
Defensive Mutualism of Endophytic Fungi: Effects of Sphaeropsidin A against a Model Lepidopteran Pest †

 , , , , , ,
, , , , , ,  , ,
, ,  ,
,  and
and 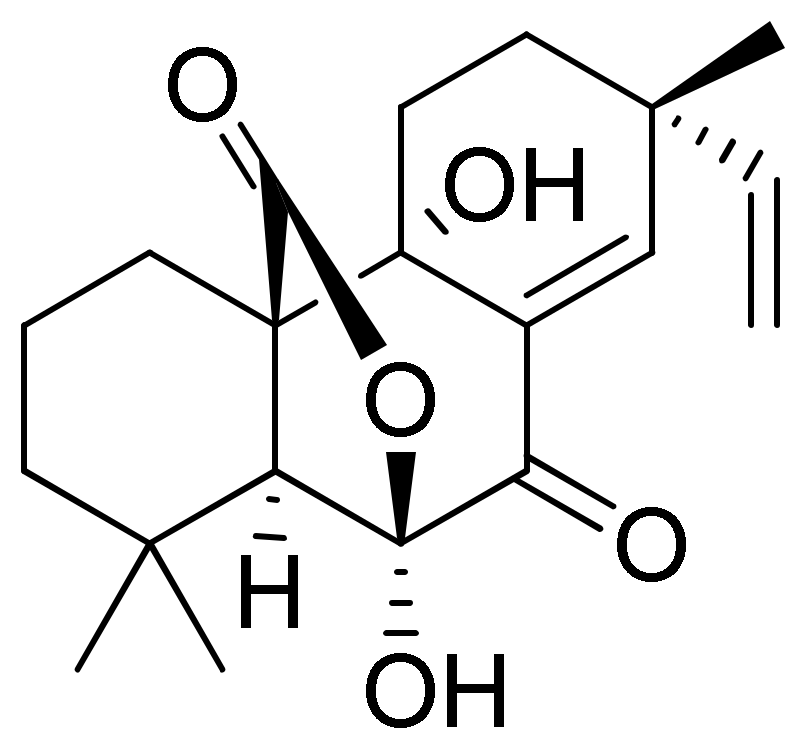{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Culturing
2.2. Isolation of SphA from Crude Extract
2.3. General Experimental Procedures
2.4. Bioassays on Spodoptera littoralis
2.4.1. Topical Application
2.4.2. Oral Administration
2.5. Statistical Analysis
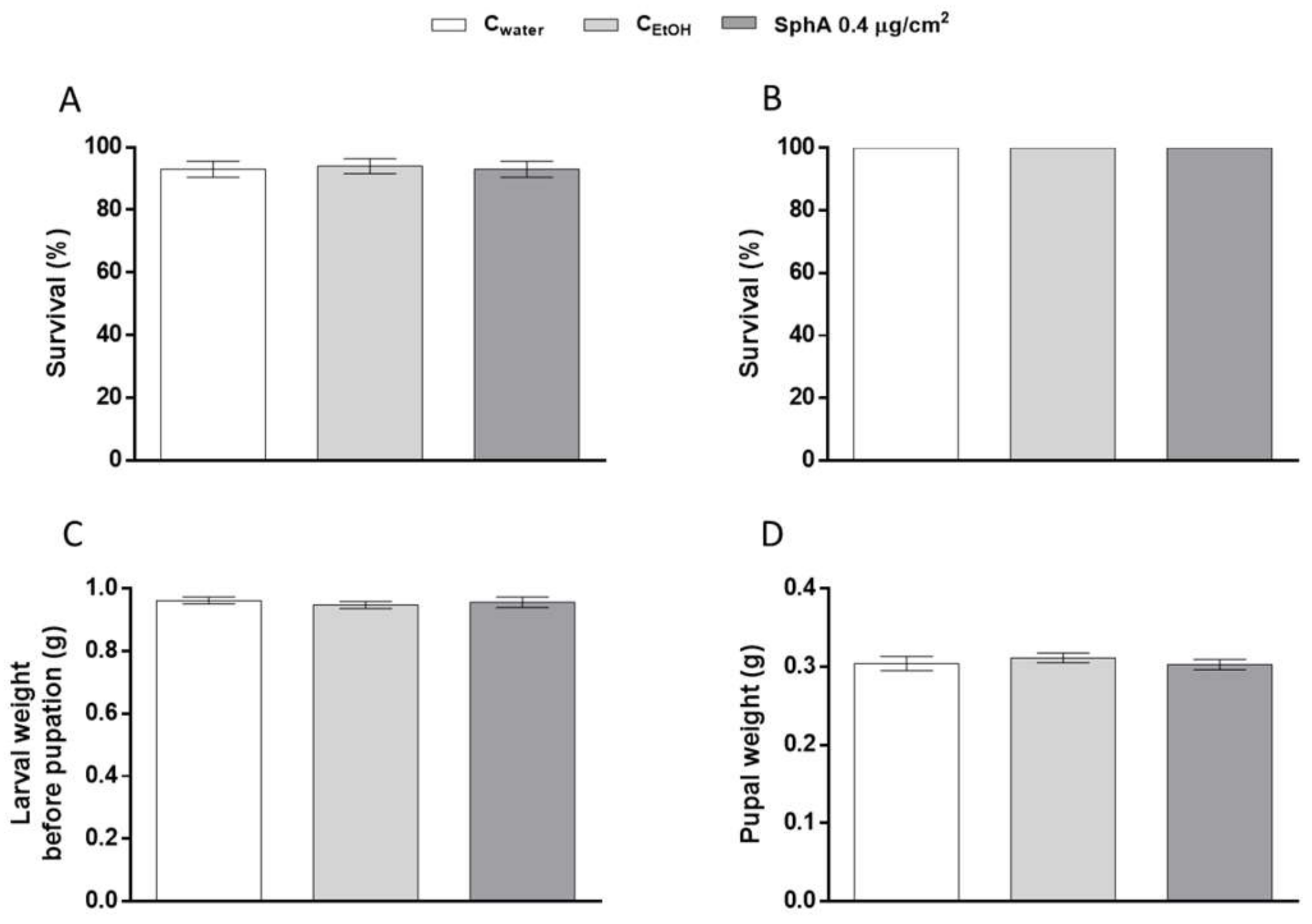
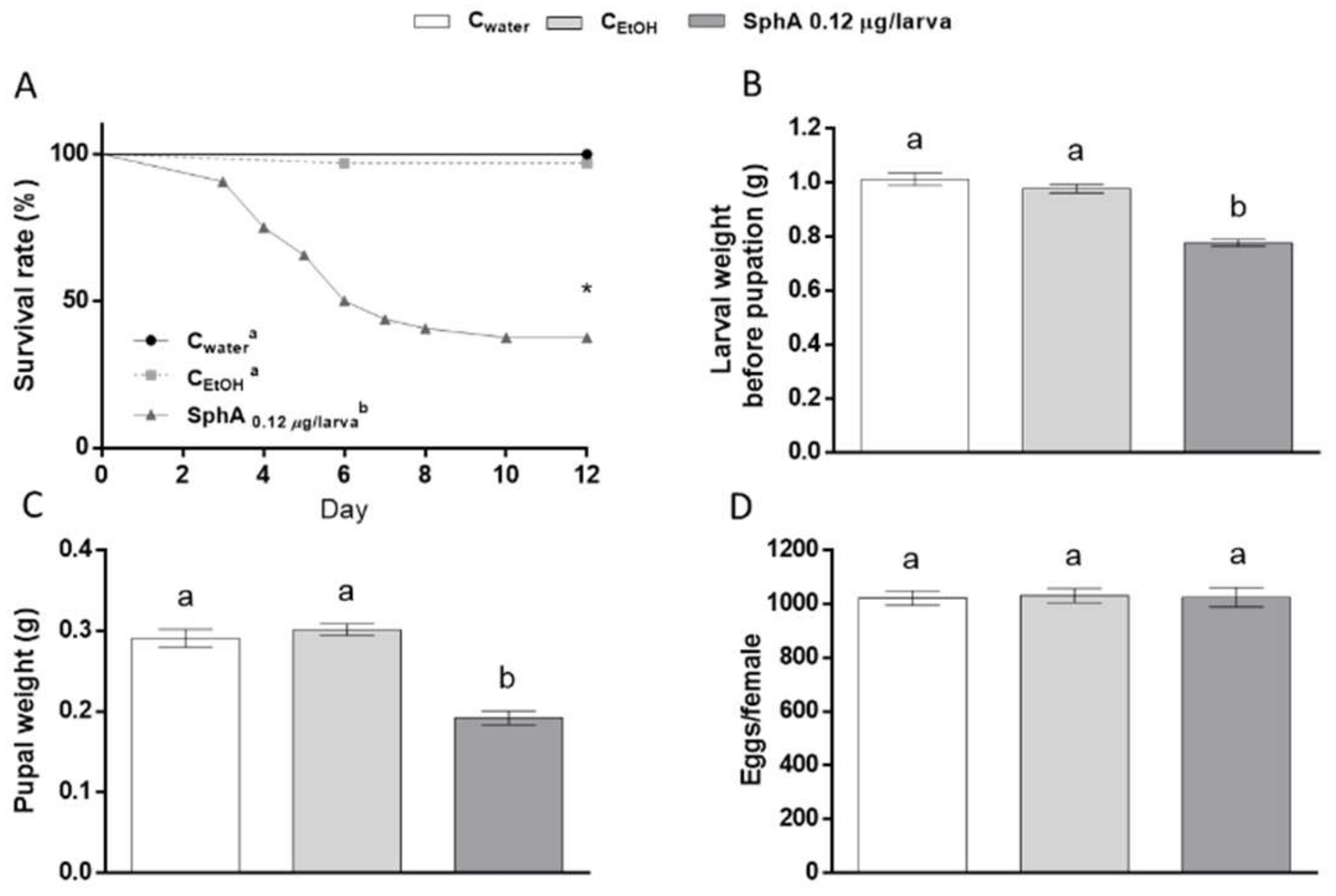
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, H.; Wei, T.; Lou, H.; Shu, X.; Chen, Q. A Critical Review on Communication Mechanism within Plant-Endophytic Fungi Interactions to Cope with Biotic and Abiotic Stresses. J. Fungi 2021, 7, 719. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Song, Y.C.; Tan, R.X. Biology and chemistry of endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. N. Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fung. Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Redman, R.S.; Dunigan, D.D.; Rodriguez, R.J. Fungal symbiosis: From mutualism to parasitism, who controls the outcome, host or invader? N. Phytol. 2001, 151, 705–716. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Salvatore, M.M.; Andolfi, A.; Nicoletti, R. The thin line between pathogenicity and endophytism: The case of Lasiodiplodia theobromae. Agriculture 2020, 10, 488. [Google Scholar] [CrossRef]
- Berestetskiy, A.; Hu, Q. The Chemical Ecology Approach to Reveal Fungal Metabolites for Arthropod Pest Management. Microorganisms 2021, 9, 1379. [Google Scholar] [CrossRef]
- Nicoletti, R.; Fiorentino, A. Plant bioactive metabolites and drugs produced by endophytic fungi of Spermatophyta. Agriculture 2015, 5, 918–970. [Google Scholar] [CrossRef] [Green Version]
- Popay, A.J.; Rowan, D.D. Endophytic fungi as mediators of plant-insect interactions. In Insect-Plant Interactions, 5th ed.; Bernays, E.A., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 83–104. [Google Scholar] [CrossRef]
- Collemare, J.; Navarro-Munoz, J.C.; Mosunova, O. The Biosynthesis of Fungal Secondary Metabolites: From Fundamentals to Biotechnological Applications. In Encyclopedia of Mycology, 1st ed.; Zaragoza, O., Casadevall, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 458–476. [Google Scholar]
- Demain, A.L.; Fang, A. The Natural Functions of Secondary Metabolites. In History of Modern Biotechnology I, 1st ed.; Fiechter, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 1–39. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Scanu, B.; Evidente, A.; Cimmino, A. Bioactive metabolites from pathogenic and endophytic fungi of forest trees. Curr. Med. Chem. 2018, 25, 208–252. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef]
- Marrone, P.G. Pesticidal natural products-status and future potential. Pest Manag. Sci. 2019, 75, 2325–2340. [Google Scholar] [CrossRef]
- Andolfi, A.; Maddau, L.; Basso, S.; Linaldeddu, B.T.; Cimmino, A.; Scanu, B.; Deidda, A.; Tuzi, A.; Evidente, A.A. Diplopimarane, a 20-nor-ent-pimarane produced by the oak pathogen Diplodia quercivora. J. Nat. Prod. 2014, 77, 2352–2360. [Google Scholar] [CrossRef]
- Sparapano, L.; Bruno, G.; Fierro, O.; Evidente, A. Studies on structure–activity relationship of sphaeropsidins A–F, phytotoxins produced by Sphaeropsis sapinea f. sp. cupressi. Phytochemistry 2004, 65, 189–198. [Google Scholar] [CrossRef]
- Ellestad, G.A.; Kunstmann, M.P.; Mirando, P.; Morton, G.O. Structures of fungal diterpene antibiotics LL-S491. beta. and-. gamma. J. Am. Chem. Soc. 1972, 94, 6206–6208. [Google Scholar] [CrossRef]
- Morooka, N.; Tatsuno, T.; Tsunoda, H.; Kobayashi, K.; Sakurai, T. Chemical and Toxicological Studies of the Phytotoxin, 6α,7β,9α-Trihydroxy-8,15-isopimaradiene-20,6-γ-lactone, Produced by a Parasitic Fungus, Phomopsis sp., in Wilting Pine Trees. Agric. Biol. Chem. 1986, 50, 2003–2007. [Google Scholar]
- Weber, R.W.; Kappe, R.; Paululat, T.; Mösker, E.; Anke, H. Anti-Candida metabolites from endophytic fungi. Phytochemistry 2007, 68, 886–892. [Google Scholar] [CrossRef]
- Wang, X.N.; Bashyal, B.P.; Wijeratne, E.K.; U’Ren, J.M.; Liu, M.X.; Gunatilaka, M.K.; Arnold, A.E.; Gunatilaka, A.L. Smardaesidins A–G, Isopimarane and 20-nor-Isopimarane Diterpenoids from Smardaea sp., a Fungal Endophyte of the Moss Ceratodon purpureus. J. Nat. Prod. 2011, 74, 2052–2061. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Guo, Z.K.; Jiang, R.; Wei, W.; Wang, T.; Guo, Y.; Song, Y.C.; Jiao, R.H.; Tan, R.X.; Ge, H.M. New flavonol and diterpenoids from the endophytic fungus Aspergillus sp. YXf3. Planta Med. 2013, 79, 348–352. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Scott, R.; Hooper, A.R.; Bartholomeusz, G.A.; Kornienko, A.; Bills, G.F. Aspergillus candidus is a newly recognized source of sphaeropsidin A: Isolation, semi-synthetic derivatization and anticancer evaluation. Bioorg. Med. Chem. Lett. 2017, 27, 5436–5440. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, G.F.; Adpressa, D.A.; Bruhn, T.; Loesgen, S. Polyketides from marine-derived Aspergillus porosus: Challenges and opportunities for determining absolute configuration. J. Nat. Prod. 2019, 82, 2780–2789. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, X.Y.; Zhang, P.L.; Gao, H.M.; Zhang, L.T.; Shen, T.; Ren, D.M.; Lou, H.X.; Wang, X.N. New terpenoids and triketides from culture of the fungus Botrysphaeria laricina. Fitoterapia 2020, 147, 104758. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Andolfi, A.; Avolio, F.; Ali, A.; Tabanca, N.; Khan, I.A.; Evidente, A. Cyclopaldic Acid, Seiridin, and Sphaeropsidin A as Fungal Phytotoxins, and Larvicidal and Biting Deterrents against Aedes aegypti (Diptera: Culicidae): Structure-Activity Relationships. Chem. Biodivers. 2013, 10, 1239–1251. [Google Scholar] [CrossRef]
- Mahamedi, A.E.; Phillips, A.J.; Lopes, A.; Djellid, Y.; Arkam, M.; Eichmeier, A.; Zitouni, A.; Alves, A.; Berraf-Tebbal, A. Diversity, distribution and host association of Botryosphaeriaceae species causing oak decline across different forest ecosystems in Algeria. Eur. J. Plant Pathol. 2020, 158, 745–765. [Google Scholar] [CrossRef]
- Salvatore, M.M.; DellaGreca, M.; Nicoletti, R.; Salvatore, F.; Tuzi, A.; De Tommaso, G.; Alves, A.; Mahamedi, A.E.; Berraf-Tebbal, A.; Andolfi, A. Mitidjospirone, a new spirodioxynaphthalene and GC-MS screening of secondary metabolites produced by strains of Lasiodiplodia mitidjana associated to Citrus sinensis dieback. Nat. Prod. Res. 2021, 1–10. [Google Scholar] [CrossRef]
- Di Lelio, I.; Varricchio, P.; Di Prisco, G.; Marinelli, A.; Lasco, V.; Caccia, S.; Casartelli, M.; Giordana, B.; Rao, R.; Gigliotti, S.; et al. Functional analysis of an immune gene of Spodoptera littoralis by RNAi. J. Insect Physiol. 2014, 64, 90–97. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Lelio, I.; Salvatore, M.M.; Della Greca, M.; Mahamedi, A.E.; Alves, A.; Berraf-Tebbal, A.; Volpe, G.; Russo, E.; Becchimanzi, A.; Nicoletti, R.; et al. Defensive Mutualism of Endophytic Fungi: Effects of Sphaeropsidin A against a Model Lepidopteran Pest. Chem. Proc. 2022, 10, 42. https://doi.org/10.3390/IOCAG2022-12216
Di Lelio I, Salvatore MM, Della Greca M, Mahamedi AE, Alves A, Berraf-Tebbal A, Volpe G, Russo E, Becchimanzi A, Nicoletti R, et al. Defensive Mutualism of Endophytic Fungi: Effects of Sphaeropsidin A against a Model Lepidopteran Pest. Chemistry Proceedings. 2022; 10(1):42. https://doi.org/10.3390/IOCAG2022-12216
Chicago/Turabian StyleDi Lelio, Ilaria, Maria Michela Salvatore, Marina Della Greca, Alla Eddine Mahamedi, Artur Alves, Akila Berraf-Tebbal, Gennaro Volpe, Elia Russo, Andrea Becchimanzi, Rosario Nicoletti, and et al. 2022. "Defensive Mutualism of Endophytic Fungi: Effects of Sphaeropsidin A against a Model Lepidopteran Pest" Chemistry Proceedings 10, no. 1: 42. https://doi.org/10.3390/IOCAG2022-12216