
Anti-Inflammatory Efficacy of Resveratrol-Enriched Rice Callus Extract on Lipopolysaccharide-Stimulated RAW264.7 Macrophages

Abstract
:
1. Introduction
2. Materials and Methods
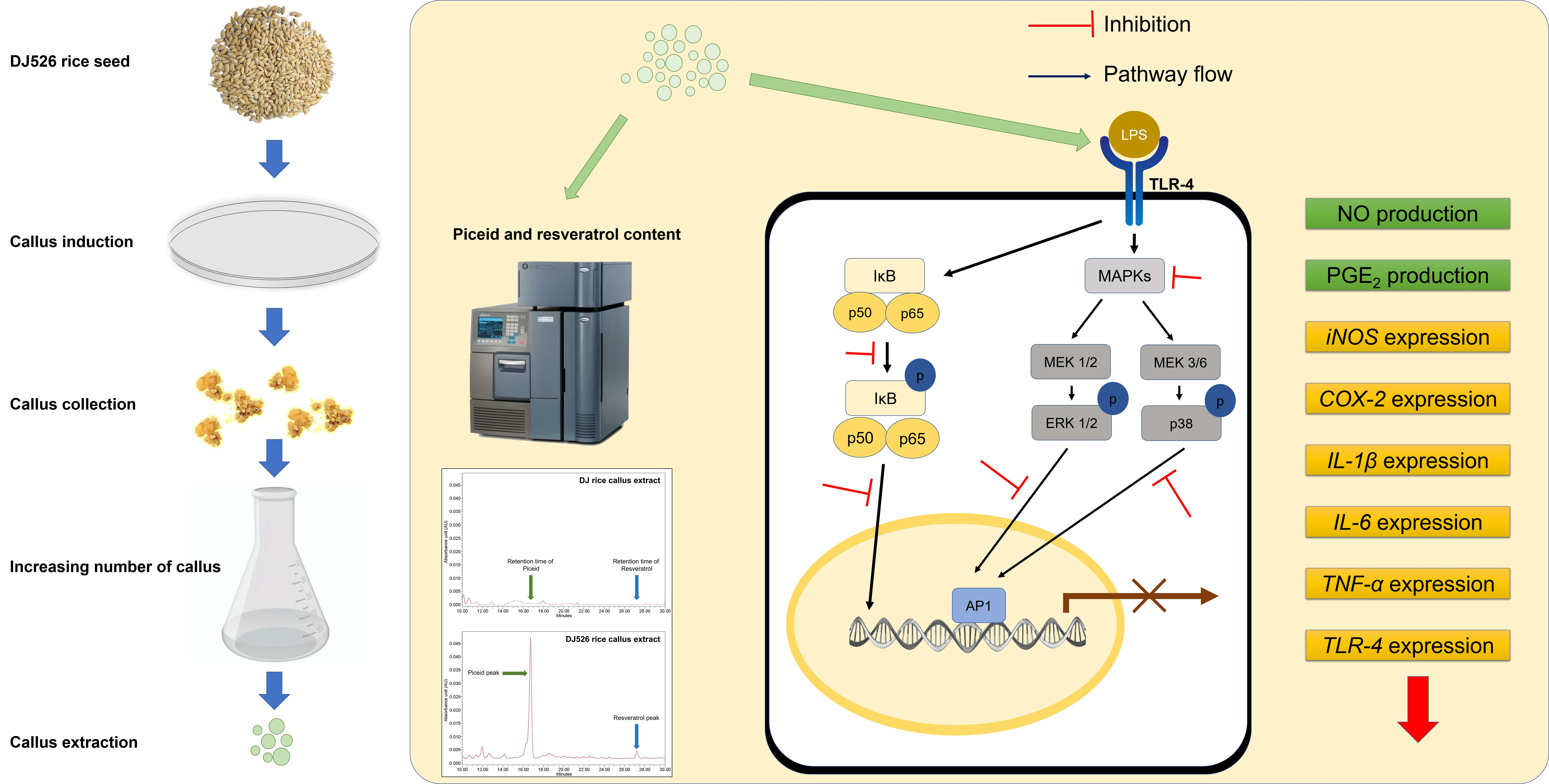
2.1. Callus Culture
2.2. Extraction of Resveratrol-Enrich Compound from Rice Callus
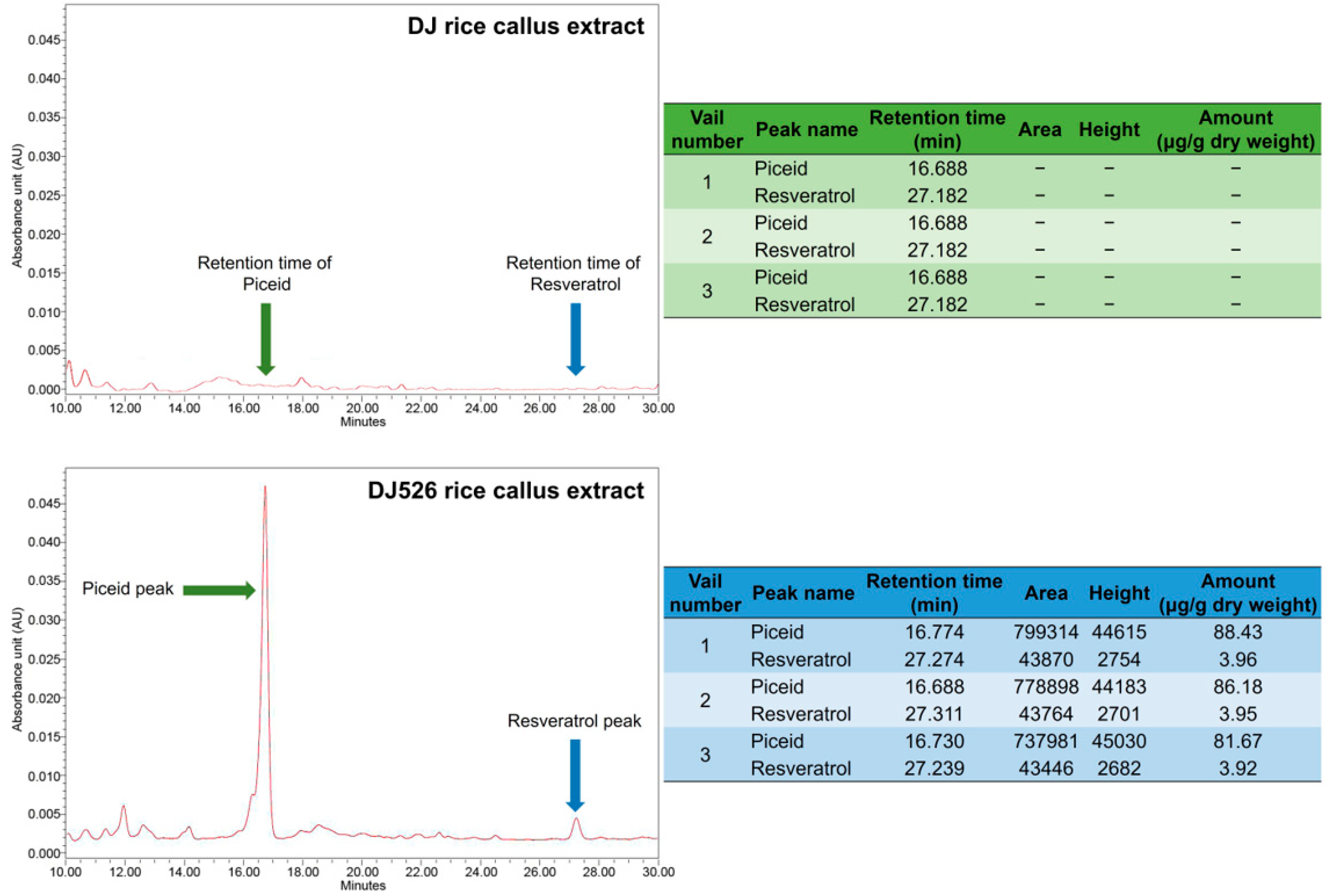
2.3. Quantification of Piceid and Resveratrol Content Using High-Performance Liquid Chromatography (HPLC) Analysis
2.4. RAW264.7 Cell Culture
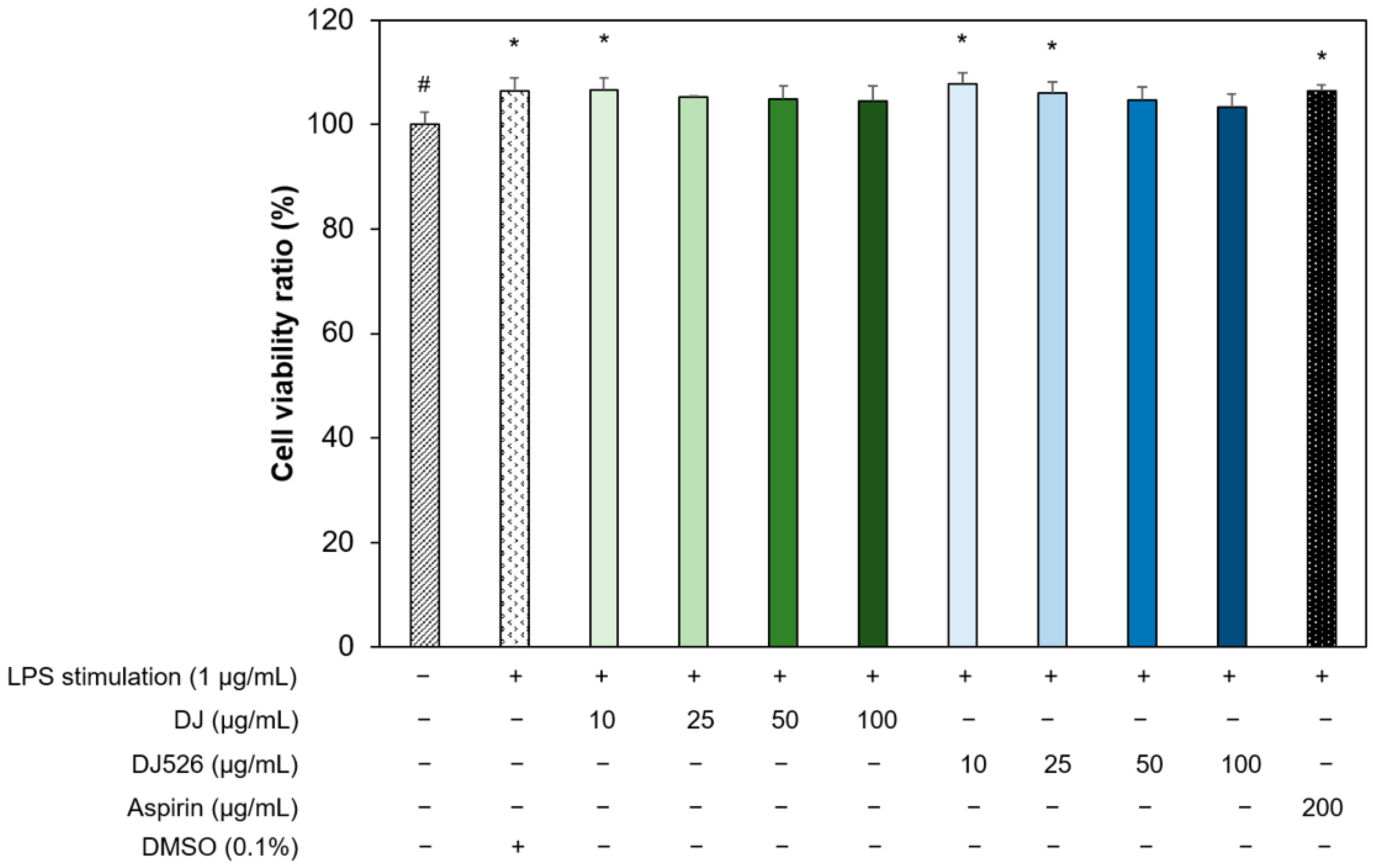
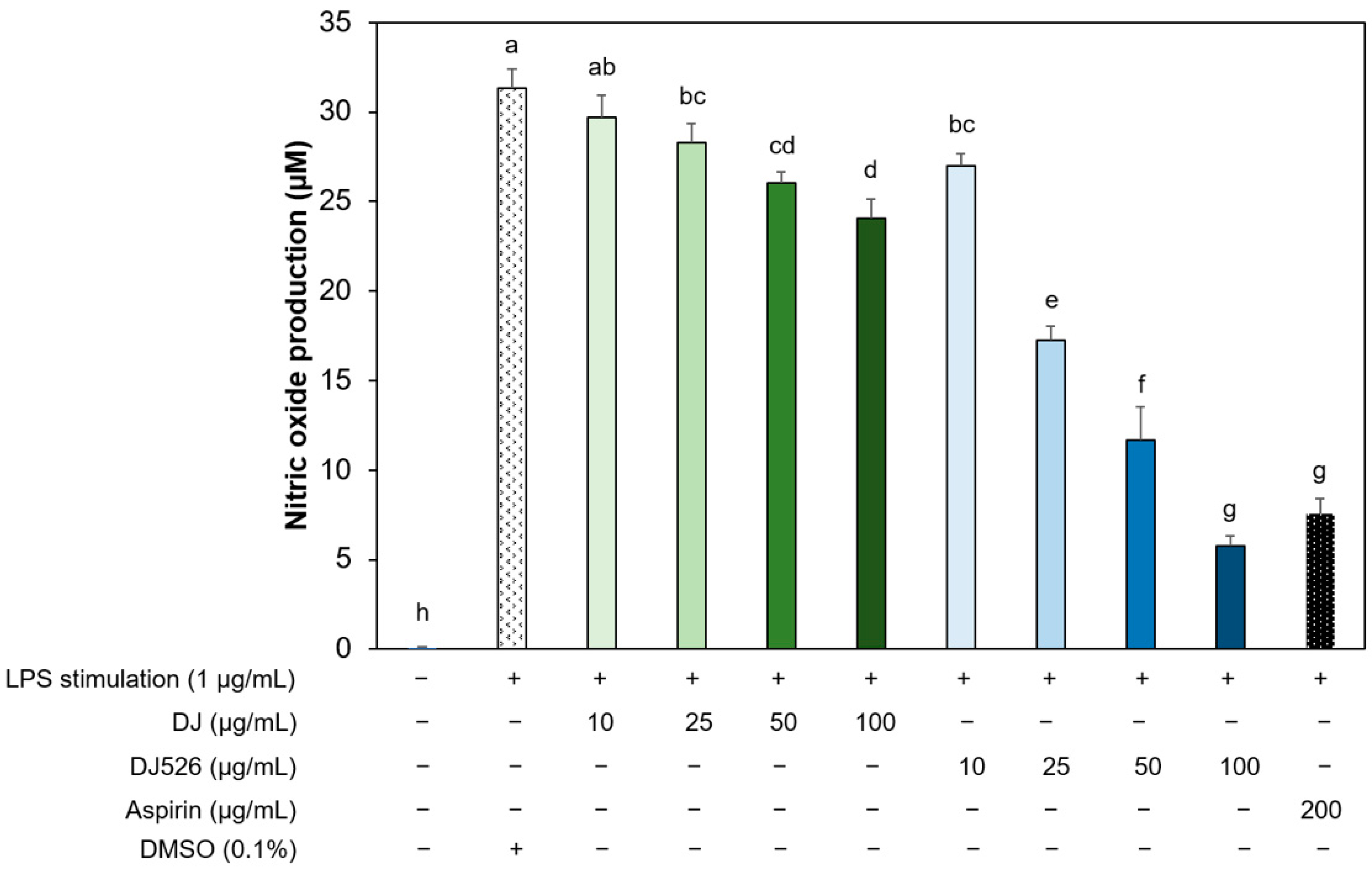
2.5. Cell Viability and Nitric Oxide Production Analysis
2.6. RNA Isolation, RNA Quantification, and cDNA Synthesis
2.7. Measurement of mRNA from Inflammatory Genes by RT-qPCR
2.8. Prostaglandin E2 (PGE2) Production
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. Piceid and Resveratrol Contents in Rice Callus Extracts
3.2. Effects of Resveratrol-Enriched Rice Callus Extract on the Viability of LPS-Stimulated RAW264.7 Cells
3.3. Resveratrol-Enriched Rice Callus Extract Inhibited LPS-Induced NO Production by RAW264.7 Cells
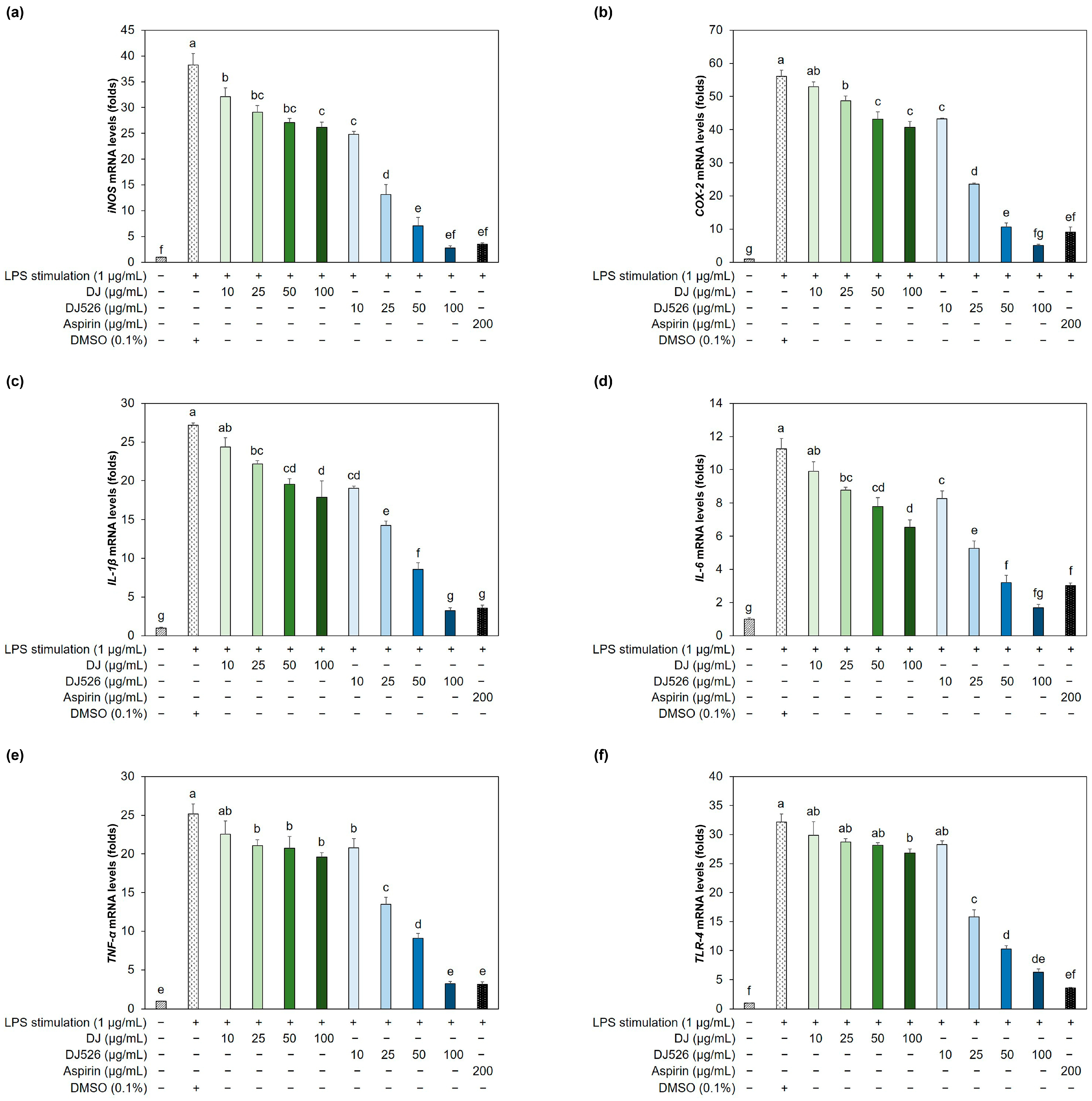
3.4. Resveratrol-Enriched Rice Callus Extract Suppressed LPS-Induced Upregulation of Multiple Proinflammatory Genes in RAW264.7 Cells
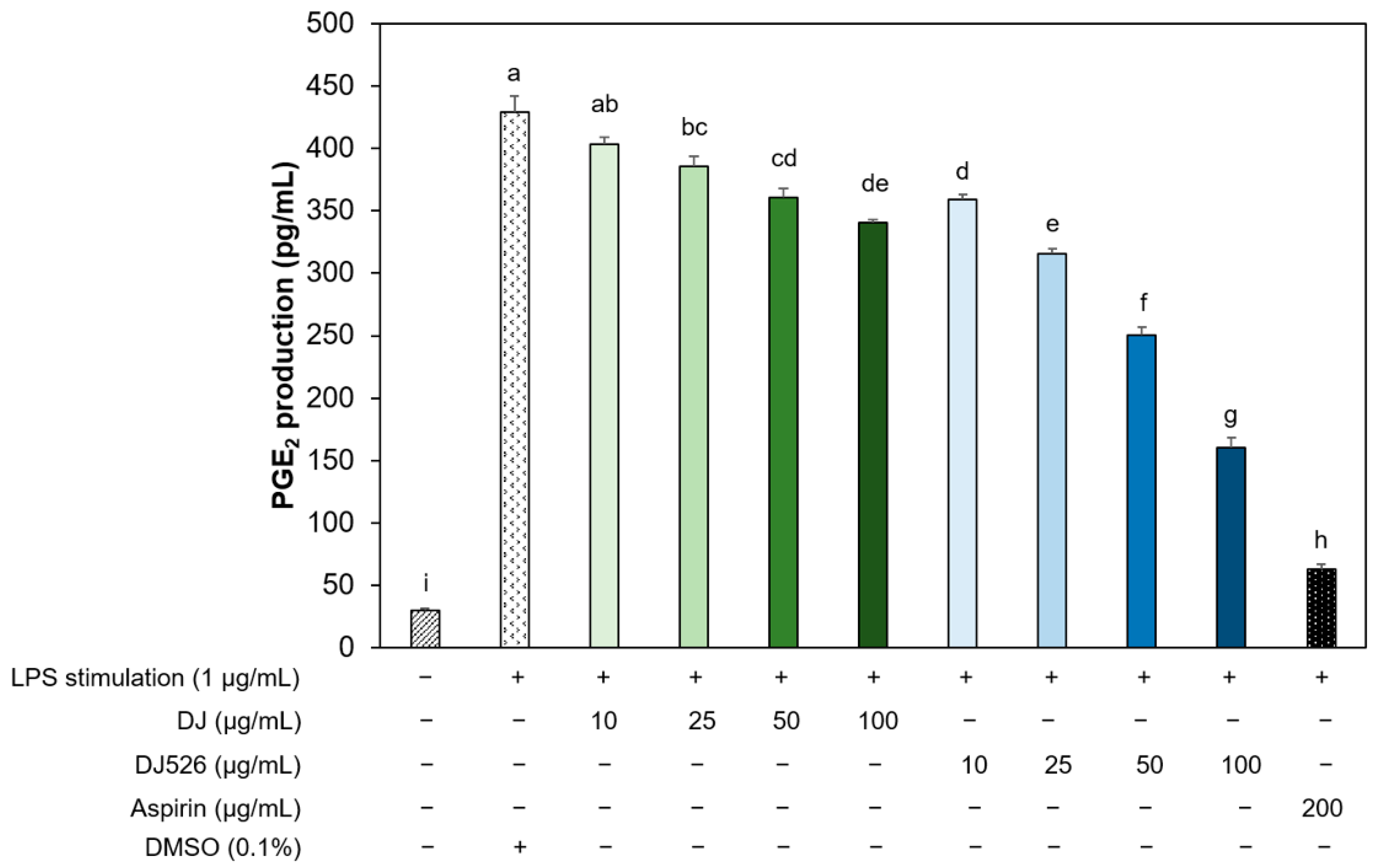
3.5. Resveratrol-Enriched Rice Callus Extract Reduced the Production of PGE2 by LPS-Stimulated RAW264.7 Cells
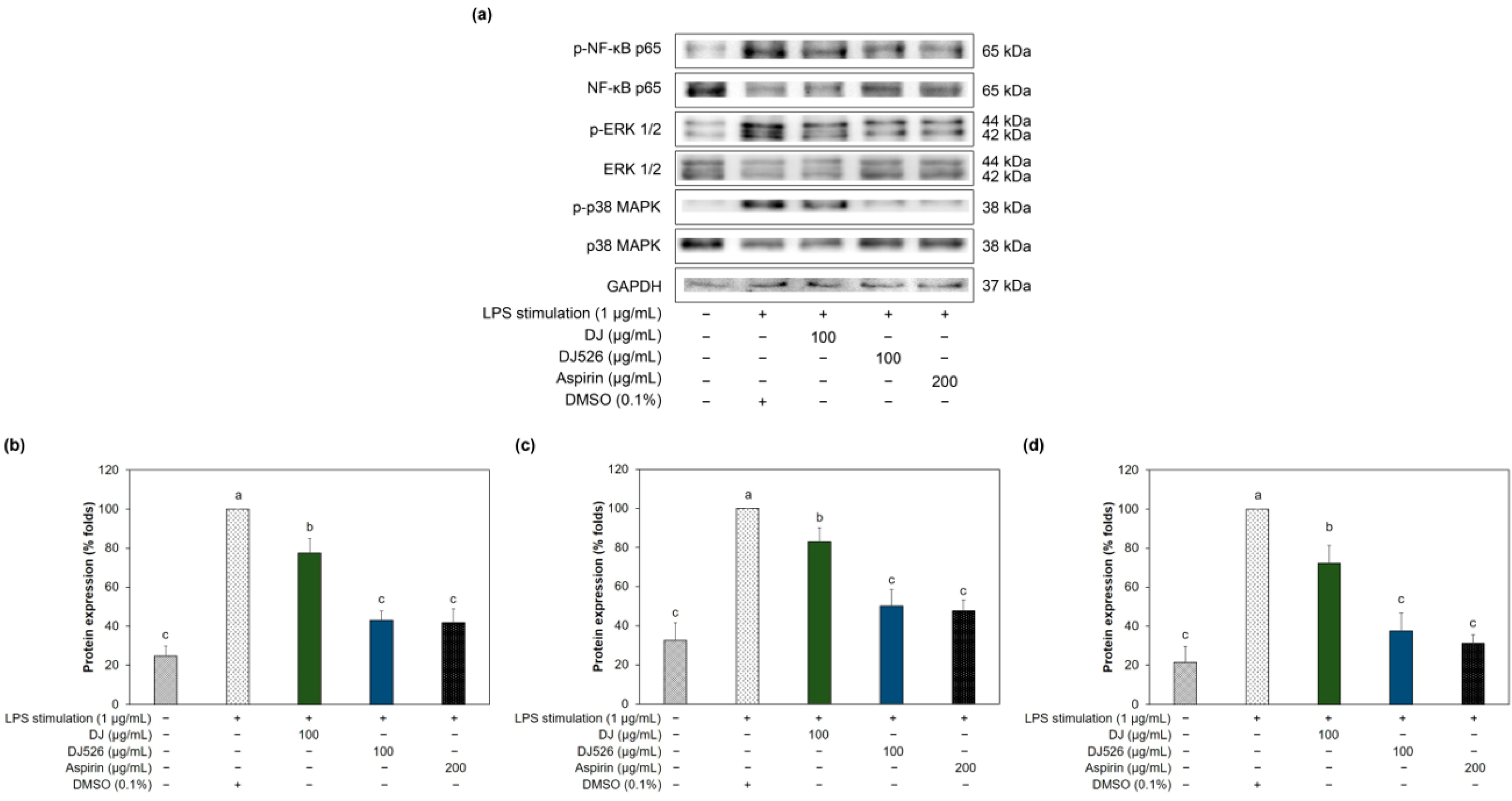
3.6. Resveratrol-Enriched Rice Callus Extract Inhibited MAPK and NF-κB Pathway Activation in LPS-Stimulated RAW264.7 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Zhang, H.; Liu, Z.; Ma, C.; Kang, W. Immunomodulation of ADPs-1a and ADPs-3a on RAW264.7 cells through NF-κB/MAPK signaling pathway. Int. J. Biol. Macromol. 2019, 132, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yin, S.; Chen, Y.; Wu, Y.; Zheng, W.; Dong, H.; Bai, Y.; Qin, Y.; Li, J.; Feng, S.; et al. LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Mol. Med. Rep. 2018, 17, 5484–5491. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Chen, Z.; Han, P.; Hu, Q.; Xiao, Y. Activation of macrophages by lipopolysaccharide for assessing the immunomodulatory property of biomaterials. Tissue Eng. Part. A 2017, 23, 1100–1109. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Hu, S.; Su, Z.; Wang, Q.; Meng, G.; Guo, T.; Zhang, J.; Gao, P. Myricetin attenuates LPS-induced inflammation in RAW 264.7 macrophages and mouse models. bioRxiv 2018, 299420. [Google Scholar] [CrossRef] [PubMed]
- Ngkelo, A.; Meja, K.; Yeadon, M.; Adcock, I.; Kirkham, P.A. LPS induced inflammatory responses in human peripheral blood mononuclear cells is mediated through NOX4 and Giα dependent PI-3kinase signalling. J. Inflamm. 2012, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Raduolovic, K.; Mak’Anyengo, R.; Kaya, B.; Steinert, A.; Niess, J.H. Injections of lipopolysaccharide into mice to mimic entrance of microbial-derived products after intestinal barrier breach. J. Vis. Exp. 2018, 135, e57610. [Google Scholar] [CrossRef]
- Tucureanu, M.M.; Rebleanu, D.; Constantinescu, C.A.; Deleanu, M.; Voicu, G.; Butoi, E.; Calin, M.; Manduteanu, I. Lipopolysaccharide-induced inflammation in monocytes/macrophages is blocked by liposomal delivery of G(i)-protein inhibitor. Int. J. Nanomed. 2018, 13, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Lull, C.; Wichers, H.J.; Savelkoul, H.F.J. Antiinflammatory and immunomodulating properties of fungal metabolites. Mediat. Inflamm. 2005, 2005, 892572. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, X.; Qu, Y.; Wang, L.; Geng, D.; Chen, W.; Li, L.; Tian, Y.; Chang, S.; Zhao, C.; et al. The roles of p38 MAPK → COX2 and NF-κB → COX2 signal pathways in age-related testosterone reduction. Sci. Rep. 2019, 9, 10556. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Kim, M.J.; Li, Y.; Park, Y.N.; Lee, J.; Lee, Y.J.; Kim, S.G.; Park, H.J.; Son, J.K.; Chang, H.W.; et al. Britanin suppresses LPS-induced nitric oxide, PGE2 and cytokine production via NF-κB and MAPK inactivation in RAW 264.7 cells. Int. Immunopharmacol. 2013, 15, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Athar, M.; Back, J.H.; Tang, X.; Kim, K.H.; Kopelovich, L.; Bickers, D.R.; Kim, A.L. Resveratrol: A review of preclinical studies for human cancer prevention. Toxicol. Appl. Pharmacol. 2007, 224, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Szajdek, A.; Borowska, E.J. Bioactive compounds and health-promoting properties of berry fruits: A review. Plant Foods Hum. Nutr. 2008, 63, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.S.; Salve, A.R.; Chauhan, S. Peanuts as functional food: A review. J. Food Sci. Technol. 2016, 53, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Sebastià, N.; Montoro, A.; Mañes, J.; Soriano, J.M. A preliminary study of presence of resveratrol in skins and pulps of European and Japanese plum cultivars. J. Sci. Food Agric. 2012, 92, 3091–3094. [Google Scholar] [CrossRef] [PubMed]
- Banez, M.J.; Geluz, M.I.; Chandra, A.; Hamdan, T.; Biswas, O.S.; Bryan, N.S.; Von Schwarz, E.R. A systemic review on the antioxidant and anti-inflammatory effects of resveratrol, curcumin, and dietary nitric oxide supplementation on human cardiovascular health. Nutr. Res. 2020, 78, 11–26. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Durazzo, A.; Lucarini, M.; Souto, E.B.; Santini, A.; Imran, M.; Moussa, A.Y.; Mostafa, N.M.; El-Shazly, M.; et al. Resveratrol’ biotechnological applications: Enlightening its antimicrobial and antioxidant properties. J. Herb. Med. 2022, 32, 100550. [Google Scholar] [CrossRef]
- Vestergaard, M.; Ingmer, H. Antibacterial and antifungal properties of resveratrol. Int. J. Antimicrob. Agents 2019, 53, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Ferraz da Costa, D.C.; Pereira Rangel, L.; Martins-Dinis, M.M.; Ferretti, G.D.; Ferreira, V.F.; Silva, J.L. Anticancer potential of resveratrol, β-lapachone and their analogues. Molecules 2020, 25, 893. [Google Scholar] [CrossRef] [PubMed]
- Angellotti, G.; Di Prima, G.; Belfiore, E.; Campisi, G.; De Caro, V. Chemopreventive and anticancer role of resveratrol against oral squamous cell carcinoma. Pharmaceutics 2023, 15, 275. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-inflammatory action and mechanisms of resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Z.; Yao, L.; Jiao, M.M.; Shi, J.B.; Tan, Y.; Ruan, B.F.; Liu, X.H. Novel resveratrol-based flavonol derivatives: Synthesis and anti-inflammatory activity in vitro and in vivo. Eur. J. Med. Chem. 2019, 175, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-A.; Chen, C.-S.; Wang, Y.-C.; Lin, E.-S.; Chang, C.-Y.; Chen, J.J.; Wu, M.-Y.; Lin, H.-J.; Wan, L. Anti-inflammatory effects of resveratrol on human retinal pigment cells and a myopia animal model. Curr. Issues Mol. Biol. 2021, 43, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Griñán-Ferré, C.; Bellver-Sanchis, A.; Izquierdo, V.; Corpas, R.; Roig-Soriano, J.; Chillón, M.; Andres-Lacueva, C.; Somogyvári, M.; Sőti, C.; Sanfeliu, C.; et al. The pleiotropic neuroprotective effects of resveratrol in cognitive decline and Alzheimer’s disease pathology: From antioxidant to epigenetic therapy. Ageing Res. Rev. 2021, 67, 101271. [Google Scholar] [CrossRef] [PubMed]
- Sawda, C.; Moussa, C.; Turner, R.S. Resveratrol for Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2017, 1403, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Arbo, B.D.; André-Miral, C.; Nasre-Nasser, R.G.; Schimith, L.E.; Santos, M.G.; Costa-Silva, D.; Muccillo-Baisch, A.L.; Hort, M.A. Resveratrol derivatives as potential treatments for Alzheimer’s and Parkinson’s disease. Front. Aging Neurosci. 2020, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.-M.; Zong, Y.; Sun, L.; Guo, J.-Z.; Zhang, W.; He, Y.; Song, R.; Wang, W.-M.; Xiao, C.-J.; Lu, D. Resveratrol Inhibits Inflammatory Responses via the Mammalian Target of Rapamycin Signaling Pathway in Cultured LPS-Stimulated Microglial Cells. PLoS ONE 2012, 7, e32195. [Google Scholar] [CrossRef]
- Pinheiro, D.M.L.; de Oliveira, A.H.S.; Coutinho, L.G.; Fontes, F.L.; de Medeiros Oliveira, R.K.; Oliveira, T.T.; Faustino, A.L.F.; Lira da Silva, V.; de Melo Campos, J.T.A.; Lajus, T.B.P.; et al. Resveratrol decreases the expression of genes involved in inflammation through transcriptional regulation. Free Radic. Biol. Med. 2019, 130, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Palomera-Ávalos, V.; Griñán-Ferré, C.; Izquierdo, V.; Camins, A.; Sanfeliu, C.; Canudas, A.M.; Pallàs, M. Resveratrol modulates response against acute inflammatory stimuli in aged mouse brain. Exp. Gerontol. 2018, 102, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.-H.; Shin, W.-C.; Ryu, H.-S.; Lee, D.-W.; Moon, E.; Seo, C.-S.; Hwang, E.; Lee, H.-S.; Ahn, M.-H.; Jeon, Y.; et al. Creation of resveratrol-enriched rice for the treatment of metabolic syndrome and related diseases. PLoS ONE 2013, 8, e57930. [Google Scholar] [CrossRef] [PubMed]
- Monmai, C.; Kim, J.-S.; Baek, S.-H. Use of germination to enhance resveratrol content and its anti-inflammatory activity in lipopolysaccharide-stimulated RAW264.7 cells. Molecules 2023, 28, 4898. [Google Scholar] [CrossRef] [PubMed]
- Cho, D.-H.; Lim, S.-T. Changes in phenolic acid composition and associated enzyme activity in shoot and kernel fractions of brown rice during germination. Food Chem. 2018, 256, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Park, S.; Kim, H.-J.; Lee, K.-J.; Kim, D.H.; Baek, S.-H.; Hong, S.-T. The resveratrol rice DJ526 callus significantly increases the lifespan of Drosophila (resveratrol rice DJ526 callus for longevity). Nutrients 2019, 11, 983. [Google Scholar] [CrossRef] [PubMed]
- Monmai, C.; Kim, J.-S.; Baek, S.-H. Transgenic rice seed extracts exert immunomodulatory effects by modulating immune-related biomarkers in RAW264.7 macrophage cells. Nutrients 2022, 14, 4143. [Google Scholar] [CrossRef] [PubMed]
- Kantayos, V.; Shin, W.-C.; Kim, J.-S.; Jeon, S.-H.; Rha, E.-S.; Baek, S.-H. Resveratrol-enriched rice identical to original Dongjin rice variety with respect to major agronomic traits in different cultivation years and regions. GM Crops Food 2021, 12, 449–458. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, S.; Li, X.; Feng, J.; Du, J.; Guo, L.; Su, Y.; Zhou, J.; Ding, G.; Bai, Y.; et al. Aspirin inhibits LPS-induced macrophage activation via the NF-κB pathway. Sci. Rep. 2017, 7, 11549. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, W.; Yue, Y.; Sun, C.; Zhang, Q. Proteoglycan from Bacillus sp. BS11 inhibits the inflammatory response by suppressing the MAPK and NF-κB pathways in lipopolysaccharide-induced RAW264.7 macrophages. Mar. Drugs 2020, 18, 585. [Google Scholar] [CrossRef] [PubMed]
- Adhami, V.M.; Afaq, F.; Ahmad, N. Suppression of ultraviolet B exposure-mediated activation of NF-κB in normal human keratinocytes by resveratrol. Neoplasia 2003, 5, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Pirola, L.; Fröjdö, S. Resveratrol: One molecule, many targets. IUBMB Life 2008, 60, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, F.; Wang, F.; Li, J.; Lin, C.; Du, J. Resveratrol inhibits the proliferation of A549 cells by inhibiting the expression of COX-2. OncoTargets Ther. 2018, 11, 2981–2989. [Google Scholar] [CrossRef]
- Magrone, T.; Magrone, M.; Russo, M.A.; Jirillo, E. Recent advances on the anti-Inflammatory and antioxidant properties of red grape polyphenols: In vitro and in vivo studies. Antioxidants 2020, 9, 35. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.W.; Fong, H.H.S.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Subbaramaiah, K.; Chung, W.J.; Michaluart, P.; Telang, N.; Tanabe, T.; Inoue, H.; Jang, M.; Pezzuto, J.M.; Dannenberg, A.J. Resveratrol inhibits cyclooxygenase-2 transcription and activity in phorbol ester-treated human mammary epithelial cells. J. Biol. Chem. 1998, 273, 21875–21882. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wu, F.; Jin, Z.; Zhai, Z.; Wang, Y.; Tu, B.; Yan, W.; Tang, T. Plumbagin inhibits LPS-induced inflammation through the inactivation of the nuclear factor-kappa B and mitogen activated protein kinase signaling pathways in RAW 264.7 cells. Food Chem. Toxicol. 2014, 64, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Wang, Y.; Dong, L.; Li, M.; Cai, W. Anti-inflammatory effect of resveratrol through the suppression of NF-κB and JAK/STAT signaling pathways. Acta Biochim. Biophys. Sin. 2015, 47, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Xiao, C.; Wei, Z.; Wang, J.; Yang, Z.; Fu, Y. Resveratrol inhibits LPS-induced mice mastitis through attenuating the MAPK and NF-κB signaling pathway. Microb. Pathog. 2017, 107, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Schmidt, S.V.; Sander, J.; Draffehn, A.; Krebs, W.; Quester, I.; De Nardo, D.; Gohel, T.D.; Emde, M.; Schmidleithner, L.; et al. Transcriptome-based network analysis reveals a spectrum model of human macrophage activation. Immunity 2014, 40, 274–288. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Lv, R.; Bao, Q.; Li, Y. Regulation of M1-type and M2-type macrophage polarization in RAW264.7 cells by Galectin-9. Mol. Med. Rep. 2017, 16, 9111–9119. [Google Scholar] [CrossRef]
- Lee, J.H.; Moon, S.H.; Kim, H.S.; Park, E.; Ahn, D.U.; Paik, H.D. Immune-enhancing activity of phosvitin by stimulating the production of pro-inflammatory mediator. Poult. Sci. 2017, 96, 3872–3878. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, K.A.; Amici, S.A.; Webb, L.M.; Ruiz-Rosado Jde, D.; Popovich, P.G.; Partida-Sanchez, S.; Guerau-de-Arellano, M. Novel markers to delineate murine M1 and M2 macrophages. PLoS ONE 2015, 10, e0145342. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Diao, P.; Shu, X.; Li, L.; Xiong, L. Quercetin and quercitrin attenuates the inflammatory response and oxidative stress in LPS-induced RAW264.7 cells: In vitro assessment and a theoretical model. Biomed. Res. Int. 2019, 2019, 7039802. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; Kell, D.B.; Pretorius, E. The role of lipopolysaccharide-induced cell signalling in chronic inflammation. Chronic Stress. 2022, 6, 24705470221076390. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin prevents inflammation in LPS-induced RAW264.7 macrophages by inhibiting FoxO3a/autophagy signaling pathway. Nutrients 2019, 11, 2794. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, N.; Fernandez, R. Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front. Cell Infect. Microbiol. 2013, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Zamyatina, A.; Heine, H. Lipopolysaccharide recognition in the crossroads of TLR4 and Caspase-4/11 mediated inflammatory pathways. Front. immunol 2020, 11, 585146. [Google Scholar] [CrossRef] [PubMed]
- Nijland, R.; Hofland, T.; van Strijp, J.A. Recognition of LPS by TLR4: Potential for anti-inflammatory therapies. Mar. Drugs 2014, 12, 4260–4273. [Google Scholar] [CrossRef] [PubMed]
- Eo, H.J.; Kwon, H.Y.; Da Kim, S.; Kang, Y.; Park, Y.; Park, G.H. GC/MS analysis and anti-inflammatory effect of leaf extracts from Hibiscus syriacus through inhibition of NF-κB and MAPKs signaling in LPS-stimulated RAW264.7 macrophages. Plant Biotechnol. Rep. 2020, 14, 539–546. [Google Scholar] [CrossRef]
- Zong, Y.; Sun, L.; Liu, B.; Deng, Y.S.; Zhan, D.; Chen, Y.L.; He, Y.; Liu, J.; Zhang, Z.J.; Sun, J.; et al. Resveratrol inhibits LPS-induced MAPKs activation via activation of the phosphatidylinositol 3-kinase pathway in murine RAW 264.7 macrophage cells. PLoS ONE 2012, 7, e44107. [Google Scholar] [CrossRef] [PubMed]
- Bigagli, E.; Cinci, L.; Paccosi, S.; Parenti, A.; D’Ambrosio, M.; Luceri, C. Nutritionally relevant concentrations of resveratrol and hydroxytyrosol mitigate oxidative burst of human granulocytes and monocytes and the production of pro-inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. Int. Immunopharmacol. 2017, 43, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Su, D.; Li, L.; Cai, H.; Zhang, M.; Zhai, J.; Li, M.; Wu, X.; Hu, K. Anti-inflammatory effects of Aureusidin in LPS-stimulated RAW264.7 macrophages via suppressing NF-κB and activating ROS- and MAPKs-dependent Nrf2/HO-1 signaling pathways. Toxicol. Appl. Pharmacol. 2020, 387, 114846. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Jia, Y.; Huang, L.; Wang, T.; Wang, H.; Dong, Y.; Zhang, H.; Fan, M.; Lv, P. Lipoxin A4 methyl ester ameliorates cognitive deficits induced by chronic cerebral hypoperfusion through activating ERK/Nrf2 signaling pathway in rats. Pharmacol. Biochem. Behav. 2014, 124, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Chen, X.; Song, X.; Chen, Y.; Jia, R.; Zou, Y.; Li, L.; Yin, L.; He, C.; Liang, X.; et al. Resveratrol inhibits LPS-induced inflammation through suppressing the signaling cascades of TLR4-NF-κB/MAPKs/IRF3. Exp. Ther. Med. 2020, 19, 1824–1834. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.G.; Kim, S.; Boo, K.-H.; Kim, J.-H.; Kim, C.S. Anti-inflammatory effects of immature Citrus unshiu fruit extracts via suppression of NF-κB and MAPK signal pathways in LPS-induced RAW264.7 macrophage cells. Food Sci. Biotechnol. 2023, 33, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ji, N.; Pan, S.; Zhang, Z.; Wang, R.; Qiu, Y.; Jin, M.; Kong, D. Roburic acid suppresses NO and IL-6 production via targeting NF-κB and MAPK pathway in RAW264.7 cells. Inflammation 2017, 40, 1959–1966. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, F.; Shi, J. Effect of sevoflurane treatment on microglia activation, NF-kB and MAPK activities. Immunobiology 2019, 224, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Frattaruolo, L.; Carullo, G.; Brindisi, M.; Mazzotta, S.; Bellissimo, L.; Rago, V.; Curcio, R.; Dolce, V.; Aiello, F.; Cappello, A.R. Antioxidant and anti-inflammatory activities of flavanones from Glycyrrhiza glabra L. (licorice) leaf phytocomplexes: Identification of licoflavanone as a modulator of NF-kB/MAPK pathway. Antioxidants 2019, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Kang, N.; Hu, C.; Zhang, Z.; Xu, Q.; Liu, Y.; Yang, S. Ginsenoside Rb1 exerts anti-inflammatory effects in vitro and in vivo by modulating toll-like receptor 4 dimerization and NF-kB/MAPKs signaling pathways. Phytomedicine 2020, 69, 153197. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Kang, S.; Wu, Q.; Wang, S.; Crickmore, N.; Zhou, X.; Bravo, A.; Soberón, M.; Zhang, Y. The regulation landscape of MAPK signaling cascade for thwarting Bacillus thuringiensis infection in an insect host. PLoS Pathog. 2021, 17, e1009917. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration | A260:A280 | A260:A230 | RNA Concentration (ng/µL) | CV * |
|---|---|---|---|---|---|
| Nontreatment | − | 1.888 | 1.908 | 394.05 ± 9.55 | 2.42 |
| DMSO | 0.1% | 1.966 | 1.877 | 547.81 ± 8.23 | 1.50 |
| DJ | 10 µg/mL | 1.927 | 1.839 | 453.12 ± 11.64 | 2.57 |
| DJ | 25 µg/mL | 1.997 | 1.955 | 521.39 ± 10.57 | 2.03 |
| DJ | 50 µg/mL | 1.980 | 1.925 | 537.57 ± 12.12 | 2.26 |
| DJ | 100 µg/mL | 1.912 | 1.990 | 1033.92 ± 44.85 | 4.34 |
| DJ526 | 10 µg/mL | 1.948 | 1.899 | 497.55 ± 11.60 | 2.33 |
| DJ526 | 25 µg/mL | 1.927 | 1.813 | 450.40 ± 12.55 | 2.79 |
| DJ526 | 50 µg/mL | 1.882 | 1.956 | 551.36 ± 19.13 | 3.47 |
| DJ526 | 100 µg/mL | 1.876 | 1.893 | 441.44 ± 14.03 | 3.18 |
| Aspirin | 200 µg/mL | 1.992 | 1.930 | 548.85 ± 13.80 | 2.51 |
| Gene | Nucleotide Sequence (5′−3′) | Accession Number | Target Size (bp) |
|---|---|---|---|
| COX-2 | Forward primer: 1409-AGAAGGAAATGGCTGCAGAA-1428 Reverse primer: 1602-GCTCGGCTTCCAGTATTGAG-1583 | NM_011198.5 | 194 |
| iNOS | Forward primer: 185-TTCCAGAATCCCTGGACAAG-204 Reverse primer: 364-TGGTCAAACTCTTGGGGTTC-345 | BC062378.1 | 180 |
| IL-1β | Forward primer: 531-GGGCCTCAAAGGAAAGAATC-550 Reverse primer: 713-TACCAGTTGGGGAACTCTGC-694 | NM_008361.4 | 183 |
| IL-6 | Forward primer: 33-AGTTGCCTTCTTGGGACTGA-52 Reverse primer: 223-CAGAATTGCCATTGCACAAC-204 | NM_031168.2 | 191 |
| TNF-α | Forward primer: 1-ATGAGCACAGAAAGCATGATC-21 Reverse primer: 276-TACAGGCTTGTCACTCGAATT-256 | D84199.2 | 276 |
| TLR-4 | Forward primer: 2281-CGCTCTGGCATCATCTTCAT-2300 Reverse primer: 2498-GTTGCCGTTTCTTGTTCTTCC-2478 | NM_021297.3 | 218 |
| β-actin | Forward primer: 605-CCACAGCTGAGAGGGAAATC-624 Reverse primer: 797-AAGGAAGGCTGGAAAAGAGC-778 | NM_007393.5 | 193 |
| Process | Temperature | Time | Cycle |
|---|---|---|---|
| Predenaturation | 95 °C | 10 min | 1 cycle |
| Denaturation | 95 °C | 20 s | 40 cycles |
| Annealing | 60 °C | 20 s | |
| Extension | 72 °C | 30 s | |
| Final extension | 72 °C | 5 min | 1 cycle |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monmai, C.; Kim, J.-S.; Baek, S.-H. Anti-Inflammatory Efficacy of Resveratrol-Enriched Rice Callus Extract on Lipopolysaccharide-Stimulated RAW264.7 Macrophages. Immuno 2024, 4, 131-146. https://doi.org/10.3390/immuno4020009
Monmai C, Kim J-S, Baek S-H. Anti-Inflammatory Efficacy of Resveratrol-Enriched Rice Callus Extract on Lipopolysaccharide-Stimulated RAW264.7 Macrophages. Immuno. 2024; 4(2):131-146. https://doi.org/10.3390/immuno4020009
Chicago/Turabian StyleMonmai, Chaiwat, Jin-Suk Kim, and So-Hyeon Baek. 2024. "Anti-Inflammatory Efficacy of Resveratrol-Enriched Rice Callus Extract on Lipopolysaccharide-Stimulated RAW264.7 Macrophages" Immuno 4, no. 2: 131-146. https://doi.org/10.3390/immuno4020009