
Vocal Cues to Assess Arousal State of Bottlenose Dolphins (Tursiops spp.) Involved in Public Presentations

Abstract
:1. Introduction
2. Materials and Methods
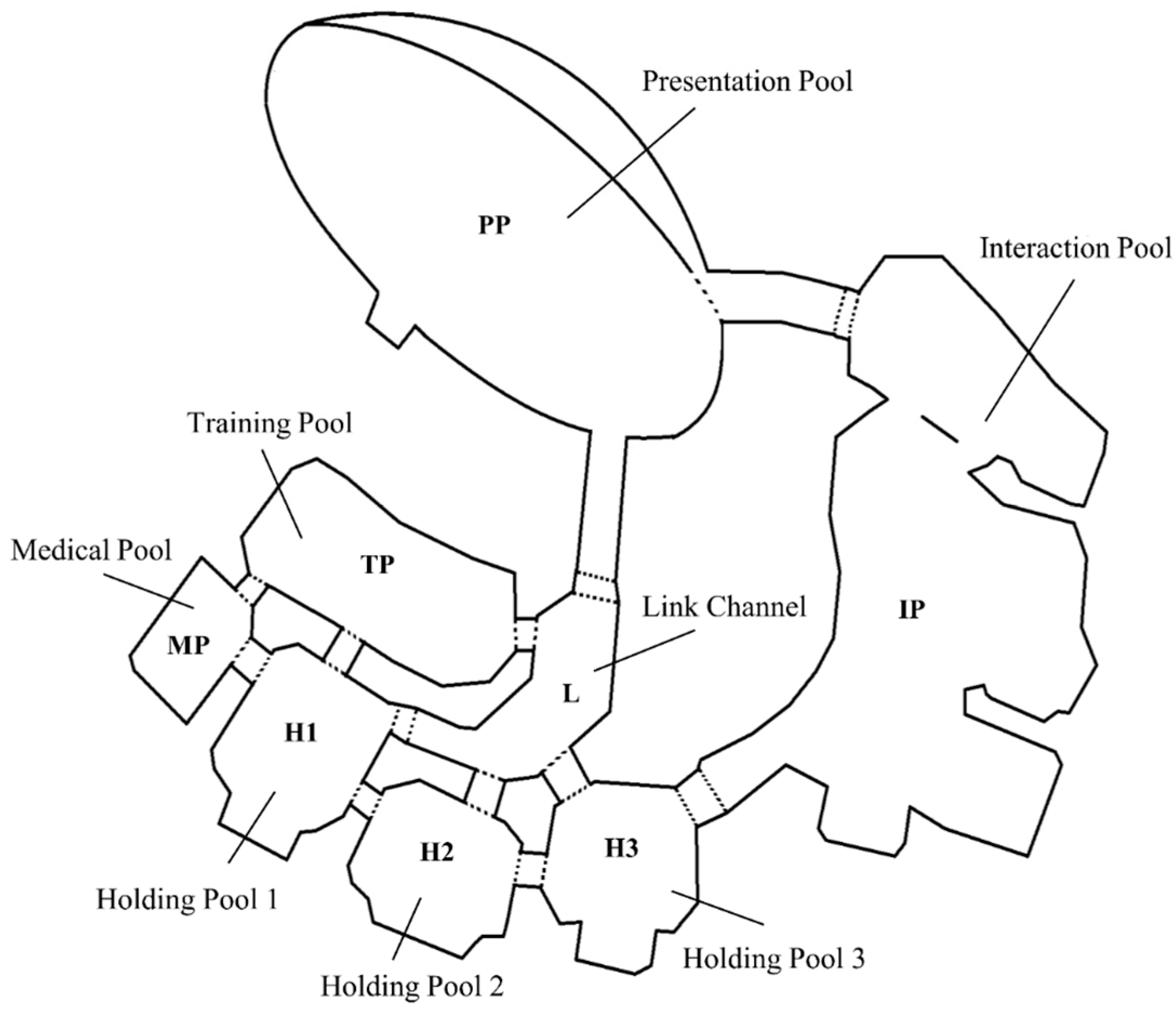
2.1. Data Collection and Study Animals
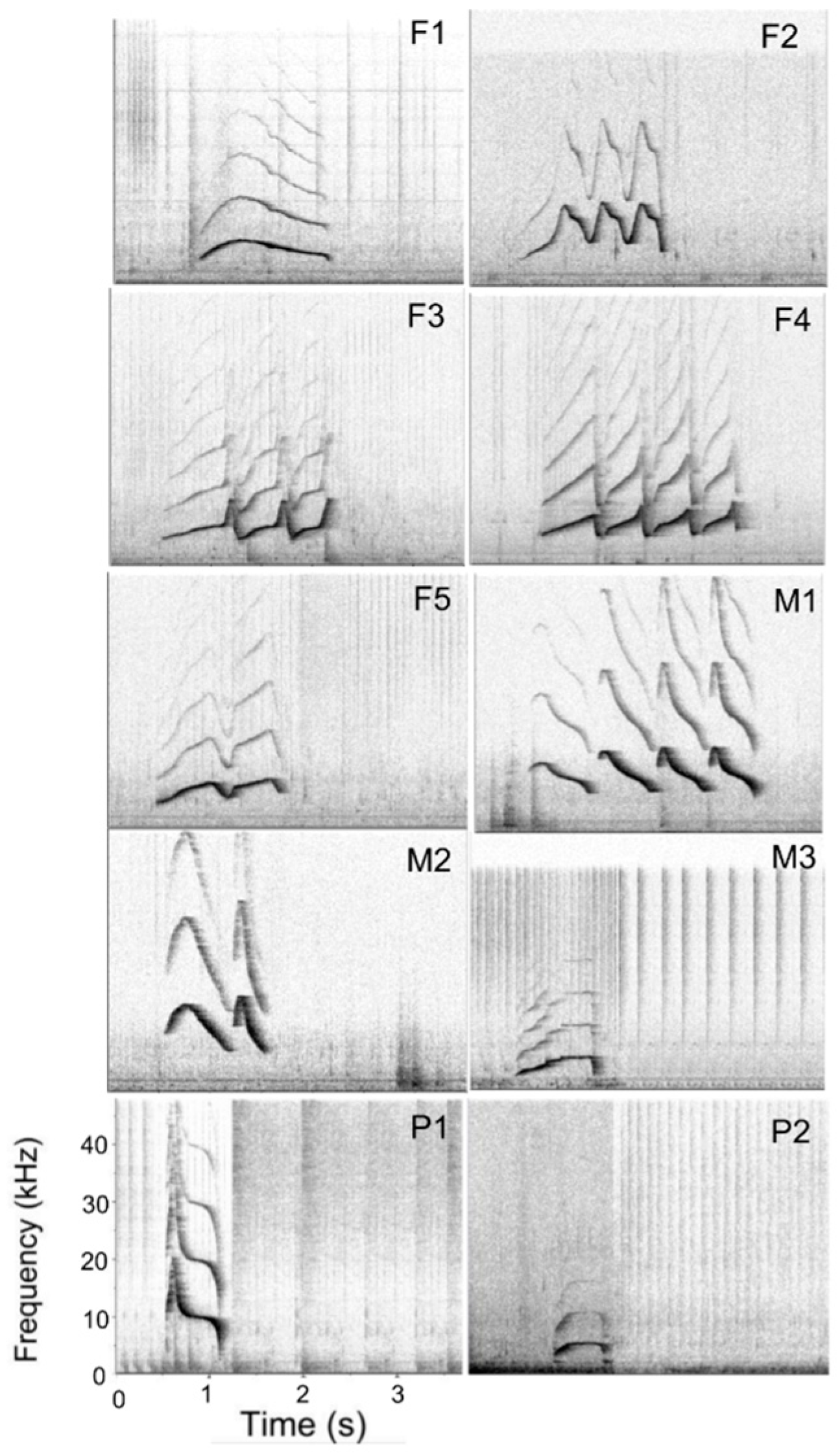
2.2. Signature Whistle Catalogue
2.3. Behavioural Contexts
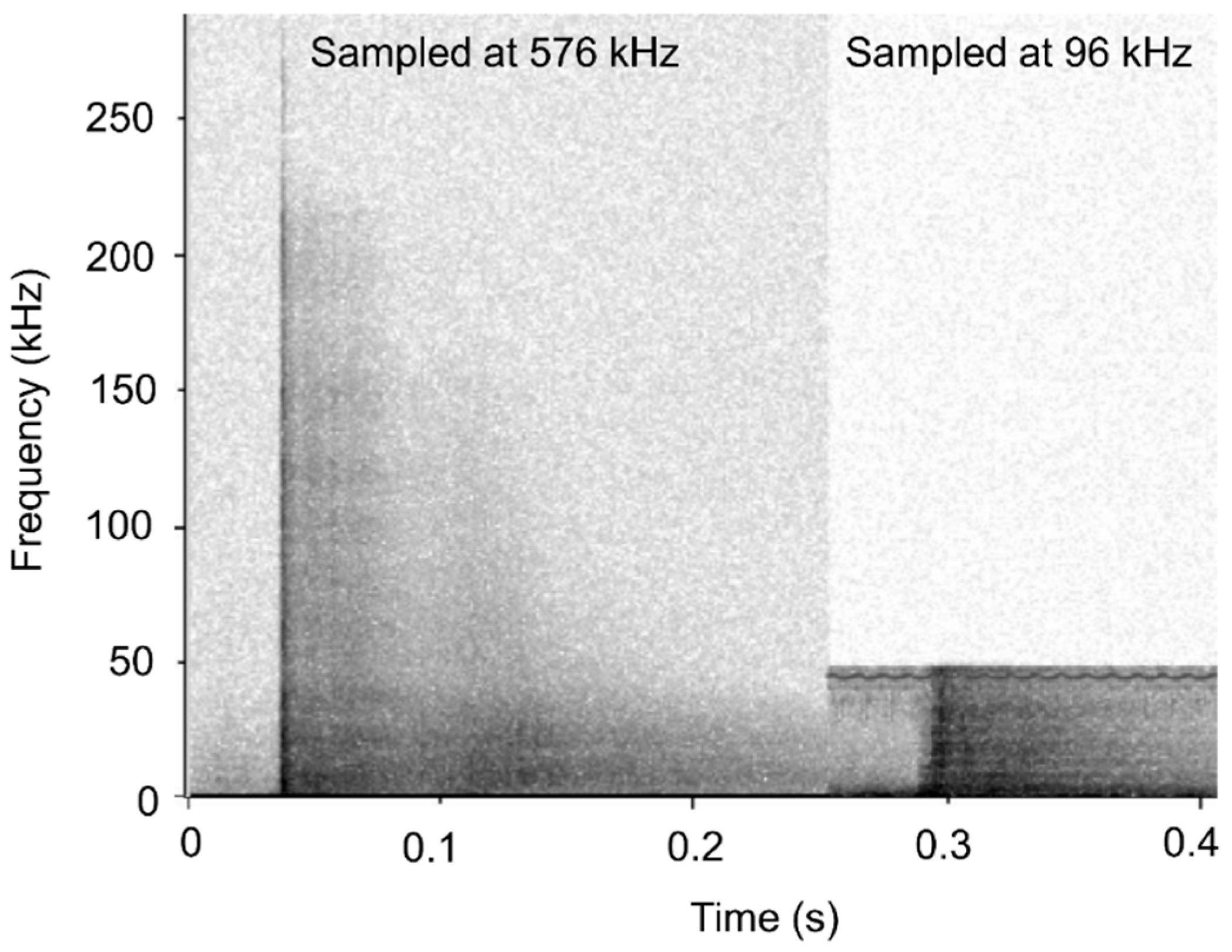
2.4. Identification of Sound Types
2.5. Influence of Context on Group and Individual Vocal Production Rates
2.6. Shifts in Individual Signature Whistle Characteristics between Low and High Arousal Contexts
3. Results
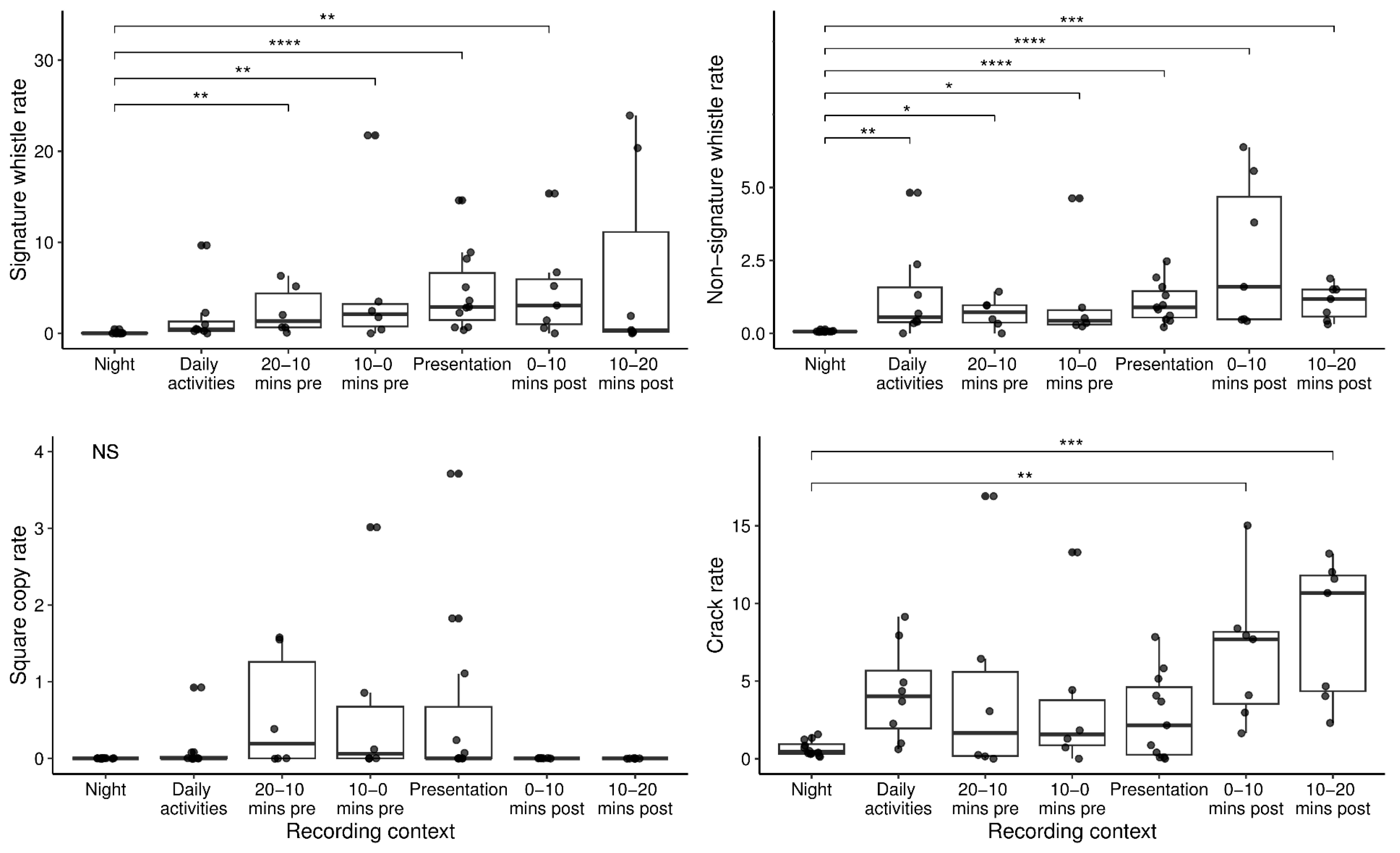
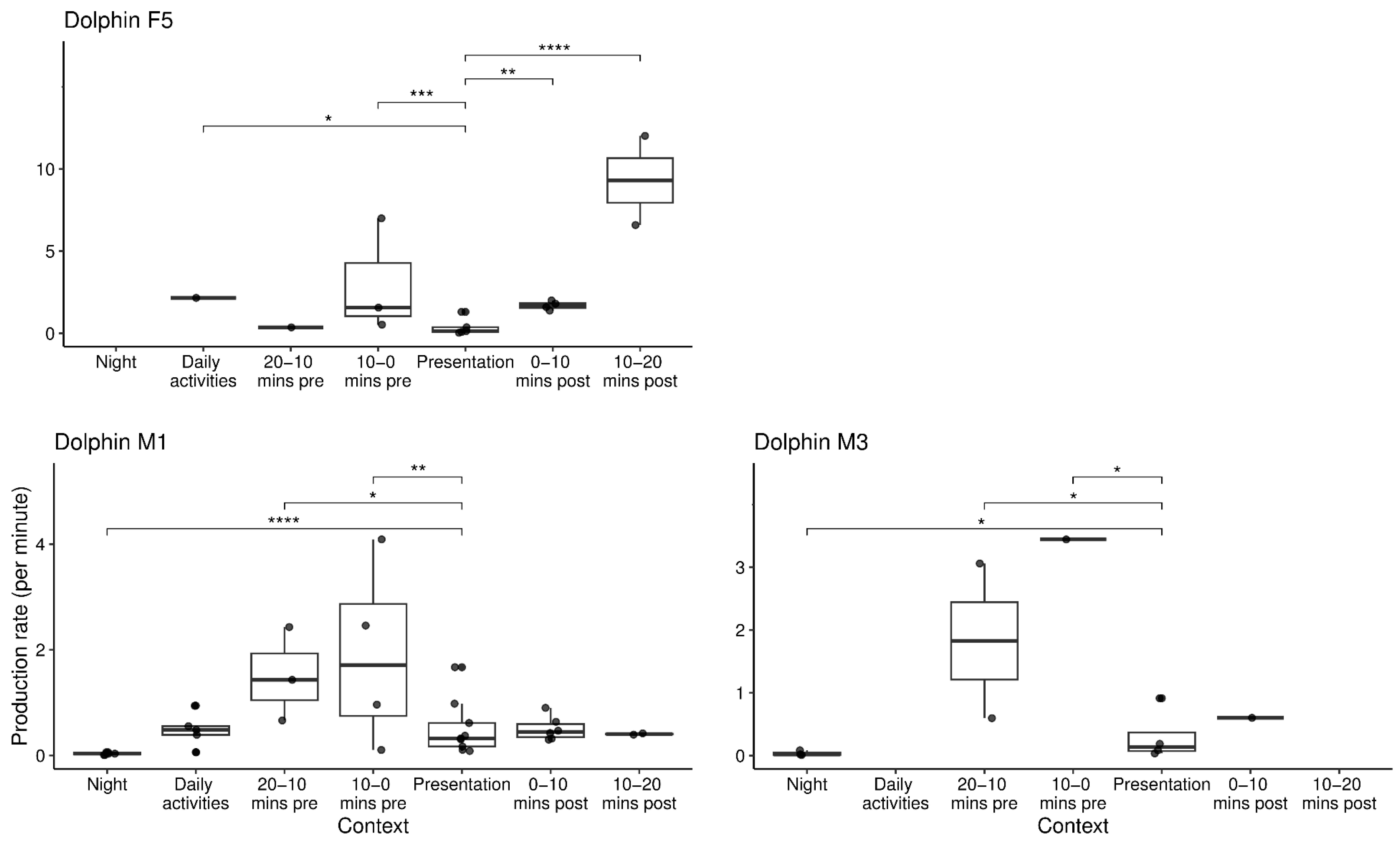
3.1. Influence of Context on Group and Individual Vocal Production Rates
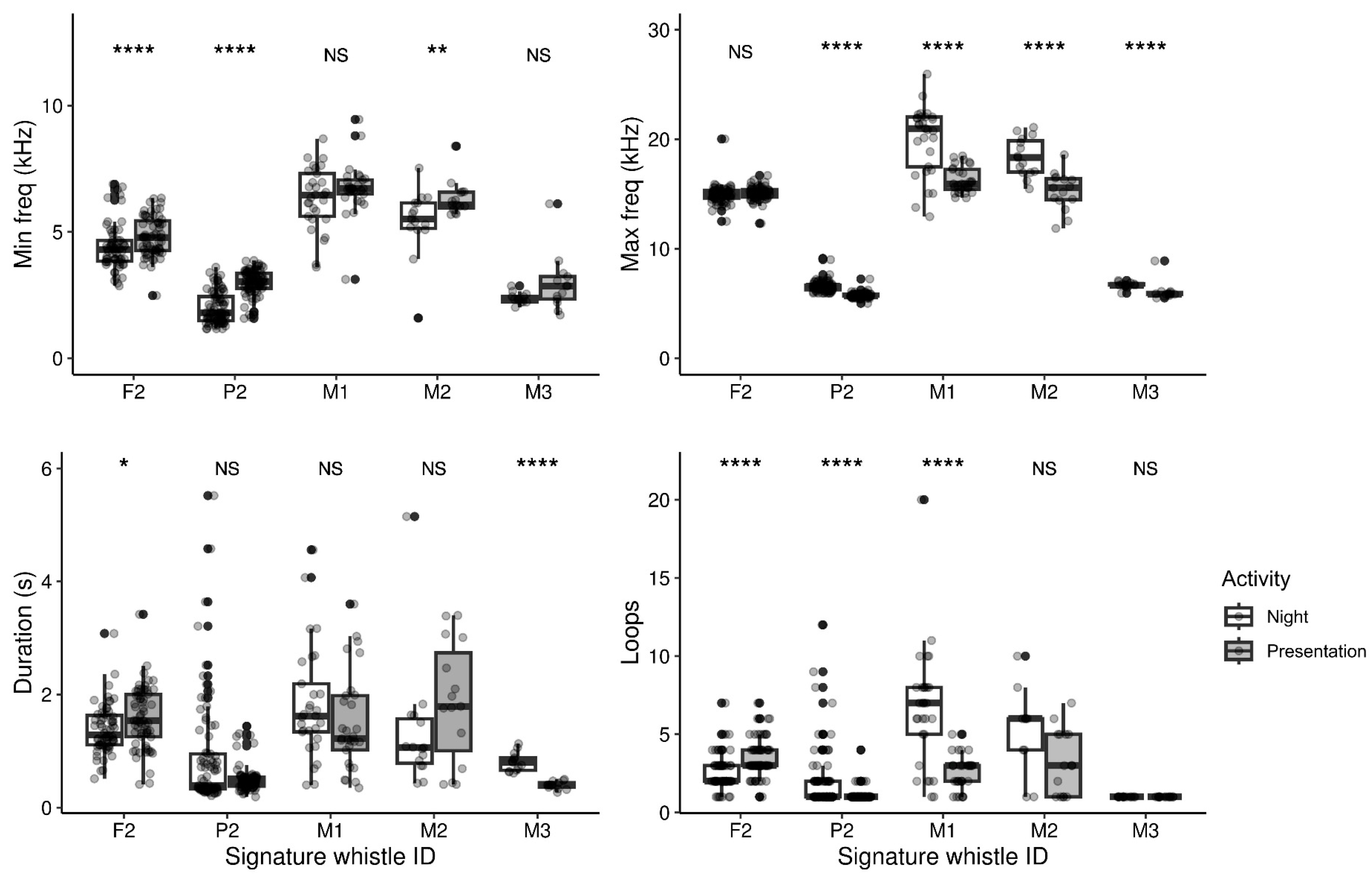
3.2. Shifts in Individual Signature Whistle Characteristics between Low and High Arousal Contexts
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Social Group | Name | Species | Sex | In Human Care Since | Date of Birth | Age in 2018 | Signature Whistle ID |
|---|---|---|---|---|---|---|---|
| Female group | Affrika | Tt | F | Born in facility | 12/05/1995 | 23 | F1 |
| Zulu | Ta-Tt Hybrid | F | Born in facility | 09/12/1998 | 19 | F2 | |
| Khanya | Ta-Tt F2 | F | Born in facility | 17/07/1993 | 25 | F3 | |
| Tombi | Ta-Tt Hybrid | F | Born in facility | 23/05/1993 | 25 | F4 | |
| Khethiwe b | Ta-Tt Hybrid | F | Born in facility | 25/11/2008 | 9 | F5 | |
| Male group | Ingelosi | Ta-Tt Hybrid | M | Born in facility | 22/05/2004 | 14 | M1/M2 |
| Khwezi | Ta-Tt Hybrid | M | Born in facility | 07/09/1995 | 24 | M1/M2 | |
| Kelpie | Tt | M | Born in facility | 22/01/1984 | 34 | M3 | |
| Jula | Ta-Tt Hybrid | M | Born in facility | 28/07/1990 | Deceased 2015 | M4 | |
| Mixed group | Gambit | Tt | M | 08/12/1976 | 08/12/1971 a | 46 | P1 |
| Frodo | Ta | F | 26/06/1979 | 26/06/1974 a | 44 | P2 |
| Valence | Context | Setting | Whistle Production | Frequency Parameters | Duration | Loops | Reference |
|---|---|---|---|---|---|---|---|
| Unsure | Tour boat presence | Wild | NA | Increase Min F Increase Max F | No shifts | NA | [46] |
| Unsure | Tour boat presence | Wild | Increase | No shifts | No shifts | NA | [97] |
| Unsure | Birth | Human care | Increase | NA | NA | NA | [98] |
| Unsure | Public presentations | Human care | Increase | Decrease Max F | Increase | Varied between individuals | This study |
| Likely positive | Human in-water interactions | Human care | Increase | NA | NA | NA | [99] |
| Positive | Feeding, staff presence | Human care | Increase | Decrease Max F | Decrease | Decrease | [25] |
| Positive | ‘Swim with dolphin’ tours | Wild | Increase | NA | NA | NA | [83] |
| Positive | Social, feeding | Wild | Increase | NA | NA | NA | [100] |
| Positive | Social, dispersed | Wild | Increase | NA | NA | NA | [101] |
| Positive | Social, feeding | Wild | Increase | NA | NA | NA | [102] |
| Positive | Social | Wild | Increase | NA | NA | NA | [34] |
| Positive | Feeding, social | Wild | NA | Increase Max F (feeding) Decrease Max F (social) | No shifts | NA | [103] |
| Positive | Feeding, social | Wild | NA | Increase Max F (feeding) Decrease Max F (social) | Increase (feeding) Decrease (social) | NA | [104] |
| Positive | Social, foraging | Wild | Increase | NA | NA | NA | [105] |
| Positive + negative | Sexual + aggressive combined | Wild | Increase | Decrease Min F Decrease F range | No shifts | NA | [85] |
| Negative | Capture-release | Wild | Increase | Increase Max F | Increase | Increase | [24] |
| Negative | Lowering water levels in tanks | Human care | Decrease | NA | NA | NA | [43] |
References
- Waples, K.A.; Gales, N.J. Evaluating and minimising social stress in the care of captive bottlenose dolphins (Tursiops aduncus). Zoo Biol. 2002, 21, 5–26. [Google Scholar] [CrossRef]
- Mason, G.J. Species differences in responses to captivity: Stress, welfare and the comparative method. Trends Ecol. Evol. 2010, 25, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.K.; Borger-Turner, J.L.; Eskelinen, H.C. C-Well: The development of a welfare assessment index for captive bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2015, 24, 267–282. [Google Scholar] [CrossRef]
- de Waal, F.B.M. What is an animal emotion? Ann. N. Y. Acad. Sci. 2011, 1224, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Burman, O.H.; Paul, E.S. An integrative and functional framework for the study of animal emotion and mood. Proc. R. Soc. B Biol. Sci. 2010, 277, 2895–2904. [Google Scholar] [CrossRef] [PubMed]
- Briefer, E.F. Vocal expression of emotions in mammals: Mechanisms of production and evidence. J. Zool. 2012, 288, 1–20. [Google Scholar] [CrossRef]
- Briefer, E.F.; Tettamanti, F.; McElligott, A. Emotions in goats: Mapping physiological, behavioural and vocal 2 profiles. Anim. Behav. 2015, 99, 131–143. [Google Scholar] [CrossRef]
- Baciadonna, L.; Briefer, E.F.; Favaro, L.; McElligott, A. Goats distinguish between positive and negative emotion-linked vocalisations. Front. Zool. 2019, 16, 25. [Google Scholar] [CrossRef]
- Maigrot, A.L. Vocal Expression of Emotions in Przewalski’s Horses and Wild Boars: Encoding and Decoding. Doctoral Dissertation, University of Bern, Bern, Switzerland, 2018. [Google Scholar]
- Carder, G.; Semple, S. Visitor effects on anxiety in two captive groups of western lowland gorillas. Appl. Anim. Behav. Sci. 2008, 115, 211–220. [Google Scholar] [CrossRef]
- Yon, L.; Williams, E.; Harvey, N.D.; Asher, L. Development of a behavioural welfare assessment tool for routine use with captive elephants. PLoS ONE 2019, 14, e0210783. [Google Scholar] [CrossRef]
- Linhart, P.; Ratcliffe, V.F.; Reby, D.; Špinka, M. Expression of emotional arousal in two different piglet call types. PLoS ONE 2015, 10, e0135414. [Google Scholar] [CrossRef]
- Luo, L.; Reimert, I.; de Haas, E.N.; Kemp, B.; Bolhuis, J.E. Effects of early and later life environmental enrichment and personality on attention bias in pigs (Sus scrofa domesticus). Anim. Cogn. 2019, 22, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Melotti, L.; Oostindjer, M.; Bolhuis, J.E.; Held, S.; Mendl, M. Coping personality type and environmental enrichment affect aggression at weaning in pigs. Appl. Anim. Behav. Sci. 2011, 133, 144–153. [Google Scholar] [CrossRef]
- Kuczaj, S.A., II; Highfill, L.; Byerly, H. The importance of considering context in the assessment of personality characteristics: Evidence from ratings of dolphin personality. Int. J. Comp. Psychol. 2012, 25, 309–329. [Google Scholar] [CrossRef]
- Highfill, L.E.; Kuczaj, S.A. Do bottlenose dolphins (Tursiops truncatus) have distinct and stable personalities? Aquat. Mamm. 2007, 33, 380. [Google Scholar] [CrossRef]
- Waitt, C.; Buchanan-Smith, H.M. What time is feeding? How delays and anticipation of feeding schedules affect stump-tailed macaque behavior. Appl. Anim. Behav. Sci. 2001, 75, 75–85. [Google Scholar] [CrossRef]
- Bassett, L.; Buchanan-Smith, H.M. Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 2007, 102, 223–245. [Google Scholar] [CrossRef]
- Ulyan, M.J.; Burrows, A.E.; Buzzell, C.A.; Raghanti, M.A.; Marcinkiewicz, J.; Phillips, K.A. The effects of predictable and unpredictable feeding schedules on the behavior and physiology of captive brown capuchins (Cebus apella). Appl. Anim. Behav. Sci. 2006, 101, 154–160. [Google Scholar] [CrossRef]
- Gottlieb, D.H.; Coleman, K.; McCowan, B. The effects of predictability in daily husbandry routines on captive rhesus macaques (Macaca mulatta). Appl. Anim. Behav. Sci. 2013, 143, 117–127. [Google Scholar] [CrossRef]
- de Oliveira, L.B.S.; da Silva Vasconcellos, A. May unpredictable events affect monkey welfare under human care? Behav. Process. 2022, 200, 104665. [Google Scholar] [CrossRef]
- Miller, L.J.; Lauderdale, L.K.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Assessment of animal management and habitat characteristics associated with social behavior in bottlenose dolphins across zoological facilities. PLoS ONE 2021, 16, e0253732. [Google Scholar] [CrossRef]
- Miller, L.J.; Lauderdale, L.K.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Relationships between animal management and habitat characteristics with two potential indicators of welfare for bottlenose dolphins under professional care. PLoS ONE 2021, 16, e0252861. [Google Scholar] [CrossRef] [PubMed]
- Esch, H.C.; Sayigh, L.S.; Blum, J.E.; Wells, R.S. Whistles as potential indicators of stress in bottlenose dolphins (Tursiops truncatus). J. Mammal. 2009, 90, 638–650. [Google Scholar] [CrossRef]
- Probert, R.; Bastian, A.; Elwen, S.H.; James, B.S.; Gridley, T. Vocal correlates of arousal in bottlenose dolphins (Tursiops spp.) in human care. PLoS ONE 2021, 16, e0250913. [Google Scholar] [CrossRef]
- Rendall, D. Acoustic correlates of caller identity and affect intensity in the vowel-like grunt vocalizations of baboons. J. Acoust. Soc. Am. 2003, 113, 3390–3402. [Google Scholar] [CrossRef] [PubMed]
- Trut, L.; Kharlamova, A.; Gogoleva, S.; Volodina, E.; Volodin, I. Sign and strength of emotional arousal: Vocal correlates of positive and negative attitudes to humans in silver foxes (Vulpes vulpes). Behaviour 2010, 147, 1713–1736. [Google Scholar] [CrossRef]
- Gouzoules, H.; Gouzoules, S. Agonistic screams differ among four species of macaques: The significance of motivation-structural rules. Anim. Behav. 2000, 59, 501–512. [Google Scholar] [CrossRef]
- Robbins, R.; McCreery, E.K. African wild dog pup vocalizations with special reference to Morton’s model. Behaviour 2003, 140, 333–351. [Google Scholar] [CrossRef]
- Feighny, J.A.; Williamson, K.E.; Clarke, J.A. North American elk bugle vocalizations: Male and female bugle call structure and context. J. Mammal. 2006, 87, 1072–1077. [Google Scholar] [CrossRef]
- Taylor, A.M.; Reby, D.; McComb, K. Context-related variation in the vocal growling behaviour of the domestic dog (Canis familiaris). Ethology 2009, 115, 905–915. [Google Scholar] [CrossRef]
- Cranford, T.W. In search of impulse sound sources in odontocetes. In Hearing by Whales and Dolphins; Fay, R.R., Popper, A.N., Eds.; Springer: New York, NY, USA, 2000; pp. 109–155. [Google Scholar]
- Sayigh, L.S.; Tyack, P.L.; Wells, R.S.; Scott, M.D. Signature whistles of free-ranging bottlenose dolphins Tursiops truncatus: Stability and mother-offspring comparisons. Behav. Ecol. Sociobiol. 1990, 26, 247–260. [Google Scholar] [CrossRef]
- Cook, M.L.H.; Sayigh, L.S.; Blum, J.E.; Wells, R.S. Signature–whistle production in undisturbed free–ranging bottlenose dolphins (Tursiops truncatus). Proc. R. Soc. B Biol. Sci. 2004, 271, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Janik, V.M.; King, S.L.; Sayigh, L.S.; Wells, R.S. Identifying signature whistles from recordings of groups of unrestrained bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 2013, 29, 109–122. [Google Scholar] [CrossRef]
- Therrien, S.C.; Thomas, J.A.; Therrien, R.E.; Stacey, R. Time of Day and Social Change Affect Underwater Sound Production by Bottlenose Dolphins (Tursiops truncatus) at the Brookfield Zoo. Aquat. Mamm. 2012, 38, 65–75. [Google Scholar] [CrossRef]
- Herzing, D.L. Clicks, whistles and pulses: Passive and active signal use in dolphin communication. Acta. Astronaut. 2014, 105, 534–537. [Google Scholar] [CrossRef]
- Au, W.W. (Ed.) Characteristics of dolphin sonar signals. In The Sonar of Dolphins; Springer: New York, NY, USA, 1993; pp. 115–139. [Google Scholar]
- Janik, V.M.; Slater, P.J. Context-specific use suggests that bottlenose dolphin signature whistles are cohesion calls. Anim. Behav. 1998, 56, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.J.; Sayigh, L.S. Geographic variation in rates of vocal production of free-ranging bottlenose dolphins. Mar. Mammal Sci. 2002, 18, 374–393. [Google Scholar] [CrossRef]
- Janik, V.M.; Sayigh, L.S. Communication in bottlenose dolphins: 50 years of signature whistle research. J. Comp. Physiol. A 2013, 199, 479–489. [Google Scholar] [CrossRef]
- King, S.L.; Sayigh, L.S.; Wells, R.S.; Fellner, W.; Janik, V.M. Vocal copying of individually distinctive signature whistles in bottlenose dolphins. Proc. R. Soc. Lond. 2013, 280, 20130053. [Google Scholar] [CrossRef]
- Caldwell, M.C.; Caldwell, D.K.; Tyack, P.L. Review of the signature-whistle hypothesis for the Atlantic bottlenose dolphin. In The Bottlenose Dolphin; Leatherwood, S., Reeves, R.R., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 199–234. [Google Scholar] [CrossRef]
- Quick, N.J.; Janik, V.M. Bottlenose dolphins exchange signature whistles when meeting at sea. Proc. R. Soc. B Biol. Sci. 2012, 279, 2539–2545. [Google Scholar] [CrossRef]
- Janik, V.M.; Todt, D.; Dehnhardt, G. Signature whistle variations in a bottlenosed dolphin, Tursiops truncatus. Behav. Ecol. Sociobiol. 1994, 35, 243–248. [Google Scholar] [CrossRef]
- Heiler, J.; Elwen, S.; Kriesell, H.; Gridley, T. Changes in bottlenose dolphin whistle parameters related to vessel presence, surface behaviour and group composition. Anim. Behav. 2016, 117, 167–177. [Google Scholar] [CrossRef]
- Blomqvist, C.; Amundin, M. High-frequency burst-pulse sounds in agonistic/aggressive interactions in bottlenose dolphins, Tursiops truncatus. In Echolocation in Bats and Dolphins; Thomas, J., Moss, C., Vater, M., Eds.; The University of Chicago Press: Chicago, IL, USA, 2004; pp. 425–431. [Google Scholar]
- Gridley, T.; Nastasi, A.; Kriesell, H.; Elwen, S. The acoustic repertoire of wild common bottlenose dolphins (Tursiops truncatus) in Walvis Bay, Namibia. Bioacoustics 2015, 24, 153–174. [Google Scholar] [CrossRef]
- Caldwell, M.C.; Caldwell, D.K. Dolphin community life. Quarterly 1967, 5, 12–15. [Google Scholar]
- Jones, B.; Zapetis, M.; Samuelson, M.M.; Ridgway, S. Sounds produced by bottlenose dolphins (Tursiops): A review of the defining characteristics and acoustic criteria of the dolphin vocal repertoire. Bioacoustics 2020, 29, 399–440. [Google Scholar] [CrossRef]
- Caldwell, M.C.; Haugen, R.M.; Caldwell, D.K. High-energy sounds associated with fright in the dolphin. Science 1962, 138, 907–908. [Google Scholar] [CrossRef]
- Sayigh, L.; Dziki, A.; Janik, V.; Kim, E.; McHugh, K.; Tyack, P.L.; Wells, R.; Jensen, F.H. Non-whistle sounds used in bottlenose dolphin aggressive interactions recorded on digital acoustic tags. Acoust. Soc. Am. 2017, 141, 3947. [Google Scholar] [CrossRef]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Firkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef]
- Brudzynski, S.M. Vocalizations as indicators of emotional states in rats and cats. In Evolution of Emotional Communication: From Sounds in Nonhuman Mammals to Speech and Music in Man; Altenmüller, E., Schmidt, S., Zimmermann, E., Eds.; Oxford University Press: Oxford UK, 2013; pp. 75–91. [Google Scholar]
- Soltis, J.; Leong, K.; Savage, A. African elephant vocal communication II: Rumble variation reflects the individual identity and emotional state of callers. Anim. Behav. 2005, 70, 589–599. [Google Scholar] [CrossRef]
- Castellote, M.; Fossa, F. Measuring acoustic activity as a method to evaluate welfare in captive beluga whales (Delphinapterus leucas). Aquat. Mamm. 2006, 32, 325–333. [Google Scholar] [CrossRef]
- Leliveld, L.M.; Düpjan, S.; Tuchscherer, A.; Puppe, B. Vocal correlates of emotional reactivity within and across contexts in domestic pigs (Sus scrofa). Physiol. Behav. 2017, 181, 117–126. [Google Scholar] [CrossRef]
- Watts, J.M.; Stookey, J.M. Vocal behaviour in cattle: The animal’s commentary on its biological processes and welfare. Appl. Anim. Behav. Sci. 2000, 67, 15–33. [Google Scholar] [CrossRef]
- Gridley, T.; Elwen, S.H.; Harris, G.; Moore, D.M.; Hoelzel, A.R.; Lampen, F. Hybridization in bottlenose dolphins—A case study of Tursiops aduncus× T. truncatus hybrids and successful backcross hybridization events. PLoS ONE 2018, 13, e0201722. [Google Scholar] [CrossRef]
- Joseph, C.A. An Assessment of Behaviour, Stress and Gastric Ulceration in Bottlenose Dolphins, Tursiops sp., at uShaka Sea World. Masters’ Thesis, University of KwaZulu-Natal, Durban, South Africa, 2020. [Google Scholar]
- Jensen, A.-L.M.; Delfour, F.; Carter, T. Anticipatory behaviour in captive bottlenose dolphins (Tursiops truncatus): A preliminary study. Zoo Biol. 2013, 32, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Center for Conservation Bioacoustics. Raven Pro: Interactive Sound Analysis Software, Version 1.6.1; The Cornell Lab of Ornithology: Ithica, NY, USA, 2019. Available online: http://ravensoundsoftware.com/(accessed on 28 October 2023).
- Gridley, T.; Berggren, P.; Cockcroft, V.G.; Janik, V.M. Whistle vocalizations of Indo-Pacific bottlenose dolphins (Tursiops aduncus) inhabiting the south-west Indian Ocean. J. Acoust. Soc. Am. 2012, 132, 4032–4040. [Google Scholar] [CrossRef]
- King, S.L.; Harley, H.E.; Janik, V.M. The role of signature whistle matching in bottlenose dolphins, Tursiops truncatus. Anim. Behav. 2014, 96, 79–86. [Google Scholar] [CrossRef]
- Kriesell, H.J.; Elwen, S.H.; Nastasi, A.; Gridley, T. Identification and characteristics of signature whistles in wild bottlenose dolphins (Tursiops truncatus) from Namibia. PLoS ONE 2014, 9, e106317. [Google Scholar] [CrossRef]
- Janik, V.M. Whistle matching in wild bottlenose dolphins (Tursiops truncatus). Science 2000, 289, 1355–1357. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 28 October 2023).
- Araya-Salas, M.; Smith-Vidaurre, G. warbleR: An r package to streamline analysis of animal acoustic signals. Methods Ecol. Evol. 2017, 8, 184–191. [Google Scholar] [CrossRef]
- Ligges, U.; Krey, S.; Mersmann, O.; Schnackenberg, S. tuneR: Analysis of Music and Speech. 2018. Available online: https://CRAN.R-project.org/package=tuneR (accessed on 28 October 2023).
- Sueur, J.; Aubin, T.; Simonis, C. Seewave: A free modular tool for sound analysis and synthesis. Bioacoustics 2008, 18, 213–226. [Google Scholar] [CrossRef]
- Dinno, A. Conover.test: Conover-Iman Test of Multiple Comparisons Using Rank Sums. R Package Version 1.1.5. 2017. Available online: https://CRAN.R-project.org/package=conover.test (accessed on 28 October 2023).
- Platto, S.; Serres, A. A quantitative assessment of trainers-dolphins’ interactions in the absence of food reward. Animals 2023, 13, 1651. [Google Scholar] [CrossRef]
- Clegg, I.L.; Rödel, H.G.; Boivin, X.; Delfour, F. Looking forward to interacting with their caretakers: Dolphins’ anticipatory behaviour indicates motivation to participate in specific events. Appl. Anim. Behav. Sci. 2018, 202, 85–93. [Google Scholar] [CrossRef]
- Spruijt, B.M.; Van den Bos, R.; Pijlman, F. A concept of welfare based on how the brain evaluates its own activity: Anticipatory behaviour as an indicator for this activity. Appl. Anim. Behav. Sci. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Galhardo, L.; Appleby, M.C.; Waran, N.K.; Dos Santos, M.E. Spontaneous activities of captive performing bottlenose dolphins (Tursiops truncatus). Anim. Welf. 1996, 5, 373–389. [Google Scholar] [CrossRef]
- Díaz López, B.D. When personality matters: Personality and social structure in wild bottlenose dolphins, Tursiops truncatus. Anim. Behav. 2020, 163, 73–84. [Google Scholar] [CrossRef]
- Vazire, S.; Gosling, S.D. Personality and temperament. In Encyclopedia of Animal Behavior; Bekoff, M., Ed.; Greenwood Press: Westport, UK, 2004; pp. 818–822. [Google Scholar]
- Hertel, A.G.; Niemelä, P.T.; Dingemanse, N.J.; Mueller, T. A guide for studying among-individual behavioral variation from movement data in the wild. Mov. Ecol. 2020, 8, 30. [Google Scholar] [CrossRef]
- Fischer, J.; Hammerschmidt, K.; Todt, D. Factors affecting acoustic variation in Barbary-macaque (Macaca sylvanus) disturbance calls. Ethology 1995, 101, 51–66. [Google Scholar] [CrossRef]
- Anzai, H.; Hirata, M. Individual monitoring of behavior to enhance productivity and welfare of animals in small-scale intensive cattle grazing systems. Front. Sustain. Food Syst. 2021, 5, 694413. [Google Scholar] [CrossRef]
- Stoinski, T.S.; Jaicks, H.F.; Drayton, L.A. Visitor effects on the behavior of captive western lowland gorillas: The importance of individual differences in examining welfare. Zoo Biol. 2012, 31, 586–599. [Google Scholar] [CrossRef]
- Whitham, J.C.; Wielebnowski, N. Animal-based welfare monitoring: Using keeper ratings as an assessment tool. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2009, 28, 545–560. [Google Scholar] [CrossRef]
- Scarpaci, C.; Bigger, S.W.; Corkeron, P.J.; Nugegoda, D. Bottlenose dolphins (Tursiops truncatus) increase whistling in the presence of ‘swim-with-dolphin’ tour operations. J. Cetacean Res. Manag. 2000, 2, 183–185. [Google Scholar] [CrossRef]
- Pongrácz, P.; Molnár, C.; Miklósi, A.; Csányi, V. Human listeners are able to classify dog (Canis familiaris) barks recorded in different situations. J. Comp. Psychol. 2005, 119, 136. [Google Scholar] [CrossRef]
- McGovern, B. Flexible Information within the Communication Signals of Wild, Provisioned Indo-Pacific Bottlenose Dolphins. Doctoral Dissertation, University of Queensland, St Lucia, QLD, Australia, 2022. [Google Scholar]
- Serres, A.; Xu, C.; Hao, Y.; Wang, D. The click production of captive yangtze finless porpoises (Neophocaena asiaeorientalis asiaorientalis) is influenced by social and environmental factors. Animals 2021, 11, 511. [Google Scholar] [CrossRef]
- Lopez-Marulanda, J.; Rödel, H.G.; Colpaert, N.; Vanderheul, S.; Adam, O.; Delfour, F. Contexts of emission of non-signature whistles in bottlenose dolphins (Tursiops truncatus) under human care. Behav. Process. 2020, 181, 104255. [Google Scholar] [CrossRef] [PubMed]
- Overstrom, N.A. Association between burst-pulse sounds and aggressive behavior in captive Atlantic bottlenosed dolphins (Tursiops truncatus). Zoo Biol. 1983, 2, 93–103. [Google Scholar] [CrossRef]
- Meagher, R.K. Is boredom an animal welfare concern? Anim. Welf. 2019, 28, 21–32. [Google Scholar] [CrossRef]
- Dawkins, M.S. Behaviour as a tool in the assessment of animal welfare. Zoology 2003, 106, 383–387. [Google Scholar] [CrossRef]
- Buchanan-Smith, H.M.; Griciute, J.; Daoudi, S.; Leonardi, R.; Whiten, A. Interspecific interactions and welfare implications in mixed species communities of capuchin (Sapajus apella) and squirrel monkeys (Saimiri sciureus) over 3 years. Appl. Anim. Behav. Sci. 2013, 147, 324–333. [Google Scholar] [CrossRef]
- Mason, G.; Clubb, R.; Latham, N.; Vickery, S. Why and how should we use environmental enrichment to tackle stereotypic behaviour? Appl. Anim. Behav. Sci. 2007, 102, 163–188. [Google Scholar] [CrossRef]
- Grandin, T. The feasibility of using vocalization scoring as an indicator of poor welfare during cattle slaughter. Appl. Anim. Behav. Sci. 1998, 56, 121–128. [Google Scholar] [CrossRef]
- Cordeiro, A.F.; Nääs, I.D.; Oliveira, S.R.; Violaro, F.; Almeida, A.C.; Neves, D.P. Understanding vocalization might help to assess stressful conditions in piglets. Animals 2013, 3, 923–934. [Google Scholar] [CrossRef]
- Sadeghi, M.; Banakar, A.; Khazaee, M.; Soleimani, M.R. An intelligent procedure for the detection and classification of chickens infected by clostridium perfringens based on their vocalization. Braz. J. Poult. Sci. 2015, 17, 537–544. [Google Scholar] [CrossRef]
- Jones, B.L.; Oswald, M.; Tufano, S.; Baird, M.; Mulsow, J.; Ridgway, S.H. A system for monitoring acoustics to supplement an animal welfare plan for bottlenose dolphins. J. Zool. Bot. Gard. 2021, 2, 222–233. [Google Scholar] [CrossRef]
- Buckstaff, K.C. Effects of watercraft noise on the acoustic behavior of bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar. Mammal Sci. 2004, 20, 709–725. [Google Scholar] [CrossRef]
- King, S.; Guarino, E.; Donegan, K.; Hecksher, J.; Jaakkola, K. Further insights into postpartum signature whistle use in bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 2016, 32, 1458–1469. [Google Scholar] [CrossRef]
- Frohoff, T.G. Behavior of Captive Bottlenose Dolphins (Tursiops truncatus) and Humans during Controlled in-Water Interactions. Doctoral Dissertation, Texas A & M University, College Station, TX, USA, 1993. [Google Scholar]
- Luís, A.R.; Couchinho, M.N.; Dos Santos, M.E. Signature whistles in wild bottlenose dolphins: Long-term stability and emission rates. Acta Etholog. 2016, 19, 113–122. [Google Scholar] [CrossRef]
- Quick, N.J.; Janik, V.M. Whistle rates of wild bottlenose dolphins (Tursiops truncatus): Influences of group size and behavior. J. Comp. Psychol. 2008, 122, 305. [Google Scholar] [CrossRef]
- Dos Santos, M.E.D.; Louro, S.; Couchinho, M.N.; Brito, C.M. Whistles of bottlenose dolphins (Tursiops truncatus) in the Sado Estuary, Portugal: Characteristics, production rates, and long-term contour stability. Aquat. Mamm. 2005, 31, 453–462. [Google Scholar] [CrossRef]
- La Manna, G.; Rako-Gòspic, N.; Manghi, M.; Ceccherelli, G. Influence of environmental, social and behavioural variables on the whistling of the common bottlenose dolphin (Tursiops truncatus). Behav. Ecol. Sociobiol. 2019, 73, 121. [Google Scholar] [CrossRef]
- Díaz López, B. Whistle characteristics in free-ranging bottlenose dolphins (Tursiops truncatus) in the Mediterranean Sea: Influence of behaviour. Mamm. Biol. 2011, 76, 180–189. [Google Scholar] [CrossRef]
- Lemon, M.; Cato, D.H.; Lynch, T.P.; Harcourt, R.G. Variation in sound production in relation to behavior of bottlenose dolphins (Tursiops aduncus). J. Acoust. Soc. Am. 2006, 120, 3014. [Google Scholar] [CrossRef]






| Context (Year; No. of Sessions) | Time of Day | Details | Trainers’ Activities | Dolphins’ Activities |
|---|---|---|---|---|
| Night (2018; 12 randomly selected from 24) | 20:00–03:00 | Lights were off, dolphinarium was quiet | Absent from facility | Minimal to no human interactions * |
| Daily activities (2010, 2016; 8) | 09:30–16:00 | All other normal daytime dolphinarium regimes excluding the morning feeding session and presentations | Interactions included training/feeding sessions with the dolphins | Involved in training sessions, feeding sessions or at rest |
| 20–10 min pre (2010, 2016; 6) | 10–20 min before presentations | The 10 min period leading up to presentation preparation | Starting preparation for the presentations (preparing boxes of fish, more staff activity around the presentation pool) | Starting to get prepared for the presentation |
| 10–0 min pre (2010, 2016; 6) | Within 10 min before presentations | The period immediately before and within 10 min preceding presentations. | Preparing for the presentation (sound/music checks, moving dolphins around to different pools) | Moved between pools in preparation for the presentation |
| Presentation (2010, 2016; 11) | 10:30, 12:30 and 15:00 | Included one to five dolphins at a time. Music playing. | Participating in presentations and back-of-house preparing to shift dolphins around | Either participating in the presentation or back-of-house |
| 0–10 min post (2010, 2016; 7) | Within 10 min after presentations | The period immediately after and up to 10 min following presentations. Music stops playing. | Moving dolphins around to designated pools | All dolphins were reunited, within their social groups, back-of-house and moved around to designated pools |
| 10–20 min post (2010, 2016; 7) | 10–20 min after presentations | The 10 min period after which all dolphins are reunited back of house | Either preparing for further training sessions or resting periods | All dolphins were moved into their designated pools |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Probert, R.; James, B.S.; Elwen, S.H.; Gridley, T. Vocal Cues to Assess Arousal State of Bottlenose Dolphins (Tursiops spp.) Involved in Public Presentations. J. Zool. Bot. Gard. 2023, 4, 711-727. https://doi.org/10.3390/jzbg4040050
Probert R, James BS, Elwen SH, Gridley T. Vocal Cues to Assess Arousal State of Bottlenose Dolphins (Tursiops spp.) Involved in Public Presentations. Journal of Zoological and Botanical Gardens. 2023; 4(4):711-727. https://doi.org/10.3390/jzbg4040050
Chicago/Turabian StyleProbert, Rachel, Bridget S. James, Simon H. Elwen, and Tess Gridley. 2023. "Vocal Cues to Assess Arousal State of Bottlenose Dolphins (Tursiops spp.) Involved in Public Presentations" Journal of Zoological and Botanical Gardens 4, no. 4: 711-727. https://doi.org/10.3390/jzbg4040050