
Insulinoma-Associated Protein 1 (INSM1): Diagnostic, Prognostic, and Therapeutic Use in Small Cell Lung Cancer



Abstract
:1. Introduction
2. Methodology
3. INSM1 and SCLC Oncogenesis
4. INSM1 Diagnostic Use
4.1. Performance of INSM1 as a Diagnostic Biomarker
4.2. Immunohistochemistry
4.3. Immunocytochemistry
4.4. INSM1 Gene Expression and Diagnostic Use
4.5. INSM1 Diagnostic Use—Conclusions

{kind=link}
{kind=link}
| Study | No. of SCLC Samples | Type of Sample | Preparation | INSM1 Positive Cutoff Value | INSM1 Sensitivity, % (Positive Cases/ Total Cases) | CGA/SYN/CD56 Sensitivity, % (Positive Cases/ Total Cases) | Combined Classic NE Markers Sensitivity, % (Positive Cases/ Total Cases) | INSM1 Specificity (SCLC vs. NSCLC *), % | INSM1 Expression on Other Lung NE Tumors: LCNEC/ Carcinoids (Typical and Atypical), % (Positive Cases/Total Cases) |
|---|---|---|---|---|---|---|---|---|---|
| Fujino 2015 [4] | 27 | Resections | FFPE section | Any nuclear staining in tumor cells | 100 (27/27) | 70 (19/27)/ 63 (17/27)/ n/a | n/a | 100 | n/a |
| Fujino 2017 [38] | 19 | Resections | FFPE section | Any nuclear staining in tumor cells | 100 (19/19) | 74 (14/19)/ 58 (11/19)/ 68(13/19) | n/a | 100 | 100 (4/4)/ 100 (5/5) |
| Rooper 2017 [37] | 39 | Resections (TMA) | FFPE section | Any nuclear staining in tumor cells | 95 (37/39) | 49 (19/39)/ 62 (24/39)/ 70 (21/30) | 74 (29/39) | 96 | 91 (21/23)/ 100 (48/48) ** |
| Kriegsmann 2018 [35] | 144 | Resections (TMA) | FFPE section | ≥1% stained tumor cells in at least 1 of 2 cores per patient | 86 (124/144) | 74 (107/144)/ 85 (122/144)/ 92 (132/144) | 95 (137/144) | 99 | 42 (32/77)/ 79 (120/151) |
| Doxtader 2018 [36] | 41 | Transbronchial FNA, CT-guided aspirates and a fluid cytology specimen | Cellient cell blocks | Any nuclear staining in tumor cells | 93 (38/41) | 35 (14/40)/ 93 (37/40)/ 100 (40/40) | 100 (40/40) | 100 | 100 (1/1)/ 90 (9/10) |
| Rodriguez 2018 [34] | 32 | Aspirates (31) and bronchoalveolar lavage (1) | Cell blocks | Nuclear positivity in ≥1% tumor cells | 97 (31/32) | 63 (10/16)/ 78 (14/17)/ 96 (22/23) | n/a | 100 | n/a |
| Abe 2019 *** [33] | 24 | Bronchial brushing | Cell blocks | 8.68% cutoff determined through ROC curve analysis | 98 (23/24) | Only SYN: 86 (21/24) | n/a | 100 | n/a |
| Mukhopadhyay 2019 [23] | 64 | Small biopsies (32) and whole-tissue sections of resected tumor (32) | FFPE section | Any nuclear staining in tumor cells | 98 (63/64) | 83 (53/64)/ 100(64/64)/ 95 (61/64) | 100 (64/64) | 97 | 75 (18/24)/ 98 (63/64) |
| Švajdler 2019 [31] | 112 | Endoscopic biopsies (112) | n/a | Any nuclear staining in tumor cells | 81 (81/100) | Only CD56: 84 (84/100) | n/a | n/a | n/a |
| n/a—cytology sample (13) | Cell blocks (cytoblock) | 54 (7/13) | Only CD56: 100 (13/13) | ||||||
| Nakra 2019 [32] | 60 | Aspirates (36) | Direct smears | Any nuclear staining in tumor cells | 91 (30/33) | n/a | n/a | 100 | n/a |
| Small biopsies (37) | FFPE section | 97 (36/37) | 100 (18/18)/ 96 (27/28)/ 100 (6/6) | ||||||
| Hou 2020 [29] | 11 | Aspirates | Direct smears | Nuclear positivity in ≥ 5% of tumor cells | 91 (10/11) | 73 (8/11)/ 82 (9/11)/ n/a | n/a | n/a | Only carcinoids: 90 (9/10) |
| Narayanan 2020 [6] | 19 | Resections (TMA) | FFPE section | Any nuclear staining in tumor cells | 100 (19/19) | 80 (15/19)/ 100 (19/19)/ n/a | 100 (19/19) | n/a | 100 (9/9)/ 95 (83/87) |
| Sakakibara 2020 [24] | 78 | Resections (whole slides and TMA) | FFPE section | Calculated H-score of 5 | 92 (72/78) | 48 (37/78)/ 55 (43/78)/ 81 (63/78) | 85 (66/78) | 95 | 68 (30/44)/ 95 (18/19) |
| Staaf 2020 [28] | 24 | Resections (whole slides and TMA) and biopsies | FFPE section | Any nuclear staining in tumor cells | 92 (22/24) | 67 (16/24)/ 83 (20/24)/ 96(23/24) | n/a | 98 | 87 (20/23)/ 100 (7/7) |
| At least 10% positive tumor cells | 75 (18/24) | 46 (11/24)/ 79 (19/24)/ 88 (21/24) | 88 (21/24) | 99 | 61 (14/23)/ 100 (7/7) |
5. INSM1 as a Prognostic Marker
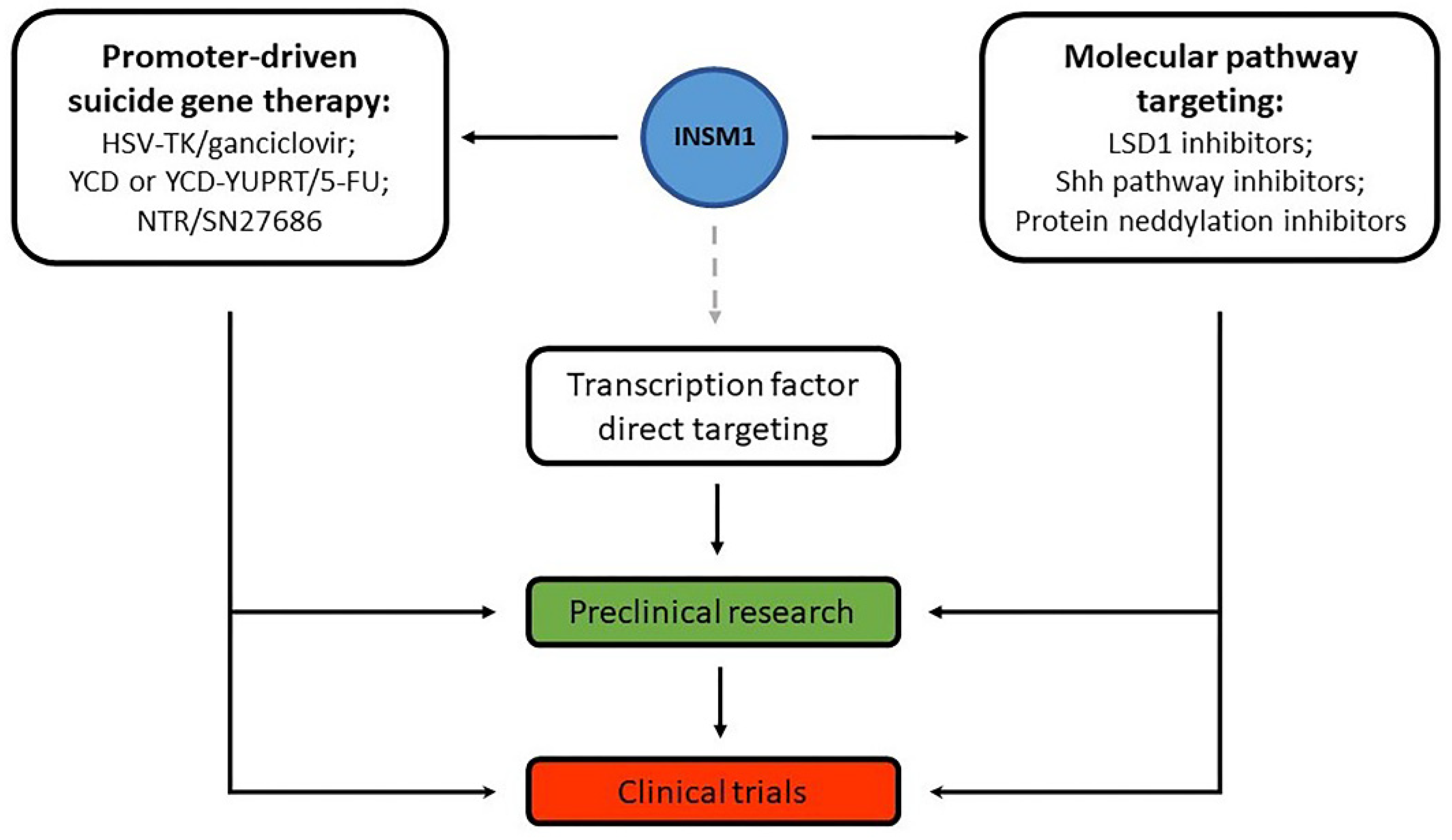
6. Gene Therapy and Other Treatment Options Targeting INSM1 or Related Molecules
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cancer.Net—Lung Cancer—Small Cell: Statistics. Available online: https://www.cancer.net/cancer-types/lung-cancer-small-cell/statistics (accessed on 18 April 2022).
- Rodriguez, E.; Lilenbaum, R.C. Small cell lung cancer: Past, present, and future. Curr. Oncol. Rep. 2010, 12, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Kris, M.G.; Benowitz, S.I.; Adams, S.; Diller, L.; Ganz, P.; Kahlenberg, M.S.; Le, Q.-T.; Markman, M.; Masters, G.A.; Newman, L.; et al. Clinical Cancer Advances 2010: Annual Report on Progress Against Cancer From the American Society of Clinical Oncology. J. Clin. Oncol. 2010, 28, 5327–5347. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K.; Motooka, Y.; Hassan, W.A.; Ali Abdalla, M.O.; Sato, Y.; Kudoh, S.; Hasegawa, K.; Niimori-Kita, K.; Kobayashi, H.; Kubota, I.; et al. Insulinoma-Associated Protein 1 Is a Crucial Regulator of Neuroendocrine Differentiation in Lung Cancer. Am. J. Pathol. 2015, 185, 3164–3177. [Google Scholar] [CrossRef] [PubMed]
- WHO Classification of Tumours Editorial Board. Thoracic Tumours, 5th ed.; International Agency for Research on Cancer Publications: Lyon, France, 2021; Volume 5. [Google Scholar]
- Narayanan, D.; Mandal, R.; Hardin, H.; Chanana, V.; Schwalbe, M.; Rosenbaum, J.; Buehler, D.; Lloyd, R.V. Long Non-coding RNAs in Pulmonary Neuroendocrine Neoplasms. Endocr. Pathol. 2020, 31, 254–263. [Google Scholar] [CrossRef]
- Dingemans, A.M.C.; Früh, M.; Ardizzoni, A.; Besse, B.; Faivre-Finn, C.; Hendriks, L.E.; Lantuejoul, S.; Peters, S.; Reguart, N.; Rudin, C.M.; et al. Small-cell lung cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up☆. Ann. Oncol. 2021, 32, 839–853. [Google Scholar] [CrossRef]
- Thunnissen, E.; Borczuk, A.C.; Flieder, D.B.; Witte, B.; Beasley, M.B.; Chung, J.H.; Dacic, S.; Lantuejoul, S.; Russell, P.A.; den Bakker, M.; et al. The Use of Immunohistochemistry Improves the Diagnosis of Small Cell Lung Cancer and Its Differential Diagnosis. An International Reproducibility Study in a Demanding Set of Cases. J. Thorac. Oncol. 2017, 12, 334–346. [Google Scholar] [CrossRef]
- Goto, Y.; De Silva, M.G.; Toscani, A.; Prabhakar, B.S.; Notkins, A.L.; Lan, M.S. A novel human insulinoma-associated cDNA, IA-1, encodes a protein with "zinc-finger" DNA-binding motifs. J. Biol. Chem. 1992, 267, 15252–15257. [Google Scholar] [CrossRef]
- Lan, M.S.; Breslin, M.B. Structure, expression, and biological function of INSM1 transcription factor in neuroendocrine differentiation. FASEB J. 2009, 23, 2024–2033. [Google Scholar] [CrossRef]
- Gierl, M.S.; Karoulias, N.; Wende, H.; Strehle, M.; Birchmeier, C. The zinc-finger factor Insm1 (IA-1) is essential for the development of pancreatic beta cells and intestinal endocrine cells. Genes Dev. 2006, 20, 2465–2478. [Google Scholar] [CrossRef]
- Jia, S.; Wildner, H.; Birchmeier, C. Insm1 controls the differentiation of pulmonary neuroendocrine cells by repressing Hes1. Dev. Biol. 2015, 408, 90–98. [Google Scholar] [CrossRef]
- Zombori, T.; Turkevi-Nagy, S.; Sejben, A.; Juhász-Nagy, G.; Cserni, G.; Furák, J.; Tiszlavicz, L.; Krenács, L.; Kővári, B. The panel of syntaxin 1 and insulinoma-associated protein 1 outperforms classic neuroendocrine markers in pulmonary neuroendocrine neoplasms. APMIS 2021, 129, 186–194. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, H.; Ramelot, T.A.; Kennedy, M.A.; Hu, R.; Yue, X.; Liu, M.; Yang, Y. Solution NMR structure of zinc finger 4 and 5 from human INSM1, an essential regulator of neuroendocrine differentiation. Proteins 2017, 85, 957–962. [Google Scholar] [CrossRef]
- Osipovich, A.B.; Long, Q.; Manduchi, E.; Gangula, R.; Hipkens, S.B.; Schneider, J.; Okubo, T.; Stoeckert, C.J., Jr.; Takada, S.; Magnuson, M.A. Insm1 promotes endocrine cell differentiation by modulating the expression of a network of genes that includes Neurog3 and Ripply3. Development 2014, 141, 2939–2949. [Google Scholar] [CrossRef]
- Breslin, M.B.; Zhu, M.; Notkins, A.L.; Lan, M.S. Neuroendocrine differentiation factor, IA-1, is a transcriptional repressor and contains a specific DNA-binding domain: Identification of consensus IA-1 binding sequence. Nucleic Acids Res. 2002, 30, 1038–1045. [Google Scholar] [CrossRef]
- Xie, J.; Cai, T.; Zhang, H.; Lan, M.S.; Notkins, A.L. The zinc-finger transcription factor INSM1 is expressed during embryo development and interacts with the Cbl-associated protein. Genomics 2002, 80, 54–61. [Google Scholar] [CrossRef]
- Lan, M.S.; Li, Q.; Lu, J.; Modi, W.S.; Notkins, A.L. Genomic organization, 5′-upstream sequence, and chromosomal localization of an insulinoma-associated intronless gene, IA-1. J. Biol. Chem. 1994, 269, 14170–14174. [Google Scholar] [CrossRef]
- Tsai, H.K.; Hornick, J.L.; Vivero, M. INSM1 expression in a subset of thoracic malignancies and small round cell tumors: Rare potential pitfalls for small cell carcinoma. Mod. Pathol. 2020, 33, 1571–1580. [Google Scholar] [CrossRef]
- Liu, W.D.; Wang, H.W.; Muguira, M.; Breslin, M.B.; Lan, M.S. INSM1 functions as a transcriptional repressor of the neuroD/beta2 gene through the recruitment of cyclin D1 and histone deacetylases. Biochem. J. 2006, 397, 169–177. [Google Scholar] [CrossRef]
- Rosenbaum, J.N.; Guo, Z.; Baus, R.M.; Werner, H.; Rehrauer, W.M.; Lloyd, R.V. INSM1: A Novel Immunohistochemical and Molecular Marker for Neuroendocrine and Neuroepithelial Neoplasms. Am. J. Clin. Pathol. 2015, 144, 579–591. [Google Scholar] [CrossRef]
- Rodriguez, E.F.; Fite, J.J.; Chowsilpa, S.; Maleki, Z. Insulinoma-associated protein 1 immunostaining on cytology specimens: An institutional experience. Hum. Pathol. 2019, 85, 128–135. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Dermawan, J.K.; Lanigan, C.P.; Farver, C.F. Insulinoma-associated protein 1 (INSM1) is a sensitive and highly specific marker of neuroendocrine differentiation in primary lung neoplasms: An immunohistochemical study of 345 cases, including 292 whole-tissue sections. Mod. Pathol. 2019, 32, 100–109. [Google Scholar] [CrossRef]
- Sakakibara, R.; Kobayashi, M.; Takahashi, N.; Inamura, K.; Ninomiya, H.; Wakejima, R.; Kitazono, S.; Yanagitani, N.; Horiike, A.; Ichinose, J.; et al. Insulinoma-associated Protein 1 (INSM1) Is a Better Marker for the Diagnosis and Prognosis Estimation of Small Cell Lung Carcinoma Than Neuroendocrine Phenotype Markers Such as Chromogranin A, Synaptophysin, and CD56. Am. J. Surg. Pathol. 2020, 44, 757–764. [Google Scholar] [CrossRef]
- Chen, C.; Breslin, M.B.; Lan, M.S. Ectopic expression of a small cell lung cancer transcription factor, INSM1 impairs alveologenesis in lung development. BMC Pulm. Med. 2016, 16, 49. [Google Scholar] [CrossRef]
- Yu, L.; Dong, Y.; Xue, J.; Xu, S.; Wang, G.; Kuang, D.; Duan, Y. SOX11 is a sensitive and specific marker for pulmonary high-grade neuroendocrine tumors. Diagn. Pathol. 2022, 17, 2. [Google Scholar] [CrossRef]
- Wang, M.; Abi-Raad, R.; Baldassarri, R.; Adeniran, A.J.; Cai, G. Expression of insulinoma-associated protein 1 in non-small cell lung cancers: A diagnostic pitfall for neuroendocrine tumors. Hum. Pathol. 2021, 115, 104–111. [Google Scholar] [CrossRef]
- Staaf, J.; Tran, L.; Söderlund, L.; Nodin, B.; Jirström, K.; Vidarsdottir, H.; Planck, M.; Mattsson, J.S.M.; Botling, J.; Micke, P.; et al. Diagnostic Value of Insulinoma-Associated Protein 1 (INSM1) and Comparison With Established Neuroendocrine Markers in Pulmonary Cancers. Arch. Pathol. Lab. Med. 2020, 144, 1075–1085. [Google Scholar] [CrossRef]
- Hou, T.; Gan, Q.; Joseph, C.T.; Sun, X.; Gong, Y. Insulinoma-associated protein 1 immunostaining for various types of neuroendocrine tumors on FNA smears. Cancer Cytopathol. 2020, 128, 725–732. [Google Scholar] [CrossRef]
- Viswanathan, K.; Siddiqui, M.T.; Borczuk, A.C. Insulinoma-associated protein 1 is a sensitive and specific marker for lung neuroendocrine tumors in cytologic and surgical specimens. J. Am. Soc. Cytopathol. 2019, 8, 299–308. [Google Scholar] [CrossRef]
- Švajdler, M.; Mezencev, R.; Šašková, B.; Ondič, O.; Mukenšnábl, P.; Michal, M. Triple marker composed of p16, CD56, and TTF1 shows higher sensitivity than INSM1 for diagnosis of pulmonary small cell carcinoma: Proposal for a rational immunohistochemical algorithm for diagnosis of small cell carcinoma in small biopsy and cytology specimens. Hum. Pathol. 2019, 85, 58–64. [Google Scholar] [CrossRef]
- Nakra, T.; Nambirajan, A.; Guleria, P.; Phulware, R.H.; Jain, D. Insulinoma-associated protein 1 is a robust nuclear immunostain for the diagnosis of small cell lung carcinoma in cytology smears. Cancer Cytopathol. 2019, 127, 539–548. [Google Scholar] [CrossRef]
- Abe, H.; Takase, Y.; Sadashima, E.; Fukumitsu, C.; Murata, K.; Ito, T.; Kawahara, A.; Naito, Y.; Akiba, J. Insulinoma-associated protein 1 is a novel diagnostic marker of small cell lung cancer in bronchial brushing and cell block cytology from pleural effusions: Validity and reliability with cutoff value. Cancer Cytopathol. 2019, 127, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.F.; Chowsilpa, S.; Maleki, Z. Insulinoma-Associated Protein 1 Immunostain: A Diagnostic Tool for Pulmonary Small Cell Carcinoma in Cytology. Acta Cytol. 2018, 62, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Kriegsmann, K.; Zgorzelski, C.; Kazdal, D.; Cremer, M.; Muley, T.; Winter, H.; Longuespée, R.; Kriegsmann, J.; Warth, A.; Kriegsmann, M. Insulinoma-associated Protein 1 (INSM1) in Thoracic Tumors is Less Sensitive but More Specific Compared With Synaptophysin, Chromogranin A, and CD56. Appl. Immunohistochem. Mol. Morphol. 2018, 28, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Doxtader, E.E.; Mukhopadhyay, S. Insulinoma-associated protein 1 is a sensitive and specific marker of neuroendocrine lung neoplasms in cytology specimens. Cancer Cytopathol. 2018, 126, 243–252. [Google Scholar] [CrossRef]
- Rooper, L.M.; Sharma, R.; Li, Q.K.; Illei, P.B.; Westra, W.H. INSM1 Demonstrates Superior Performance to the Individual and Combined Use of Synaptophysin, Chromogranin and CD56 for Diagnosing Neuroendocrine Tumors of the Thoracic Cavity. Am. J. Surg. Pathol. 2017, 41, 1561–1569. [Google Scholar] [CrossRef]
- Fujino, K.; Yasufuku, K.; Kudoh, S.; Motooka, Y.; Sato, Y.; Wakimoto, J.; Kubota, I.; Suzuki, M.; Ito, T. INSM1 is the best marker for the diagnosis of neuroendocrine tumors: Comparison with CGA, SYP and CD56. Int. J. Clin. Exp. Pathol. 2017, 10, 5393–5405. [Google Scholar]
- Lan, M.S.; Russell, E.K.; Lu, J.; Johnson, B.E.; Notkins, A.L. IA-1, a new marker for neuroendocrine differentiation in human lung cancer cell lines. Cancer Res. 1993, 53, 4169–4171. [Google Scholar]
- Amelung, J.T.; Bührens, R.; Beshay, M.; Reymond, M.A. Key genes in lung cancer translational research: A meta-analysis. Pathobiology 2010, 77, 53–63. [Google Scholar] [CrossRef]
- Tanigawa, M.; Nakayama, M.; Taira, T.; Hattori, S.; Mihara, Y.; Kondo, R.; Kusano, H.; Nakamura, K.; Abe, Y.; Ishida, Y.; et al. Insulinoma-associated protein 1 (INSM1) is a useful marker for pancreatic neuroendocrine tumor. Med. Mol. Morphol. 2018, 51, 32–40. [Google Scholar] [CrossRef]
- González, I.; Lu, H.C.; Sninsky, J.; Yang, C.; Bishnupuri, K.; Dieckgraefe, B.; Cao, D.; Chatterjee, D. Insulinoma-associated protein 1 expression in primary and metastatic neuroendocrine neoplasms of the gastrointestinal and pancreaticobiliary tracts. Histopathology 2019, 75, 568–577. [Google Scholar] [CrossRef]
- McHugh, K.E.; Mukhopadhyay, S.; Doxtader, E.E.; Lanigan, C.; Allende, D.S. INSM1 Is a Highly Specific Marker of Neuroendocrine Differentiation in Primary Neoplasms of the Gastrointestinal Tract, Appendix, and Pancreas. Am. J. Clin. Pathol. 2020, 153, 811–820. [Google Scholar] [CrossRef]
- Chen, J.F.; Yang, C.; Sun, Y.; Cao, D. Expression of novel neuroendocrine marker insulinoma-associated protein 1 (INSM1) in genitourinary high-grade neuroendocrine carcinomas: An immunohistochemical study with specificity analysis and comparison to chromogranin, synaptophysin, and CD56. Pathol.-Res. Pract. 2020, 216, 152993. [Google Scholar] [CrossRef]
- Kim, I.E., Jr.; Amin, A.; Wang, L.J.; Cheng, L.; Perrino, C.M. Insulinoma-associated Protein 1 (INSM1) Expression in Small Cell Neuroendocrine Carcinoma of the Urinary Tract. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 687–693. [Google Scholar] [CrossRef]
- Xin, Z.; Zhang, Y.; Jiang, Z.; Zhao, L.; Fan, L.; Wang, Y.; Xie, S.; Shangguan, X.; Zhu, Y.; Pan, J.; et al. Insulinoma-associated protein 1 is a novel sensitive and specific marker for small cell carcinoma of the prostate. Hum. Pathol. 2018, 79, 151–159. [Google Scholar] [CrossRef]
- Kuji, S.; Watanabe, R.; Sato, Y.; Iwata, T.; Hirashima, Y.; Takekuma, M.; Ito, I.; Abe, M.; Nagashio, R.; Omae, K.; et al. A new marker, insulinoma-associated protein 1 (INSM1), for high-grade neuroendocrine carcinoma of the uterine cervix: Analysis of 37 cases. Gynecol. Oncol. 2017, 144, 384–390. [Google Scholar] [CrossRef]
- Rooper, L.M.; Bishop, J.A.; Westra, W.H. INSM1 Is a Sensitive and Specific Marker of Neuroendocrine Differentiation in Head and Neck Tumors. Am. J. Surg. Pathol. 2018, 42, 665–671. [Google Scholar] [CrossRef]
- Yuan, C.; Jiao, F.; Zhai, C.; Zhang, J.; Wang, S.; Zhu, L. Application of INSM1 in Diagnosis and Grading of Laryngeal Neuroendocrine Carcinoma. Laryngoscope 2021, 131, E2662–E2668. [Google Scholar] [CrossRef]
- Parra, O.; Linos, K.; Yan, S.; Lilo, M.; LeBlanc, R.E. Comparative performance of insulinoma-associated protein 1 (INSM1) and routine immunohistochemical markers of neuroendocrine differentiation in the diagnosis of endocrine mucin-producing sweat gland carcinoma. J. Cutan. Pathol. 2021, 48, 41–46. [Google Scholar] [CrossRef]
- Rush, P.S.; Rosenbaum, J.N.; Roy, M.; Baus, R.M.; Bennett, D.D.; Lloyd, R.V. Insulinoma-associated 1: A novel nuclear marker in Merkel cell carcinoma (cutaneous neuroendocrine carcinoma). J. Cutan. Pathol. 2018, 45, 129–135. [Google Scholar] [CrossRef]
- Lilo, M.T.; Chen, Y.; LeBlanc, R.E. INSM1 Is More Sensitive and Interpretable than Conventional Immunohistochemical Stains Used to Diagnose Merkel Cell Carcinoma. Am. J. Surg. Pathol. 2018, 42, 1541–1548. [Google Scholar] [CrossRef]
- Maleki, Z.; Abram, M.; Dell’Aquila, M.; Kilic, I.; Lu, R.; Musarra, T.; Barkan, G.; Rajakorpi, E.; Rossi, E.D.; Kholová, I. Insulinoma-associated protein 1 (INSM-1) expression in medullary thyroid carcinoma FNA: A multi-institutional study. J. Am. Soc. Cytopathol. 2020, 9, 185–190. [Google Scholar] [CrossRef]
- Yoshida, A.; Makise, N.; Wakai, S.; Kawai, A.; Hiraoka, N. INSM1 expression and its diagnostic significance in extraskeletal myxoid chondrosarcoma. Mod. Pathol. 2018, 31, 744–752. [Google Scholar] [CrossRef]
- Wang, H.; Krishnan, C.; Charville, G.W. INSM1 Expression in Peripheral Neuroblastic Tumors and Other Embryonal Neoplasms. Pediatr. Dev. Pathol. 2019, 22, 440–448. [Google Scholar] [CrossRef]
- Farkas, L.M.; Haffner, C.; Giger, T.; Khaitovich, P.; Nowick, K.; Birchmeier, C.; Pääbo, S.; Huttner, W.B. Insulinoma-associated 1 has a panneurogenic role and promotes the generation and expansion of basal progenitors in the developing mouse neocortex. Neuron 2008, 60, 40–55. [Google Scholar] [CrossRef]
- Wildner, H.; Gierl, M.S.; Strehle, M.; Pla, P.; Birchmeier, C. Insm1 (IA-1) is a crucial component of the transcriptional network that controls differentiation of the sympatho-adrenal lineage. Development 2008, 135, 473–481. [Google Scholar] [CrossRef]
- Mahalakshmi, B.; Baskaran, R.; Shanmugavadivu, M.; Nguyen, N.T.; Velmurugan, B.K. Insulinoma-associated protein 1 (INSM1): A potential biomarker and therapeutic target for neuroendocrine tumors. Cell. Oncol. 2020, 43, 367–376. [Google Scholar] [CrossRef]
- Chen, C.; Notkins, A.L.; Lan, M.S. Insulinoma-Associated-1: From Neuroendocrine Tumor Marker to Cancer Therapeutics. Mol. Cancer Res. 2019, 17, 1597–1604. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, C.; Breslin, M.B.; Song, K.; Lan, M.S. Extra-nuclear activity of INSM1 transcription factor enhances insulin receptor signaling pathway and Nkx6.1 expression through RACK1 interaction. Cell. Signal. 2014, 26, 740–747. [Google Scholar] [CrossRef]
- Duggan, A.; Madathany, T.; de Castro, S.C.; Gerrelli, D.; Guddati, K.; García-Añoveros, J. Transient expression of the conserved zinc finger gene INSM1 in progenitors and nascent neurons throughout embryonic and adult neurogenesis. J. Comp. Neurol. 2008, 507, 1497–1520. [Google Scholar] [CrossRef]
- Pedersen, N.; Pedersen, M.W.; Lan, M.S.; Breslin, M.B.; Poulsen, H.S. The insulinoma-associated 1: A novel promoter for targeted cancer gene therapy for small-cell lung cancer. Cancer Gene Ther. 2006, 13, 375–384. [Google Scholar] [CrossRef]
- Wang, H.W.; Breslin, M.B.; Chen, C.; Akerstrom, V.; Zhong, Q.; Lan, M.S. INSM1 promoter-driven adenoviral herpes simplex virus thymidine kinase cancer gene therapy for the treatment of primitive neuroectodermal tumors. Hum. Gene Ther. 2009, 20, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Christensen, C.L.; Gjetting, T.; Poulsen, T.T.; Cramer, F.; Roth, J.A.; Poulsen, H.S. Targeted cytosine deaminase-uracil phosphoribosyl transferase suicide gene therapy induces small cell lung cancer-specific cytotoxicity and tumor growth delay. Clin. Cancer Res. 2010, 16, 2308–2319. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, S.R.; Christensen, C.L.; Sehested, M.; Cramer, F.; Poulsen, T.T.; Patterson, A.V.; Poulsen, H.S. Single agent- and combination treatment with two targeted suicide gene therapy systems is effective in chemoresistant small cell lung cancer cells. J. Gene Med. 2012, 14, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Akerstrom, V.; Chen, C.; Lan, M.S.; Breslin, M.B. Modifications to the INSM1 promoter to preserve specificity and activity for use in adenoviral gene therapy of neuroendocrine carcinomas. Cancer Gene Ther. 2012, 19, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Akerstrom, V.; Chen, C.; Lan, M.S.; Breslin, M.B. Adenoviral insulinoma-associated protein 1 promoter-driven suicide gene therapy with enhanced selectivity for treatment of neuroendocrine cancers. Ochsner J. 2013, 13, 91–99. [Google Scholar] [PubMed]
- Tseng, A.W.; Chen, C.; Breslin, M.B.; Lan, M.S. Tumor-specific promoter-driven adenoviral therapy for insulinoma. Cell. Oncol. 2016, 39, 279–286. [Google Scholar] [CrossRef]
- Welcker, J.E.; Hernandez-Miranda, L.R.; Paul, F.E.; Jia, S.; Ivanov, A.; Selbach, M.; Birchmeier, C. Insm1 controls development of pituitary endocrine cells and requires a SNAG domain for function and for recruitment of histone-modifying factors. Development 2013, 140, 4947–4958. [Google Scholar] [CrossRef]
- Chen, C.; Breslin, M.B.; Lan, M.S. INSM1 increases N-myc stability and oncogenesis via a positive-feedback loop in neuroblastoma. Oncotarget 2015, 6, 36700–36712. [Google Scholar] [CrossRef]
- Tao, W.; Zhang, Y.; Ma, L.; Deng, C.; Duan, H.; Liang, X.; Liao, R.; Lin, S.; Nie, T.; Chen, W.; et al. Haploinsufficiency of Insm1 Impairs Postnatal Baseline β-Cell Mass. Diabetes 2018, 67, 2615–2625. [Google Scholar] [CrossRef]
- Morimoto, M.; Nishinakamura, R.; Saga, Y.; Kopan, R. Different assemblies of Notch receptors coordinate the distribution of the major bronchial Clara, ciliated and neuroendocrine cells. Development 2012, 139, 4365–4373. [Google Scholar] [CrossRef]
- Peake, J.L.; Reynolds, S.D.; Stripp, B.R.; Stephens, K.E.; Pinkerton, K.E. Alteration of pulmonary neuroendocrine cells during epithelial repair of naphthalene-induced airway injury. Am. J. Pathol. 2000, 156, 279–286. [Google Scholar] [CrossRef]
- Taniwaki, M.; Daigo, Y.; Ishikawa, N.; Takano, A.; Tsunoda, T.; Yasui, W.; Inai, K.; Kohno, N.; Nakamura, Y. Gene expression profiles of small-cell lung cancers: Molecular signatures of lung cancer. Int. J. Oncol. 2006, 29, 567–575. [Google Scholar] [CrossRef]
- Rudin, C.M.; Brambilla, E.; Faivre-Finn, C.; Sage, J. Small-cell lung cancer. Nat. Rev. Dis. Primers 2021, 7, 3. [Google Scholar] [CrossRef]
- Kudoh, S.; Tenjin, Y.; Kameyama, H.; Ichimura, T.; Yamada, T.; Matsuo, A.; Kudo, N.; Sato, Y.; Ito, T. Significance of achaete-scute complex homologue 1 (ASCL1) in pulmonary neuroendocrine carcinomas; RNA sequence analyses using small cell lung cancer cells and Ascl1-induced pulmonary neuroendocrine carcinoma cells. Histochem. Cell Biol. 2020, 153, 443–456. [Google Scholar] [CrossRef]
- Chen, C.; Breslin, M.B.; Lan, M.S. Sonic hedgehog signaling pathway promotes INSM1 transcription factor in neuroendocrine lung cancer. Cell. Signal. 2018, 46, 83–91. [Google Scholar] [CrossRef]
- Pedersen, N.; Mortensen, S.; Sørensen, S.B.; Pedersen, M.W.; Rieneck, K.; Bovin, L.F.; Poulsen, H.S. Transcriptional gene expression profiling of small cell lung cancer cells. Cancer Res. 2003, 63, 1943–1953. [Google Scholar]
- Hamanaka, W.; Motoi, N.; Ishikawa, S.; Ushijima, M.; Inamura, K.; Hatano, S.; Uehara, H.; Okumura, S.; Nakagawa, K.; Nishio, M.; et al. A subset of small cell lung cancer with low neuroendocrine expression and good prognosis: A comparison study of surgical and inoperable cases with biopsy. Hum. Pathol. 2014, 45, 1045–1056. [Google Scholar] [CrossRef]
- Nicholson, S.A.; Beasley, M.B.; Brambilla, E.; Hasleton, P.S.; Colby, T.V.; Sheppard, M.N.; Falk, R.; Travis, W.D. Small cell lung carcinoma (SCLC): A clinicopathologic study of 100 cases with surgical specimens. Am. J. Surg. Pathol. 2002, 26, 1184–1197. [Google Scholar] [CrossRef]
- Švajdler, M.; Mezencev, R.; Ondič, O.; Šašková, B.; Mukenšnábl, P.; Michal, M. P16 is a useful supplemental diagnostic marker of pulmonary small cell carcinoma in small biopsies and cytology specimens. Ann. Diagn. Pathol. 2018, 33, 23–29. [Google Scholar] [CrossRef]
- Iida, Y.; Masuda, S.; Nakanishi, Y.; Shimizu, T.; Nishimaki, H.; Takahashi, M.; Hikichi, M.; Maruoka, S.; Gon, Y.; Takahashi, N.; et al. Clinicopathological characteristics of thyroid transcription factor 1-negative small cell lung cancers. Hum. Pathol. 2018, 79, 127–134. [Google Scholar] [CrossRef]
- Nandeesh, B.; Crasta, J.; Tirumalae, R. Fine-needle aspiration cytology in the diagnosis and typing of lung carcinomas. Clin. Cancer Investig. J. 2015, 4, 637–644. [Google Scholar] [CrossRef]
- Roy-Chowdhuri, S.; Aisner, D.L.; Allen, T.C.; Beasley, M.B.; Borczuk, A.; Cagle, P.T.; Capelozzi, V.; Dacic, S.; da Cunha Santos, G.; Hariri, L.P.; et al. Biomarker Testing in Lung Carcinoma Cytology Specimens: A Perspective From Members of the Pulmonary Pathology Society. Arch. Pathol. Lab. Med. 2016, 140, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Guo, M.; Sneige, N.; Gong, Y. Value of PAX8 and WT1 Immunostaining in Confirming the Ovarian Origin of Metastatic Carcinoma in Serous Effusion Specimens. Am. J. Clin. Pathol. 2012, 137, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Symmans, W.F.; Krishnamurthy, S.; Patel, S.; Sneige, N. Optimal fixation conditions for immunocytochemical analysis of estrogen receptor in cytologic specimens of breast carcinoma. Cancer 2004, 102, 34–40. [Google Scholar] [CrossRef]
- Eichhorn, F.; Dienemann, H.; Muley, T.; Warth, A.; Hoffmann, H. Predictors of survival after operation among patients with large cell neuroendocrine carcinoma of the lung. Ann. Thorac. Surg. 2015, 99, 983–989. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ogawa, H.; Uchino, K.; Ohbayashi, C.; Maniwa, Y.; Nishio, W.; Nakao, A.; Yoshimura, M. Immunohistochemical studies of pulmonary large cell neuroendocrine carcinoma: A possible association between staining patterns with neuroendocrine markers and tumor response to chemotherapy. J. Thorac. Cardiovasc. Surg. 2013, 145, 839–846. [Google Scholar] [CrossRef]
- Xu, X.; Wang, G.; Duan, Y.; Huo, Z. Prognostic value and non-neuroendocrine role of INSM1 in small cell lung cancer. Pathol.-Res. Pract. 2021, 229, 153693. [Google Scholar] [CrossRef]
- Minami, K.; Jimbo, N.; Tanaka, Y.; Ogawa, H.; Hokka, D.; Nishio, W.; Yoshimura, M.; Itoh, T.; Maniwa, Y. Insulinoma-associated protein 1 is a prognostic biomarker in pulmonary high-grade neuroendocrine carcinoma. J. Surg. Oncol. 2020, 122, 243–253. [Google Scholar] [CrossRef]
- Baine, M.K.; Hsieh, M.S.; Lai, W.V.; Egger, J.V.; Jungbluth, A.A.; Daneshbod, Y.; Beras, A.; Spencer, R.; Lopardo, J.; Bodd, F.; et al. SCLC Subtypes Defined by ASCL1, NEUROD1, POU2F3, and YAP1: A Comprehensive Immunohistochemical and Histopathologic Characterization. J. Thorac. Oncol. 2020, 15, 1823–1835. [Google Scholar] [CrossRef]
- McColl, K.; Wildey, G.; Sakre, N.; Lipka, M.B.; Behtaj, M.; Kresak, A.; Chen, Y.; Yang, M.; Velcheti, V.; Fu, P.; et al. Reciprocal expression of INSM1 and YAP1 defines subgroups in small cell lung cancer. Oncotarget 2017, 8, 73745–73756. [Google Scholar] [CrossRef]
- Byers, L.A.; Wang, J.; Nilsson, M.B.; Fujimoto, J.; Saintigny, P.; Yordy, J.; Giri, U.; Peyton, M.; Fan, Y.H.; Diao, L.; et al. Proteomic profiling identifies dysregulated pathways in small cell lung cancer and novel therapeutic targets including PARP1. Cancer Discov. 2012, 2, 798–811. [Google Scholar] [CrossRef]
- Rudin, C.M.; Poirier, J.T.; Byers, L.A.; Dive, C.; Dowlati, A.; George, J.; Heymach, J.V.; Johnson, J.E.; Lehman, J.M.; MacPherson, D.; et al. Molecular subtypes of small cell lung cancer: A synthesis of human and mouse model data. Nat. Rev. Cancer 2019, 19, 289–297. [Google Scholar] [CrossRef]
- Lee, Y.; Joo, J.; Lee, Y.J.; Lee, E.K.; Park, S.; Kim, T.S.; Lee, S.H.; Kim, S.Y.; Wie, G.A.; Park, M.; et al. Randomized phase II study of platinum-based chemotherapy plus controlled diet with or without metformin in patients with advanced non-small cell lung cancer. Lung Cancer 2021, 151, 8–15. [Google Scholar] [CrossRef]
- Fan, H.; Bai, S.; Guan, X.; Ma, W.; Fu, Y.; Zhang, X.; Deng, L.; Tian, J. Metformin improves survival in patients with concurrent diabetes and small cell lung cancer: A meta-analysis. Minerva Endocrinol. 2021, in press. [Google Scholar] [CrossRef]
- Chun, S.G.; Liao, Z.; Jeter, M.D.; Chang, J.Y.; Lin, S.H.; Komaki, R.U.; Guerrero, T.M.; Mayo, R.C.; Korah, B.M.; Koshy, S.M.; et al. Metabolic Responses to Metformin in Inoperable Early-stage Non-Small Cell Lung Cancer Treated With Stereotactic Radiotherapy: Results of a Randomized Phase II Clinical Trial. Am. J. Clin. Oncol. 2020, 43, 231–235. [Google Scholar] [CrossRef]
- Zeng, S.; Gan, H.X.; Xu, J.X.; Liu, J.Y. Metformin improves survival in lung cancer patients with type 2 diabetes mellitus: A meta-analysis. Med. Clin. 2019, 152, 291–297. [Google Scholar] [CrossRef]
- Lu, H.; Xie, F.; Huang, Z.; Qin, J.; Han, N.; Mao, W. Effect of metformin in the prognosis of patients with small-cell lung cancer combined with diabetes mellitus. Adv. Clin. Exp. Med. 2018, 27, 1195–1199. [Google Scholar] [CrossRef]
- Owonikoko, T.K.; Park, K.; Govindan, R.; Ready, N.; Reck, M.; Peters, S.; Dakhil, S.R.; Navarro, A.; Rodríguez-Cid, J.; Schenker, M.; et al. Nivolumab and Ipilimumab as Maintenance Therapy in Extensive-Disease Small-Cell Lung Cancer: CheckMate 451. J. Clin. Oncol. 2021, 39, 1349–1359. [Google Scholar] [CrossRef]
- Iams, W.T.; Porter, J.; Horn, L. Immunotherapeutic approaches for small-cell lung cancer. Nat. Rev. Clin. Oncol. 2020, 17, 300–312. [Google Scholar] [CrossRef]
- Takagi, S.; Ishikawa, Y.; Mizutani, A.; Iwasaki, S.; Matsumoto, S.; Kamada, Y.; Nomura, T.; Nakamura, K. LSD1 Inhibitor T-3775440 Inhibits SCLC Cell Proliferation by Disrupting LSD1 Interactions with SNAG Domain Proteins INSM1 and GFI1B. Cancer Res. 2017, 77, 4652–4662. [Google Scholar] [CrossRef]
- Norton, J.P.; Augert, A.; Eastwood, E.; Basom, R.; Rudin, C.M.; MacPherson, D. Protein neddylation as a therapeutic target in pulmonary and extrapulmonary small cell carcinomas. Genes Dev. 2021, 35, 870–887. [Google Scholar] [CrossRef]
- Chen, C.; Breslin, M.B.; Guidry, J.J.; Lan, M.S. 5′-Iodotubercidin represses insulinoma-associated-1 expression, decreases cAMP levels, and suppresses human neuroblastoma cell growth. J. Biol. Chem. 2019, 294, 5456–5465. [Google Scholar] [CrossRef]
- Breslin, M.B.; Zhu, M.; Lan, M.S. NeuroD1/E47 regulates the E-box element of a novel zinc finger transcription factor, IA-1, in developing nervous system. J. Biol. Chem. 2003, 278, 38991–38997. [Google Scholar] [CrossRef]
- Tseng, A.W.; Akerstrom, V.; Chen, C.; Breslin, M.B.; Lan, M.S. Detection of neuroendocrine tumors using promoter-specific secreted Gaussia luciferase. Int. J. Oncol. 2016, 48, 173–180. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, R.; Henrique, R. Insulinoma-Associated Protein 1 (INSM1): Diagnostic, Prognostic, and Therapeutic Use in Small Cell Lung Cancer. J. Mol. Pathol. 2022, 3, 140-167. https://doi.org/10.3390/jmp3030013
Rocha R, Henrique R. Insulinoma-Associated Protein 1 (INSM1): Diagnostic, Prognostic, and Therapeutic Use in Small Cell Lung Cancer. Journal of Molecular Pathology. 2022; 3(3):140-167. https://doi.org/10.3390/jmp3030013
Chicago/Turabian StyleRocha, Renato, and Rui Henrique. 2022. "Insulinoma-Associated Protein 1 (INSM1): Diagnostic, Prognostic, and Therapeutic Use in Small Cell Lung Cancer" Journal of Molecular Pathology 3, no. 3: 140-167. https://doi.org/10.3390/jmp3030013