
Heterologous Expression of Recombinant Human Cytochrome P450 (CYP) in Escherichia coli: N-Terminal Modification, Expression, Isolation, Purification, and Reconstitution

Abstract
:1. Introduction
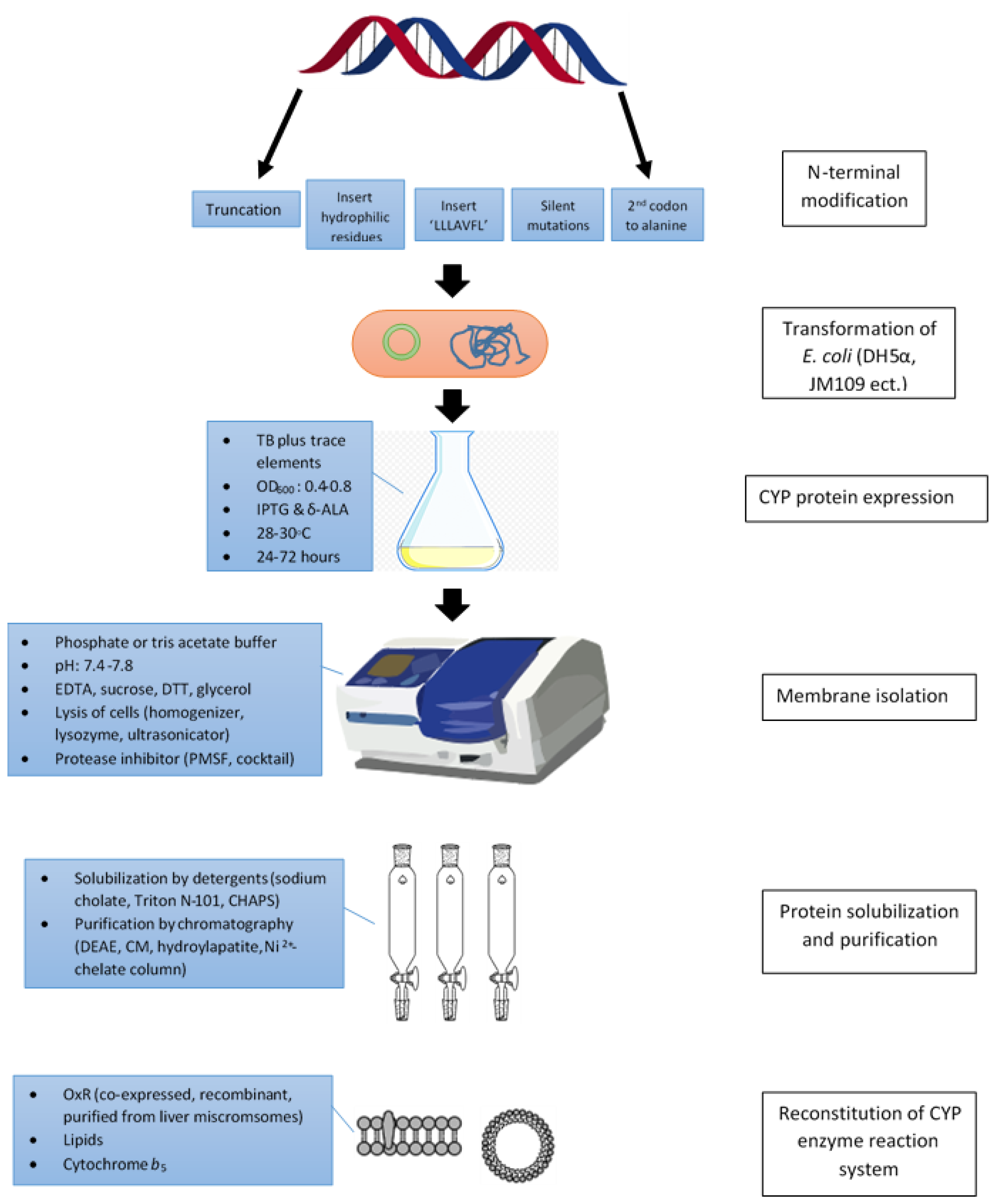
2. Modifications of N-Terminus

2.1. N-Terminal Trucation
2.2. Substitutions of N-Terminal Sequences with Hydrophilic Residues
2.3. Incorporation of the ‘LLLAVFL’ Sequence
2.4. Silent Mutations
2.5. Use Codons Encoding Alanine as the Second Codon
3. Co-Expression with Chaperon
4. Selections of Expression Vectors and E. coli Strains

{kind=link}
{kind=link}
| CYP | Expression Vector | E. coli Strain | LB to TB Ratio | OD600 | Temp (°C) | Shaking Speed (rpm) | Duration (hour) | IPTG (mM) | Δ-ALA (mM) | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| 2E1 | pCWori+ | MV1304 | 1:90 | 0.5–0.8 | N/A | N/A | 4 | 1 | N/A | [7] |
| 17A1 | pCWori+ | JM109 | N/A | 0.4–0.8 | 28 | N/A | 48 | 1 | N/A | [25] |
| 1A2 | pCWori+ | DH5α | 1:100 | N/A | 30 | 125 | 72 | 1 | N/A | [66] |
| 3A4 | pCWori+ | DH5α | 1:100 | N/A | 32 | N/A | 24 | 1 | N/A | [42] |
| 2E1&2B4 | pJL | MV1304 | N/A | 1.0 | N/A | N/A | 4 | 1 | N/A | [43] |
| 1A1 | pCWori+ | DH5α | 1:100 | N/A | 30 | Vigorous | 48 | N/A | N/A | [44] |
| 2E1 | pCWori+ | DH5α | 1:100 | N/A | 30 | Vigorous | 48 | 1 | N/A | [68] |
| 1A2 | pCWori+ | DH5α or JM109 | 1:100 | N/A | 30 | Vigorous | 48 | 1 | N/A | [38] |
| 7A1 | pJL | TOPP3 | N/A | 0.4–0.6 | 30 | 150 | 15–18 | 1 | 0.2 | [83] |
| 17A1-OxR | pCWori+ | DH5α | 1:111 | N/A | 27 | 125 | 72 | 1 | N/A | [69] |
| 3A5 | pCWori+ | DH5α | 1:100 | N/A | 30 | 220 | 24 | 1 | N/A | [70] |
| 2D6 | pDS9 | JM109 | 1:10 to 40 | 0.7–0.9 | 23 | 100 | 48 | 5 µM | 0.5–1 | [77] |
| 2D6 | pCWori+ | DH5α | N/A | N/A | 30 | 200 | 43 | 1.5 | 0.5 | [39] |
| 2E1-OxR | pJL2 | XL Blue | N/A | 0.8 | 26 | 100 | 60 | 1 | N/A | [82] |
| 1A1-OxR | pCWori+ | DH5α | 1:100 | N/A | 28 | 125 | 48 | 1 | 0.5 | [71] |
| 27A1 | pTrc99A | TOPP3 | 1:100 | N/A | 29 | 210 | 48 | 0.5 | 0.5 | [84] |
| 1A2-OxR | pCWori+ | DH5α | 1:100 | N/A | 28 | 125 | 48 | 1 | 0.5 | [85] |
| 1B1-OxR | pCWori+ | DH5α | 1:100 | N/A | 30 | 200 | 24 | 1 | 0.5 | [45] |
| 3A5 | pCWori+ | DH5α | 1:100 | 0.3 | 30 | 160 | 28 | 0.1 | 1 | [72] |
| 2A6 | pCWori+ | DH5α | 1:100 | N/A | 32 | 200 | 40 | 1 | 0.5 | [46] |
| 2B6 | pCWori+ | MV1304 | N/A | N/A | 28 | 200 | 40–48 | 1 | 0.5 | [47] |
| 2D6-OxR | pCWori+ | DH5α | 1:100 | 0.6–1.0 | 26 | 190 | 40–48 | 1 | 0.5 | [27] |
| 1A2-HDJ-1 | pCWori+ | DH5α | N/A | 0.4–0.5 | 37 | N/A | 24 | 1 | 1.5 | [26] |
| 2B6-GroES/EL | pCWori+ | JM109 | 1:100 | N/A | 30 | 160 | 72 | 1 | 0.5 | [64] |
| 27C1 | pCWori+ | JM109 | 1:100 | N/A | 27 | 200 | 48 | 1 | 0.5 | [79] |
| 4X1 | pCWori+ | DH5α | 1:100 | 0.5 | 28 | 190 | 17–21 | 1 | 0.5 | [52] |
| 2S1 | pBdtacHR | LMG194 | 1:400 | N/A | 30 | 115 | 24–36 | 0.5 | 0.5 | [48] |
| 1A1-OxR | pCWori+ | DH5α | 1:1000 | 0.5–0.7 | 30 | 200 | 24 | 1 | 0.5 | [8] |
| 2C10 &2C9 | pCWori+ | DH5α and JM109 | 1:100 | N/A | 30 | Vigorous | 24 | 1 | N/A | [24] |
| 4A11 | pCWori+ | DH5α | N/A | 0.5 | 28 | 200 | 48 | 1 | 0.5 | [74] |
| 2J2 | pCWori+ | DH5α | N/A | N/A | 28 | N/A | 48 | 1 | 0.5 | [75] |
| 4B1 | pCWori+ | DH5α | N/A | 0.4 | 27 | 120 | 48 | 1 | 0.5 | [76] |
| 39A1-GroEL/ES | pCW-LIC | C41 | N/A | 0.6 | 26 | 110 | 48 | 0.5 | 0.5 | [86] |
| 2J2-GroEL/ES | pCWori+ | DH5α | 1:100 | 0.4–0.6 | 28 | 180 | 48 | 1 | 0.5 | [9] |
5. Bacteria Culture and Protein Expression Conditions
6. Membrane Isolation
7. CYP Protein Solubilization and Purification
8. Reconstitution of CYP Enzyme Assay Systems In Vitro
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, L.; Wang, H.; Chen, K.; Li, Y. Biomedicine & Pharmacotherapy Roles of hydroxyeicosatetraenoic acids in diabetes (HETEs and diabetes). Biomed. Pharmacother. 2022, 156, 113981. [Google Scholar] [PubMed]
- Pikuleva, I.A.; Cartier, N. Cholesterol Hydroxylating Cytochrome P450 46A1: From Mechanisms of Action to Clinical Applications. Front. Aging Neurosci. 2021, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fujino, C.; Sanoh, S.; Katsura, T. Recent Advances in the Understanding of Nuclear Receptors- and Drug-Metabolizing Enzymes-Mediated Inter-Individual Differences Variation in Expression of Cytochrome P450 3A Isoforms and Toxicological Effects: Endo- and Exogenous Substances as Regulatory. Biol. Pharm. Bull. 2021, 44, 1617–1634. [Google Scholar] [CrossRef]
- Perepechaeva, M.L.; Grishanova, A.Y. The Role of CYP3A in Health and Disease. Biomedicines 2022, 10, 2686. [Google Scholar]
- Luo, B.I.N.; Yan, D.; Yan, H.; Yuan, J. Cytochrome P450: Implications for human breast cancer (Review). Oncol. Lett. 2021, 22, 548. [Google Scholar] [CrossRef]
- Gonzalez, F.J.; Nebert, D.W. Evolution of the P450 gene superfamily: Animal-plant “warfare”, molecular drive and human genetic differences in drug oxidation. Trends Genet. 1990, 6, 182–186. [Google Scholar] [CrossRef]
- Larson, J.R.; Coon, M.J.; Porter, T.D. Purification and properties of a shortened form of cytochrome P-450 2E1: Deletion of the NH2-terminal membrane-insertion signal peptide does not alter the catalytic activities. Proc. Natl. Acad. Sci. USA 1991, 88, 9141–9145. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, S.; Dong, M.S.; Park, J.D.; Park, J.; Rhee, S.; Ryu, D.Y. Characterization of the Ala62Pro polymorphic variant of human cytochrome P450 1A1 using recombinant protein expression. Toxicol. Appl. Pharmacol. 2015, 285, 159–169. [Google Scholar] [CrossRef]
- Jeong, D.; Park, H.; Lim, Y.; Lee, Y.; Kim, V.; Cho, M.; Kim, D. Drug Metabolism and Pharmacokinetics Terfenadine metabolism of human cytochrome P450 2J2 containing genetic variations (G312R, P351L and P115L). Drug Metab. Pharmacokinet. 2018, 33, 61–66. [Google Scholar] [CrossRef]
- Van Der Wouden, C.H.; Van Rhenen, M.H.; Jama, W.O.M.; Ingelman-sundberg, M.; Lauschke, V.M.; Konta, L.; Schwab, M.; Swen, J.J.; Guchelaar, H. Development of the PGx-Passport: A Panel of Actionable Germline Genetic Variants for Emptive Pharmacogenetic Testing. Clin. Pharmacokinet. 2019, 106, 866–873. [Google Scholar]
- Zhou, S.; Di, Y.M.; Chan, E.; Du, Y.; Chow, V.D.; Xue, C.C.; Lai, X.; Wang, J.; Li, C.G.; Tian, M.; et al. Clinical Pharmacogenetics and Potential Application in Personalized Medicine. Curr. Drug Metab. 2008, 9, 738–784. [Google Scholar] [CrossRef]
- Fujikura, K.; Ingelman-sundberg, M.; Lauschke, V.M. Genetic variation in the human cytochrome P450 supergene family. Pharmacogenet. Genom. 2015, 25, 584–594. [Google Scholar] [CrossRef]
- Kumondai, M.; Hishinuma, E.; Marie, E.; Rico, G.; Ito, A. Heterologous expression of high-activity cytochrome P450 in mammalian cells. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Iratsuka, M.H. Review In Vitro Assessment of the Allelic Variants of Cytochrome P450. Drug Metab. Pharmacokinet. 2012, 27, 68–84. [Google Scholar] [CrossRef]
- Taimi, M.; Helvig, C.; Wisniewski, J.; Ramshaw, H.; White, J.; Amad, M.; Korczak, B.; Petkovich, M. A Novel Human Cytochrome P450, CYP26C1, Involved in Metabolism of 9-cis and All-trans Isomers of Retinoic Acid. J. Biol. Chem. 2004, 279, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, V.R.; Siddique, M.U.M.; Gatchie, L.; Williams, I.S.; Bharate, S.B.; Jayaprakash, V.; Sinha, B.N.; Chaudhuri, B. CYP enzymes, expressed within live human suspension cells, are superior to widely-used microsomal enzymes in identifying potent CYP1A1/CYP1B1 inhibitors: Identification of quinazolinones as CYP1A1/CYP1B1 inhibitors that efficiently reverse B[a]P toxicity. Eur. J. Pharm. Sci. 2019, 131, 177–194. [Google Scholar] [CrossRef]
- Schroer, K.; Kittelmann, M.; Lütz, S. Recombinant human cytochrome P450 monooxygenases for drug metabolite synthesis. Biotechnol. Bioeng. 2010, 106, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Kusano, K.; Sakaguchi, M.; Kagawa, N.; Waterman, M.R.; Omura, T. Microsomal P450s use specific proline-rich sequences for efficient folding, but not for maintenance of the folded structure. J. Biochem. 2001, 129, 259–269. [Google Scholar] [CrossRef]
- Miyauchi, Y.; Kimura, A.; Sawai, M.; Fujimoto, K.; Hirota, Y.; Tanaka, Y.; Takechi, S.; Mackenzie, P.I.; Ishii, Y. Use of a Baculovirus-Mammalian Cell Expression-System for Expression of Drug-Metabolizing Enzymes: Optimization of Infection With a Focus on Cytochrome P450 3A4. Front. Pharmacol. 2022, 13, 832931. [Google Scholar] [CrossRef]
- Imaoka, S.; Yamada, T.; Hiroi, T.; Hayashi, K.; Sakaki, T.; Yabusaki, Y.; Funae, Y. Multiple forms of human P450 expressed in Saccharomyces cerevisiae systematic characterization and comparison with those of the rat. Biochem. Pharmacol. 1996, 51, 1041–1050. [Google Scholar] [CrossRef]
- Yasumori, T. Expression of a Human Cytochrome P450 Form in Schizosaccharomyces pombe: Comparison with Expression in Saccharomyces cerevisiae. In Foreign Gene Expression in Fission Yeast: Schizosaccharomyces pombe; Springer: Berlin/Heidelberg, Germany, 1997; pp. 111–121. [Google Scholar]
- Cheng, J.; Wan, D.F.; Gu, J.R.; Gong, Y.; Yang, S.L.; Hao, D.C.; Yang, L. Establishment of a yeast system that stably expresses human cytochrome P450 reductase: Application for the study of drug metabolism of cytochrome P450s in vitro. Protein Expr. Purif. 2006, 47, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Zelasko, S.; Palaria, A.; Das, A. Optimizations to achieve high-level expression of cytochrome P450 proteins using Escherichia coli expression systems. Protein Expr. Purif. 2013, 92, 77–87. [Google Scholar] [CrossRef]
- Sandhu, P.; Baba, T.; Guengerich, F.P. Expression of modified cytochrome P450 2C10 (2C9) in Escherichia Coli, purification, and reconstitution of catalytic activity. Arch. Biochem. Biophys. 1993, 306, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Barnes, H.J.; Arlotto, M.P.; Waterman, M.R. Expression and enzymatic activity of recombinant cytochrome P450 17α-hydroxylase in Escherichia coli. Proc. Natl. Acad. Sci. USA 1991, 88, 5597–5601. [Google Scholar] [CrossRef] [PubMed]
- Ahn, T.; Yang, S.; Yun, C.H. High-level expression of human cytochrome P450 1A2 by co-expression with human molecular chaperone HDJ-1(Hsp40). Protein Expr. Purif. 2004, 36, 48–52. [Google Scholar] [CrossRef]
- Deeni, Y.Y.; Paine, M.J.I.; Ayrton, A.D.; Clarke, S.E.; Chenery, R.; Wolf, C.R. Expression, purification, and biochemical characterization of a human cytochrome P450 CYP2D6-NADPH cytochrome P450 reductase fusion protein. Arch. Biochem. Biophys. 2001, 396, 16–24. [Google Scholar] [CrossRef]
- Black, D.; Comprehensive, T. Membrane topology of the mammalian P450 cytochromes. FASEB J. 1992, 6, 680–685. [Google Scholar] [CrossRef]
- Otyepka, M.; Skopalík, J.; Anzenbacherová, E.; Anzenbacher, P. What common structural features and variations of mammalian P450s are known to date? Biochim. Biophys. Acta 2007, 1770, 376–389. [Google Scholar] [CrossRef]
- Mustafa, G.; Nandekar, P.P.; Camp, T.J.; Bruce, N.J.; Gregory, M.C.; Sligar, S.G.; Wade, R.C. Influence of Transmembrane Helix Mutations on Cytochrome P450-Membrane Interactions and Function. Biophys. J. 2019, 116, 419–432. [Google Scholar] [CrossRef]
- Schoch, G.A.; Yano, J.K.; Wester, M.R.; Griffin, K.J.; Stout, C.D.; Johnson, E.F. Structure of Human Microsomal Cytochrome P450 2C8. J. Biol. Chem. 2004, 279, 9497–9503. [Google Scholar] [CrossRef]
- Martin, Š.; Navrátilová, V.; Paloncýová, M.; Bazgier, V.; Berka, K.; Anzenbacher, P.; Otyepka, M. Membrane-attached mammalian cytochromes P450: An overview of the membrane’s effects on structure, drug binding, and interactions with redox partners. J. Inorg. Biochem. 2018, 183, 117–136. [Google Scholar]
- Vergères, G.; Winterhalter, K.H.; Richter, C. Localization of the N-terminal methionine of rat liver cytochrome P-450 in the lumen of the endoplasmic reticulum. Biochim. Biophys. Acta (BBA)-Biomembr. 1991, 1063, 235–241. [Google Scholar] [CrossRef]
- Gideon, D.A.; Kumari, R.; Lynn, A.M.; Manoj, K.M. What is the Functional Role of N-terminal Transmembrane Helices in the Metabolism Mediated by Liver Microsomal Cytochrome P450 and its Reductase? Cell Biochem. Biophys. 2012, 63, 35–45. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Tomiyoshi, R.; Kuroiwa, T.; Mihara, K.; Omura, T. Functions of signal and signal-anchor sequences are determined by the balance between the hydrophobic segment and the N-terminal charge. Proc. Natl. Acad. Sci. USA 1992, 89, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Powers, T.; Walter, P. Co-translational protein targeting catalyzed by the Escherichia coli signal recognition particle and its receptor. EMBO J. 1997, 16, 4880–4886. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, M.P.; Ossetian, R.; Li, D.N.; Henderson, C.J.; Burchell, B.; Wolf, C.R.; Friedberg, T. A general strategy for the expression of recombinant human cytochrome P450s in Escherichia coli using bacterial signal peptides: Expression of CYP3A4, CYP2A6, and CYP2E1. Arch. Biochem. Biophys. 1997, 345, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, P.; Guo, Z.; Baba, T.; Martin, M.V.; Tukey, R.H.; Guengerich, F.P. Expression of Modified Human Cytochrome P450 1A2 in Escherichia coli: Stabilization, Purification, Spectral Characterization, and Catalytic Activities of the Enzyme. Arch. Biochem. Biophys. 1994, 309, 168–177. [Google Scholar] [CrossRef]
- Gillam, E.M.J.; Zuyu, G.; Martin, M. V Expression of cytochrome P450 2D6 in Escherichia coli, purification, and spectral and catalytic characterization. Arch. Biochem. Biophys. 1995, 319, 540–550. [Google Scholar] [CrossRef]
- Williams, P.A.; Cosme, J.; Vinkovic, D.M.; Ward, A.; Angove, H.C.; Day, P.J.; Vonrhein, C.; Tickle, I.J.; Jhoti, H. Crystal structures of human cytochrome P450 3A4 bound to metyrapone and progesterone. Science 2004, 305, 683–686. [Google Scholar] [CrossRef]
- Li, Y.C.; Chiang, J.Y.L. The expression of a catalytically active cholesterol 7α-hydroxylase cytochrome P450 in Escherichia coli. J. Biol. Chem. 1991, 266, 19186–19191. [Google Scholar] [CrossRef]
- Gillam, E.M.J.; Baba, T.; Kim, B.R.; Ohmori, S.; Guengerich, F.P. Expression of Modified Human Cytochrome P450 3A4 in Escherichia coli and Purification and Reconstitution of the Enzyme. Arch. Biochem. Biophys. 1993, 305, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Pernecky, S.J.; Larson, J.R.; Philpot, R.M.; Coon, M.J. Expression of truncated forms of liver microsomal P450 cytochromes 2B4 and 2E1 in Escherichia coli: Influence of NH2-terminal region on localization in cytosol and membranes. Proc. Natl. Acad. Sci. USA 1993, 90, 2651–2655. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Gillam, E.M.J.; Ohmori, S.; Tukey, R.H.; Peter Guengerich, F. Expression of modified human cytochrome P450 1A1 in Escherichia coli: Effects of 5’ substitution, stabilization, purification, spectral characterization, and catalytic properties. Arch. Biochem. Biophys. 1994, 312, 436–446. [Google Scholar] [CrossRef]
- Shimada, T.; Wunsch, R.M.; Hanna, I.H.; Sutter, T.R.; Guengerich, F.P.; Gillam, E.M.J. Recombinant human cytochrome P450 1B1 expression in Escherichia coli. Arch. Biochem. Biophys. 1998, 357, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Souček, P. Expression of cytochrome P450 2A6 in Escherichia coli: Purification, spectral and catalytic characterization, and preparation of polyclonal antibodies. Arch. Biochem. Biophys. 1999, 370, 190–200. [Google Scholar] [CrossRef]
- Hanna, I.H.; Reed, J.R.; Guengerich, F.P.; Hollenberg, P.F. Expression of human cytochrome P450 2B6 in Escherichia coli: Characterization of catalytic activity and expression levels in human liver. Arch. Biochem. Biophys. 2000, 376, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Bui, P.H.; Hankinson, O. Functional characterization of human cytochrome P450 2S1 using a synthetic gene-expressed protein in Escherichia coli. Mol. Pharmacol. 2009, 76, 1031–1043. [Google Scholar] [CrossRef]
- Larson, J.R.; Coon, M.J.; Porter, T.D. Alcohol-inducible cytochrome P-450IIE1 lacking the hydrophobic NH2-terminal segment retains catalytic activity and is membrane-bound when expressed in Escherichia coli. J. Biol. Chem. 1991, 266, 7321–7324. [Google Scholar] [CrossRef]
- Williams, P.A.; Cosme, J.; Ward, A.; Angove, H.C.; Jhoti, H. Crystal structure of human cytochrome P450 2C9 with bound warfarin. Nature 2003, 424, 464–468. [Google Scholar] [CrossRef]
- Yano, J.K.; Hsu, M.; Griffin, K.J.; Stout, C.D.; Johnson, E.F. with coumarin and methoxsalen. Nat. Struct. Mol. Biol. 2005, 12, 822–823. [Google Scholar] [CrossRef]
- Stark, K.; Dostalek, M.; Guengerich, F.P. Expression and purification of orphan cytochrome P450 4X1 and oxidation of anandamide. FEBS J. 2008, 275, 3706–3717. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Abd-Rashid, B.A.; Ismail, Z.; Ismail, R.; Mak, J.W.; Ong, C.E. Heterologous Expression of Human Cytochromes P450 2D6 and CYP3A4 in Escherichia coli and Their Functional Characterization. Protein J. 2011, 30, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Mak, J.W.; Ong, C.E. Heterologous expression of human cytochrome P450 (CYP) 2C19 in Escherichia coli and establishment of RP-HPLC method to serve as activity marker. Biomed. Chromatogr. 2013, 27, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Domanski, T.L.; Finta, C.; Halpert, J.R.; Zaphiropoulos, P.G. cDNA cloning and initial characterization of CYP3A43, a novel human cytochrome P450. Mol. Pharmacol. 2001, 59, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Stormo, G.D.; Schneider, T.D.; Gold, L.M. Nucleic Acids Research. Nucleic Acids Res. 1982, 10, 2971–2996. [Google Scholar] [CrossRef]
- Tuller, T.; Waldman, Y.Y.; Kupiec, M.; Ruppin, E. Translation ef fi ciency is determined by both codon bias and folding energy. Proc. Natl. Acad. Sci. USA 2010, 107, 3645–3650. [Google Scholar] [CrossRef]
- Sharp, P.M.; Cowe, E.; Higgins, D.G.; Shields, D.C.; Wolfe, K.H.; Wright, F. Codon usage patterns in Escherichia colli, Bacillus subtilis, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Drosophila melanogaster and Homo sapiens; a review of the considerable within-species diversity. Nucleic Acids Res. 1988, 16, 8207–8211. [Google Scholar] [CrossRef]
- Wu, Z.L.; Sohl, C.D.; Shimada, T.; Guengerich, F.P. Recombinant enzymes overexpressed in bacteria show broad catalytic specificity of human cytochrome P450 2W1 and limited activity of human cytochrome P450 2S1. Mol. Pharmacol. 2006, 69, 2007–2014. [Google Scholar] [CrossRef]
- Hoover, D.M.; Lubkowski, J. DNAWorks: An automated method for designing oligonucleotides for PCR-based gene synthesis. Nucleic Acids Res. 2002, 30, 1–7. [Google Scholar] [CrossRef]
- Loomanl, A.C.; Bodlaenderl, J.; Comstock, L.J.; Eaton, D.; Jhurani, P.; De Boer, H.A.; Knippenberg, P.H. Van Influence of the codon following the AUG initiation codon expression of a modified lacZ gene in Escherichia coli. EMBO J. 1987, 6, 2489–2492. [Google Scholar] [CrossRef]
- Nishihara, K.; Kanemori, M.; Yanagi, H.; Yura, T. Overexpression of Trigger Factor Prevents Aggregation of Recombinant Proteins in Escherichia coli. Appl. Environ. Microbiol. 2000, 66, 884–889. [Google Scholar] [CrossRef]
- Todd, M.J.; Viitanen, P.V.; Lorimer, G.H. Dynamics of the Chaperonin ATPase Cycle: Implications for Facilitated Protein Folding. Science 1994, 265, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, M.; Iwasaki, M. Improvement in the expression of CYP2B6 by co-expression with molecular chaperones GroES/EL in Escherichia coli. Protein Expr. Purif. 2006, 46, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Barnes, H.J. Maximizing Expression P450s of Eukaryotic. In Methods in Enzyymology; Academic Press: Cambridge, MA, USA, 1996; Volume 272, pp. 3–14. [Google Scholar]
- Fisher, C.W.; Caudle, D.L.; Martin-Wixtrom, C.; Quattrochi, L.C.; Tukey, R.H.; Waterman, M.R.; Estabrook, R.W. High-level expression of functional human cytochrome P450 1A2 in Escherichia coli. FASEB J. 1992, 6, 759–764. [Google Scholar] [CrossRef]
- Shet, M.S.; Fisher, C.W.; Holmans, P.L.; Estabrook, R.W. Human cytochrome P450 3A4: Enzymatic properties of a purified recombinant fusion protein containing NADPH-P450 reductase. Proc. Natl. Acad. Sci. USA 1993, 90, 11748–11752. [Google Scholar] [CrossRef]
- Gillam, E.M.J.; Guo, Z.; Peter Guengerich, F. Expression of modified human cytochrome P450 2E1 in Escherichia coli, purification, and spectral and catalytic properties. Arch. Biochem. Biophys. 1994, 312, 59–66. [Google Scholar] [CrossRef]
- Shet, M.S.; Fisher, C.W.; Arlotto, M.P.; Shackleton, C.H.L.; Holmans, P.L.; Martin-Wixtrom, C.A.; Saeki, Y.; Estabrook, R.W. Purification and enzymatic properties of a recombinant fusion protein expressed in Escherichia coli containing the domains of bovine P450 17A and rat NADPH-P450 reductase. Arch. Biochem. Biophys. 1994, 311, 402–417. [Google Scholar] [CrossRef] [PubMed]
- Gillam, E.M.J.; Guo, Z.Y.; Ueng, Y.F.; Yamazaki, H.; Cock, I.; Reilly, P.E.B.; Hooper, W.D.; Guengerich, F.P. Expression of cytochrome-p450-3a5 in escherichia coli: Effects of 5′ modification, purification, spectral characterization, reconstitution conditions, and catalytic activities. Arch. Biochem. Biophys. 1995, 317, 374–384. [Google Scholar] [CrossRef]
- Chun, Y.J.; Shimada, T.; Guengerich, F.P. Construction of a human cytochrome P450 1A1:Rat NADPH-cytochrome P450 reductase fusion protein cDNA and expression in Escherichia coli, purification, and catalytic properties of the enzyme in bacterial cells and after purification. Arch. Biochem. Biophys. 1996, 330, 48–58. [Google Scholar] [CrossRef]
- Yamakoshi, Y.; Kishimoto, T.; Sugimura, K.; Kawashima, H. Human prostate CYP3A5: Identification of a unique 5’-untranslated sequence and characterization of purified recombinant protein. Biochem. Biophys. Res. Commun. 1999, 260, 676–681. [Google Scholar] [CrossRef]
- Shimada, T.; Tsumura, F.; Gillam, E.M.J.; Guengerich, F.P.; Inoue, K. Roles of NADPH-P450 reductase in the O-deethylation of 7-ethoxycoumarin by recombinant human cytochrome P450 1B1 variants in Escherichia coli. Protein Expr. Purif. 2000, 20, 73–80. [Google Scholar] [CrossRef]
- Choi, S.; Han, S.; Lee, H.; Chun, Y.; Kim, D. Evaluation of Luminescent P450 Analysis for Directed Evolution of Human CYP4A11. Biomol. Ther. 2013, 21, 487–492. [Google Scholar] [CrossRef]
- Park, H.; Lim, Y.; Han, S.; Kim, D. Expression and Characterization of Truncated Recombinant Human Cytochrome P450 2J2. Toxicol. Res. 2014, 30, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Roellecke, K.; Jäger, V.D.; Gyurov, V.H.; Kowalski, J.P.; Mielke, S.; Rettie, A.E.; Hanenberg, H.; Wiek, C.; Girhard, M. Ligand characterization of CYP4B1 isoforms modified for high-level expression in Escherichia coli and HepG2 cells. Protein Eng. Des. Sel. 2017, 30, 207–218. [Google Scholar]
- Kempf, A.C.; Zanger, U.M.; Meyer, U.A. Truncated Human P450 2D6P: Expression in Escherichia coli, Ni2+-Chelate Affinity Purification, and Characterization of Solubility and Aggregation. Arch. Biochem. Biophys. 1995, 321, 277–288. [Google Scholar] [CrossRef]
- Richardson, T.H.; Jung, F.; Griffin, K.J.; Wester, M.; Raucy, J.L.; Kemper, B.; Bornheim, L.M.; Hassett, C.; Omiecinski, C.J.; Johnson, E.F. A universal approach to the expression of human and rabbit cytochrome P450s of the 2C subfamily in Escherichia coli. Arch. Biochem. Biophys. 1995, 323, 87–96. [Google Scholar] [CrossRef]
- Wu, Z.L.; Bartleson, C.J.; Ham, A.J.L.; Guengerich, F.P. Heterologous expression, purification, and properties of human cytochrome P450 27C1. Arch. Biochem. Biophys. 2006, 445, 138–146. [Google Scholar] [CrossRef]
- Appiah-Opong, R.; Commandeur, J.N.M.; Axson, C.; Vermeulen, N.P.E. Interactions between cytochromes P450, glutathione S-transferases and Ghanaian medicinal plants. Food Chem. Toxicol. 2008, 46, 3598–3603. [Google Scholar] [CrossRef]
- Milichovký, J.; Bárta, F.; Schmeiser, H.H.; Arlt, V.M.; Frei, E.; Stiborová, M.; Martínek, V. Active site mutations as a suitable tool contributing to explain a mechanism of aristolochic acid I nitroreduction by cytochromes P450 1A1, 1A2 and 1B1. Int. J. Mol. Sci. 2016, 17, 213. [Google Scholar] [CrossRef]
- Dong, J.; Porter, T.D. Coexpression of mammalian cytochrome P450 and reductase in Escherichia coli. Arch. Biochem. Biophys. 1996, 327, 254–259. [Google Scholar] [CrossRef]
- Karam, W.G.; Chiang, J.Y.L. Expression and purification of human cholesterol 7α-hydroxylase in Escherichia coli. J. Lipid Res. 1994, 35, 1222–1231. [Google Scholar] [CrossRef]
- Pikuleva, I.A.; Bjo, I.; Waterman, M.R. Expression, purification, and enzymatic properties of recombinant human cytochrome P450c27 (CYP27). Arch. Biochem. Biophys. 1997, 343, 123–130. [Google Scholar] [CrossRef]
- Parikh, A.; Guengerich, F.P. Expression, purification, and characterization of a catalytically active human cytochrome P450 1A2:Rat NADPH-cytochrome P450 reductase fusion protein. Protein Expr. Purif. 1997, 9, 346–354. [Google Scholar] [CrossRef]
- Grabovec, I.P.; Smolskaya, S.V.; Baranovsky, A.V.; Zhabinskii, V.N.; Dichenko, Y.V.; Shabunya, P.S.; Usanov, S.A.; Strushkevich, N.V. Ligand-binding properties and catalytic activity of the purified human 24-hydroxycholesterol 7α-hydroxylase, CYP39A1. J. Steroid Biochem. Mol. Biol. 2019, 193, 105416. [Google Scholar] [CrossRef]
- Elbing, K.; Llp, E.; Brent, R. Recipes and tools for culture of Escherichia coli. Curr. Protoc. Mol. Biol. 2019, 125, 1–19. [Google Scholar] [CrossRef]
- Miura, M.; Ito, K.; Hayashi, M.; Nakajima, M.; Tanaka, T. The Effect of 5-Aminolevulinic Acid on Cytochrome P450-Mediated Prodrug Activation. PLoS ONE 2015, 10, e0131793. [Google Scholar] [CrossRef]
- Yadav, R.; Scott, E.E. cro Endogenous insertion of non-native metalloporphyrins into human membrane cytochrome P450 enzymes. J. Biol. Chem. 2018, 293, 16623–16634. [Google Scholar] [CrossRef]
- Koop, D.R. Inhibition of Ethanol-Inducible Cytochrome P450I I E I by. Chem. Res. Toxicol. 1990, 3, 377–383. [Google Scholar] [CrossRef]
- San-miguel, T.; Pérez-bermúdez, P.; Gavidia, I. Production of soluble eukaryotic recombinant proteins in E. coli is favoured in early log-phase cultures induced at low temperature. Springerplus 2013, 2, 2–5. [Google Scholar] [CrossRef]
- Yim, S.K.; Ahn, T.; Jung, H.C.; Pan, J.G.; Yun, C.H. Temperature effect on the functional expression of human cytochromes P450 2A6 and 2E1 in Escherichia coli. Arch. Pharm. Res. 2005, 28, 433–437. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, L.; Chen, S.; Chang, M.; Chen, T. A modified osmotic shock for periplasmic release of a recombinant creatinase from Escherichia coli. Biochem. Eng. J. 2004, 19, 211–215. [Google Scholar] [CrossRef]
- Timasheff, S.N. By Weak Interactions with Water: How Do Solvents Affect These Processes? Annu. Rev. Biophys. Biomol. Struct. 1993, 22, 67–97. [Google Scholar] [CrossRef] [PubMed]
- Fjelstrup, S.; Andersen, M.B.; Thomsen, J.; Wang, J.; Stougaard, M.; Pedersen, F.S.; Ho, Y.; Hede, M.S.; Knudsen, B.R. The Effects of Dithiothreitol on DNA. Sensors 2017, 17, 1201. [Google Scholar] [CrossRef]
- Kaushik, J.K.; Bhat, R. Thermal Stability of Proteins in Aqueous Polyol Solutions: Role of the Surface Tension of Water in the Stabilizing Effect of Polyols. J. Phys. Chem. B 1998, 5647, 7058–7066. [Google Scholar] [CrossRef]
- Vagenende, V.; Yap, M.G.S.; Trout, B.L. Mechanisms of Protein Stabilization and Prevention of Protein Aggregation by Glycerol. Biochemistry 2009, 48, 11084–11096. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Aryasomayajula, A.; Selvaganapathy, P.R. A Review on Macroscale and Microscale Cell Lysis Methods. Micromachines 2017, 8, 83. [Google Scholar] [CrossRef]
- Acuña, J.M.B.; Hidalgo-dumont, C.; Pacheco, N.; Cabrera, A.; Poblete-castro, I. OPEN A novel programmable lysozyme- based lysis system in Pseudomonas putida for biopolymer production. Sci. Rep. 2017, 7, 1–11. [Google Scholar]
- Liu, Y.; Liu, X.; Cui, Y.; Yuan, W. Ultrasonics Sonochemistry Ultrasound for microalgal cell disruption and product extraction: A review. Ultrason. Sonochem. 2022, 87, 106054. [Google Scholar] [CrossRef]
- Serine, A. Natural Product Communications Purification and Biochemical Characterization of. Nat. Prod. Commun. 2010, 5, 931–934. [Google Scholar]
- Tsuchiya, T.; Saito, S. Use of N-octyl-β-D-thioglucoside, a new nonionic detergent, for solubilization and reconstitution of membrane proteins. J. Biochem. 1984, 96, 1593–1597. [Google Scholar] [CrossRef]
- Kobs, S.F. Recovery of Octyl & Glucoside Detergent / Protein Mixtures from. Anal. Biochem. 1990, 191, 47–49. [Google Scholar]
- Ahn, T.; Bae, C.S.; Yun, C.H. Affinity purification of recombinant human cytochrome P450s 3A4 and 1A2 using mixed micelle systems. Protein Expr. Purif. 2014, 101, 37–41. [Google Scholar] [CrossRef]
- Maslova, V.A.; Kiselev, M.A. Structure of Sodium Cholate Micelles. Crystallogr. Rep. 2018, 63, 472–475. [Google Scholar] [CrossRef]
- Subuddhi, U.; Mishra, A.K. Micellization of bile salts in aqueous medium: A fluorescence study. Colloids Surf. B Biointerfaces 2007, 57, 102–107. [Google Scholar] [CrossRef]
- Hernandez, O.; Kohli, K.K. Removal of the Nonionic Detergent Emulgen 911 from Solubilized Microsomes by HPLC. J. Liq. Chromatogr. 1983, 6, 1459–1473. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Martin, M.V. Purification of cytochromes P450. In Cytochrome P450 Protocols. Methods in Molecular Biology; Philips, I., Shephard, E., Eds.; Humana Press: Totowa, NJ, USA, 2006; Volume 320, pp. 31–37. ISBN 9781588294418. [Google Scholar]
- Dutton, D.R.; Mcmillen, S.K. Purification of Rat Liver Microsomal Cytochrome P-450b Without the Use of Nonionic Detergent. J. Biochem. Toxicol. 1988, 3, 131–145. [Google Scholar] [CrossRef]
- Hosea, N.A.; Guengerich, F.P. Oxidation of Nonionic Detergents by Cytochrome P450 Enzymes 1. Arch. Biochem. Biophys. 1998, 353, 365–373. [Google Scholar] [CrossRef]
- Ryan, D.E.; Thomas, P.E.; Levin, W. Purification and characterization of a minor form of hepatic microsomal cytochrome P-450 from rats treated with polychlorinated biphenyls. Arch. Biochem. Biophys. 1982, 216, 272–288. [Google Scholar] [CrossRef]
- Himmelhoch, R. Guide to Ion-Exchange Chromatography. Methods Enzymol. 1971, 22, 273–286. [Google Scholar]
- Jerker, P. Immobilized Metal Ion Affinity Chromatography. Protein Expr. Purif. 1992, 3, 263–281. [Google Scholar]
- Kim, D.; Guengerich, F.P. Selection of Human Cytochrome P450 1A2 Mutants with Enhanced Catalytic Activity for Heterocyclic Amine N-Hydroxylation. Biochemistry 2004, 43, 981–988. [Google Scholar] [CrossRef]
- Reed, J.R.; Kelley, R.W.; Backes, W.L. An evaluation of methods for the reconsitution of cytochromes P450 and NADPH P450 reductase into lipid vesicles. Drug Metab. Dispos. 2007, 34, 660–666. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Wang, P.; Mason, P.S. Immunological Comparison of Rat, Rabbit, and Human Liver NADPH-Cytochrome P-450 Reductases? Biochemistry 1981, 20, 2379–2385. [Google Scholar] [CrossRef]
- Pritchard, M.P.; Glancy, M.J.; Blake, J.A.; Gilham, D.E.; Burchell, B.; Wolf, C.R.; Friedberg, T. Functional co-expression of CYP2D6 and human NADPH-cytochrome P450 reductase in Escherichia coli. Pharmacogenetics 1998, 8, 33–42. [Google Scholar] [CrossRef]
- Quehl, P.; Hollender, J.; Schüürmann, J.; Brossette, T.; Maas, R.; Jose, J. Co-expression of active human cytochrome P450 1A2 and cytochrome P450 reductase on the cell surface of Escherichia coli. Microb. Cell Fact. 2016, 15, 1–15. [Google Scholar] [CrossRef]
- Park, H.G.; Lim, Y.R.; Han, S.; Jeong, D.; Kim, D. Enhanced purification of recombinant rat NADPH-P450 reductase by using a hexahistidine-tag. J. Microbiol. Biotechnol. 2017, 27, 983–989. [Google Scholar] [CrossRef]
- Brignac-huber, L.; Reed, J.R.; Backes, W.L. Organization of NADPH-Cytochrome P450 Reductase and CYP1A2 in the Endoplasmic Reticulum—Microdomain Localization Affects Monooxygenase Function. Mol. Pharmacol. 2011, 79, 549–557. [Google Scholar] [CrossRef]
- Peterson, J.A.; Keeffe, D.H.O. Temperature Dependent of Cytochrome P-450 Reduction. J. Biol. Chem. 1976, 251, 4010–4016. [Google Scholar] [CrossRef]
- Ingelman-sundberg, M.; Haaparanta, T.; Rydstrom, J. Membrane Charge as Effector of Cytochrome P-450LM2 Catalyzed Reactions in Reconstituted Liposomest. Biochemistry 1981, 20, 4100–4106. [Google Scholar] [CrossRef]
- Kim, K.; Ahn, T.; Yun, C. Membrane Properties Induced by Anionic Phospholipids and Phosphatidylethanolamine Are Critical for the Membrane Binding and Catalytic Activity of Human Cytochrome P450 3A4†. Biochemistry 2003, 42, 15377–15387. [Google Scholar] [CrossRef]
- Yi, E.; Yun, C.; Chae, H.; Chae, H.; Ahn, T. Anionic phospholipid-induced regulation of reactive oxygen species production by human cytochrome P450 2E1. FEBS Lett. 2008, 582, 1771–1776. [Google Scholar]
- Reed, J.R.; Brignac-huber, L.M.; Backes, W.L. Physical Incorporation of NADPH-cytochrome P450 Reductase and Cytochrome P450 into Phospholipid Vesicles using Glycocholate and Biobeads. Drug Metab. Dispos. 2009, 36, 582–588. [Google Scholar] [CrossRef]
- Biokatalyse, A.; Chemistry, P. Mechanism of rate control of the NADPH-dependent reduction of cytochrome P-450 by lipids in reconstituted phospholipid vesicles. Eur. J. Biochem. 1984, 114, 509–513. [Google Scholar]
- Ingelman-sundberg, M.; Hagbjörk, A.; Ueng, Y.; Yamazaki, H.; Guengerich, F.P. High Rates of Substrate Hydroxylation by Human Cytochrome P450 3A4 in Reconstituted Membranous Vesicles: Influence of Membrane Charge. Biochem. Biophys. Res. Commun. 1996, 221, 318–322. [Google Scholar] [CrossRef]
- Jang, H.; Kim, D.; Ahn, T.; Yun, C. Functional and conformational modulation of human cytochrome P450 1B1 by anionic phospholipids. Arch. Biochem. Biophys. 2010, 493, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Murtazina, D.A.; Andersson, U.; Hahn, I.S.; Bjorkhem, I.; Ansari, G.A.S.; Pikuleva, I.A. Phospholipids modify substrate binding and enzyme activity of human cytochrome P450 27A1. J. Lipid Res. 2004, 45, 2345–2353. [Google Scholar] [CrossRef]
- Mclaughlin, L.A.; Ronseaux, S.; Finn, R.D.; Henderson, C.J.; Wolf, C.R. Deletion of Microsomal Cytochrome b 5 Profoundly Affects Hepatic and Extrahepatic Drug Metabolism. Mol. Pharmacol. 2010, 78, 269–278. [Google Scholar] [CrossRef]
- Porter, T.D. The Roles of Cytochrome b 5 in Cytochrome P450 Reactions. J. Biochem. Mol. Toxicol. 2002, 16, 311–316. [Google Scholar] [CrossRef]
- Oxygenase, H.; Yamazaki, H.; Shimada, T.; Martin, M.V.; Guengerich, F.P. Stimulation of Cytochrome P450 Reactions by Apo-cytochrome b 5. J. Biol. 2001, 276, 30885–30891. [Google Scholar]
- Akhtar, M.K.; Kelly, S.L.; Kaderbhai, M.A. Cytochrome b 5 modulation of 17 hydroxylase and 17–20 lyase ( CYP17 ) activities in steroidogenesis. J. Endocrinol. 2005, 187, 267–274. [Google Scholar] [CrossRef]

| CYP | Suspension Buffer | Lysis Approach (yes/no) | Protease Inhibitor | Ultra Centrifugation | Reference | ||
|---|---|---|---|---|---|---|---|
| High-Pressure Homogenizer | Lysozyme | Ultrasonic Cavitation | |||||
| 2E1 | Potassium phosphate buffer | Yes | No | No | N/A | 142,000 g for 1 h | [7] |
| 17A1 | Mops ** buffer | No | Yes | Yes | PMSF, leupetin, aprotinin | 225,000 g for 30 min | [25] |
| 1A2 | Potassium phosphate buffer | Yes | No | No | N/A | 100,000 g for 60 min | [66] |
| 3A4 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 180,000 g for 65 min | [42] |
| 1A1 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 180,000 g for 65 min | [44] |
| 2E1 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 180,000 g for 65 min | [68] |
| 1A2 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin | 180,000 g for 65 min | [38] |
| 7A1 | Potassium phosphate buffer | No | Yes | No | PMSF | 100,000 g for 60 min | [83] |
| 17A1-OxR | Tris-HCl buffer | No | Yes | Yes | PMSF | 100,000 g for 60 min | [69] |
| 2D6 | Potassium phosphate buffer | Yes | No | No | PMSF, leupeptin | 142,000 g for 60 min | [77] |
| 2D6 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 100,000 g for 45 min, supernatants further centrifuge at 100,000 g for 16 h | [39] |
| 2E1-OxR | Potassium phosphate buffer | Yes | No | No | N/A | 142,000 g for 1 h | [82] |
| 1A1-OxR | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 180,000 g for 65 min | [71] |
| 27A1 | Potassium phosphate buffer | No | Yes | Yes | PMSF | 146,000 g for 60 min | [84] |
| 3A5 | Tris acetate buffer | No | Yes | No | PMSF, aprotinin | 193,000 g for 40 min | [72] |
| 2A6 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 180,000 g for 65 min | [46] |
| 2B6 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin | 180,000 g for 65 min | [47] |
| 2D6-OxR | Tris acetate buffer | No | Yes | Yes | PMSF, protease inhibitor cocktail | 100,000 g | [27] |
| 1A2-HDJ-1 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin | 180,000 g for 65 min | [26] |
| 2B6-GroES/EL | Tris-HCl buffer | No | Yes | Yes | N/A | 100,000 g for 60 min | [64] |
| 27C1 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 180,000 g for 65 min | [79] |
| 4X1 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin, aprotinin, bestatin | 180,000 g for 65 min | [52] |
| 2S1 | Potassium phosphate buffer | Yes | No | No | PMSF | N/A | [48] |
| 1A1-OxR | Tris acetate buffer | No | Yes | Yes | N/A | 100,000 g for 75 min | [8] |
| 2C10 &2C9 | Tris acetate buffer | No | Yes | Yes | PMSF, leupeptin | 180,000 g for 65 min | [24] |
| 2J2 | Tris acetate buffer | No | Yes | No | PMSF, protease inhibitor cocktail | 100,000 g for 3 h | [75] |
| 4B1 | Potassium phosphate buffer | Yes | No | No | PMSF, protease inhibitor cocktail | N/A | [76] |
| CYP | Detergent | Column (s) | OxR | Cytochrome b5 | Specific Content (nmol/mg Protein) | Reference |
|---|---|---|---|---|---|---|
| 2E1 | n-octylglucoside | S-Sepharose Hydroxyapatite DEAE-Sepharose Hydroxyapatite | N/A | N/A | 15.8 | [7] |
| 2E1 | n-octylglucoside | S-Sepharose | Rabbit liver | N/A | 2 | [49] |
| 3A4 | Sodium cholate | Octylamino-Sepharose Cosmogel DEAE KB Type-S Cosmogel CM Hydroxylapatite | Rabbit liver | Human liver | 23 | [42] |
| 2E1&2B4 | Tergitol NP-10 | S-Sepharose (CYP2E1) high-resolution hydroxyapatite (CYP2B4) | N/A | N/A | N/A | [43] |
| 3A4+OxR | Emulgen 911 | 2′,5′-ADP Sepharose affinity | Fused OxR | N/A | 150–200 per L of culture | [67] |
| 1A1 | Sodium cholate Triton N-101 | DEAE-Sephacel CM-Sepharose fast-flow Hydroxylapatite | Rabbit liver | Human liver | 10–15 per L of culture | [44] |
| 2E1 | Sodium cholate Triton N-101 | DEAE-Sephacel CM-Sepharose fast-flow Hydroxylapatite | Rabbit liver | Human liver | 160 per L of culture | [68] |
| 1A2 | Sodium cholate Triton N-101 | DEAE-Sephacel CM-Sepharose fast-flow | Rabbit liver | Human liver | 225–245 per L of culture | [38] |
| 17A1-OxR | Emulgen 911 | DE-52 2′,5′-ADP-Sepharose 4B LKB Ultragel AcA34 | Co-expressed | N/A | 3.8 | [69] |
| 3A5 | Sodium cholate Triton N-101 | DEAE-Sephacel CM-Sepharose fast-flow Hydroxylapatite | Rabbit liver | Human liver | 260 per L of culture | [70] |
| 2D6 | C12E9 | Ni2+-NTA-agarose DEAE-Sephacel HTP hydroxylapatite | Rat liver | N/A | 20–40 per L of culture | [77] |
| 2D6 | Triton X-114 | E. coli flavodoxin affinity Biogel HTP hydroxylapatite | Rabbit liver | Human liver | 90 per L of culture | [39] |
| 2E1-OxR | n-octylglucoside | S-Sepharose Hydroxyapatite DEAE-Sepharose Hydroxyapatite | Co-expressed | Rabbit liver | 0.11 | [82] |
| 1A1-OxR | Emulgen 911 | DE-52 2′,5′-ADP agarose BioGel HTP hydroxylapatite | Co-expressed | Human liver | 25 per L of culture | [71] |
| 27A1 | Emulgen 913 | DEAE-cellulose Hydroxylapatite | Rat liver | N/A | 3.5 | [84] |
| 1A2-OxR | Sodium cholate | octyl-Sepharose Hydroxylapatite Adrenodoxin-Sepharose | Co-expressed | N/A | 15 | [85] |
| 1B1-OxR | Emulgen 911 | DE-52 2′,5′-ADP agarose BioGel HTP hydroxylapatite | Co-expressed | Human liver | 35 per L of culture | [45] |
| 3A5 | Sodium cholate Triton N-101 | DEAE-Sephacel CM-Sepharose (fast-flow) Hydroxylapatite | Recombinant | Recombinant | 9.2 | [72] |
| 2A6 | Renex-690 | Ni2+-NTA agarose Bio-Gel HTP hydroxylapatite | Rabbit liver | N/A | 12.35 | [46] |
| 2B6 | Sodium cholate Triton N-101 | DEAE-Sephacel Amberlite XAD-2 CM-Sepharose Hydroxylapatite | E. coli expressed rat OxR | N/A | 25–80 per L of culture | [47] |
| 2D6-OxR | Sodium cholate Triton N-101 | DEAE-Sephacel Hydroxylapatite | Co-expressed | N/A | 1–3 | [27] |
| 1A2-HDJ-1 | Emulgen 911 | Mono Q Hi-Trap 2′-5′-ADP-Sepharose Sephacryl S-200 HR 2′-5′-ADP-Sepharose | Expressed from E. coli | N/A | 60–120 per L of culture | [26] |
| 2B6-GroES/EL | Sodium cholate Triton N-101 | TOYOPEARL DEAE-650M TOYOPEARL SP-550C Hydroxyapapite | Commercial purchased | N/A | 8.2 | [64] |
| 4X1 | Sodium CHAPS | Ni2+-nitriloacetic acid | Co-expressed | N/A | 100–200 per L of culture | [52] |
| 2S1 | CHAPS | NTA agarose | Co-expressed | N/A | 16 | [48] |
| 1A1-OxR | Emulgen 911 Sodium cholate | Ni-NTA agarose | Co-expressed | N/A | N/A | [8] |
| 2C10 &2C9 | Emulgen 911 Sodium cholate | DEAE-Sephacel Hydroxylapatite | Rabbit liver | Human liver | 5–19 per L of culture | [24] |
| 4A11 | CHAPS | Ni-NTA agarose | Co-expressed | N/A | 125–320 per L of culture | [74] |
| 2J2 | CHAPS | Ni-NTA agarose | Rat liver | N/A | 16–18.6 per L of culture | [75] |
| 39A1-GroEL/ES | CHAPS TritonX100 | Ni-NTA-protino Ion-exchange Source S | Recombinant | N/A | N/A | [86] |
| 2J2-GroEL/ES | CHAPS | Ni2+-NTA agarose | Rat liver | N/A | 140–230 | [9] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shang, T.; Fang, C.M.; Ong, C.E.; Pan, Y. Heterologous Expression of Recombinant Human Cytochrome P450 (CYP) in Escherichia coli: N-Terminal Modification, Expression, Isolation, Purification, and Reconstitution. BioTech 2023, 12, 17. https://doi.org/10.3390/biotech12010017
Shang T, Fang CM, Ong CE, Pan Y. Heterologous Expression of Recombinant Human Cytochrome P450 (CYP) in Escherichia coli: N-Terminal Modification, Expression, Isolation, Purification, and Reconstitution. BioTech. 2023; 12(1):17. https://doi.org/10.3390/biotech12010017
Chicago/Turabian StyleShang, Tao, Chee Mun Fang, Chin Eng Ong, and Yan Pan. 2023. "Heterologous Expression of Recombinant Human Cytochrome P450 (CYP) in Escherichia coli: N-Terminal Modification, Expression, Isolation, Purification, and Reconstitution" BioTech 12, no. 1: 17. https://doi.org/10.3390/biotech12010017