
| Merced | 11 | 1.9 | 0.7 | 3 | 1 | 96 | 14.5 | 2.7 | 19 | 10 | 89 |
| Merced 2 | 20 | 1.6 | 0.9 | 3 | 0 | 81 | 18.3 | 3.7 | 27 | 8 | 112 |
| Grant | 14 | 1.9 | 1.0 | 3 | 0 | 97 | 12.9 | 3.6 | 22 | 12 | 79 |
| Windy Gulch | 13 | 1.8 | 0.7 | 3 | 1 | 93 | 15.9 | 3.6 | 26 | 11 | 98 |
| Cedar Flat | 4 | 1.8 | 0.5 | 2 | 1 | 88 | 15.3 | 2.6 | 19 | 11 | 94 |
| Cedar Flat 2 | 15 | 2.3 | 0.7 | 3 | 1 | 114 | 17.5 | 4.6 | 27 | 8 | 107 |
| Mountain Home | 15 | 2.4 | 0.5 | 3 | 2 | 121 | 17.7 | 4.9 | 25 | 6 | 109 |
| All | 2.0 | 0.8 | 3 | 0 | 16.3 | 4.1 | 27 | 10 | 100 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valness, C.M.; Libby, W.J.; Berrill, J.-P. Assisted Migration of Sequoiadendron Genotypes for Conservation and Timber: Performance and Morphology in a Warmer Climate Outside of Their Range. Conservation 2023, 3, 543-568. https://doi.org/10.3390/conservation3040035
Valness CM, Libby WJ, Berrill J-P. Assisted Migration of Sequoiadendron Genotypes for Conservation and Timber: Performance and Morphology in a Warmer Climate Outside of Their Range. Conservation. 2023; 3(4):543-568. https://doi.org/10.3390/conservation3040035
Chicago/Turabian StyleValness, Christopher M., William J. Libby, and John-Pascal Berrill. 2023. "Assisted Migration of Sequoiadendron Genotypes for Conservation and Timber: Performance and Morphology in a Warmer Climate Outside of Their Range" Conservation 3, no. 4: 543-568. https://doi.org/10.3390/conservation3040035