
Artificial Light at Night (ALAN) Influences Understory Plant Traits through Ecological Processes: A Two-Year Experiment in a Rubber Plantation in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Species Selection
2.3. Measurements
2.4. Data Analysis
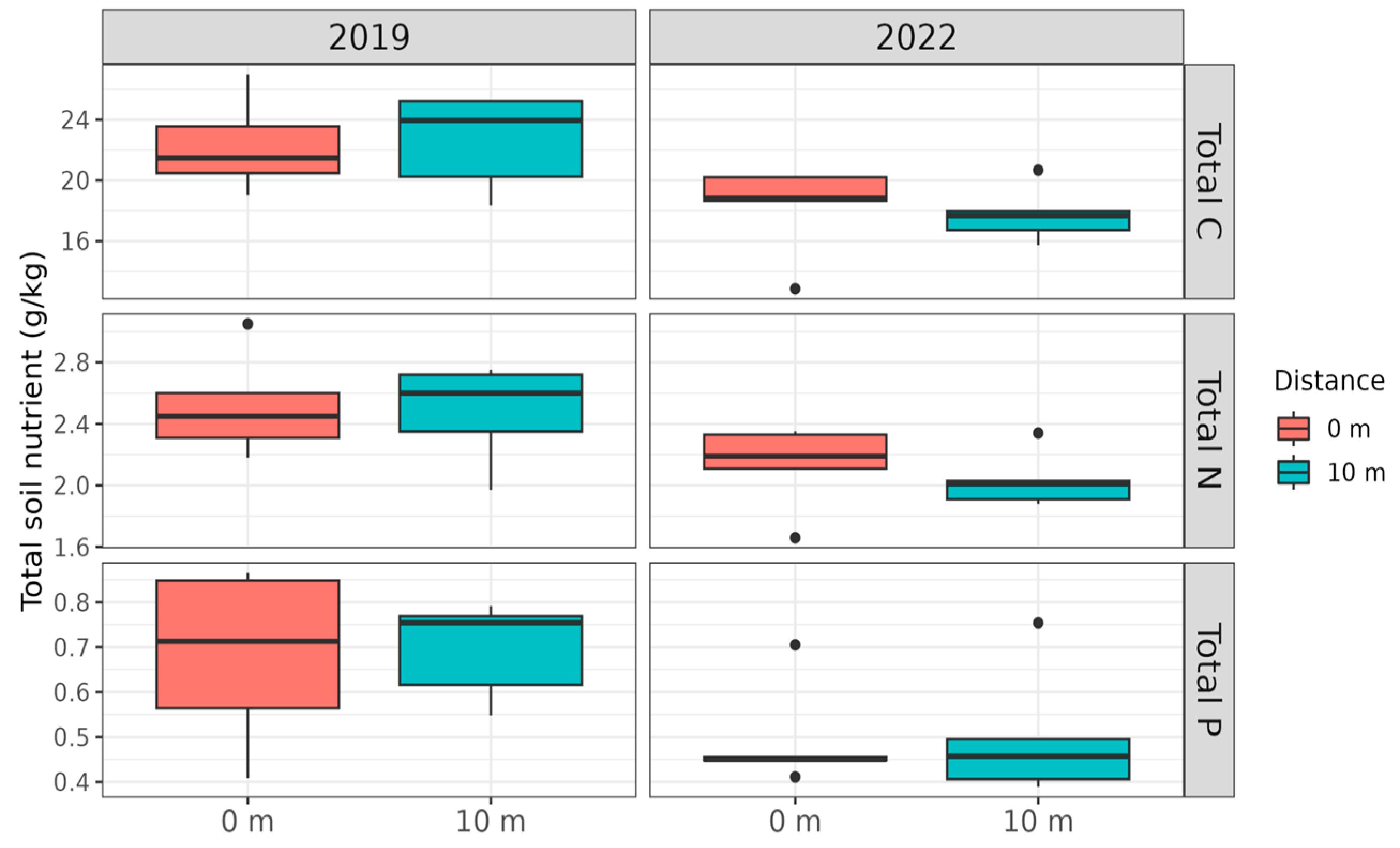
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennie, J.; Davies, T.W.; Cruse, D.; Gaston, K.J. Ecological effects of artificial light at night on wild plants. J. Ecol. 2016, 104, 611–620. [Google Scholar] [CrossRef]
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. The ecological impacts of nighttime light pollution: a mechanistic appraisal. Biol. Rev. 2013, 88, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Longcore, T.; Rich, C. Ecological light pollution. Front. Ecol. Environ. 2004, 2, 191–198. [Google Scholar] [CrossRef]
- Falchi, F.; Cinzano, P.; Duriscoe, D.; Kyba, C.C.M.; Elvidge, C.D.; Baugh, K.; Portnov, B.A.; Rybnikova, N.A.; Furgoni, R. The new world atlas of artificial night sky brightness. Sci. Adv. 2016, 2, e1600377. [Google Scholar] [CrossRef]
- Falcón, J.; Torriglia, A.; Attia, D.; Viénot, F.; Gronfier, C.; Behar-Cohen, F.; Martinsons, C.; Hicks, D. Exposure to Artificial Light at Night and the Consequences for Flora, Fauna, and Ecosystems. Front. Neurosci. 2020, 141, 1183. [Google Scholar] [CrossRef] [PubMed]
- Forsburg, Z.R.; Guzman, A.; Gabor, C.R. Artificial light at night (ALAN) affects the stress physiology but not the behavior or growth of Rana berlandieri and Bufo valliceps. Environ. Pollut. 2021, 277, 116775. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Loss, S.S.; Marra, P.P. Bird collisions in the United States: Estimates of annual mortality and species vulnerability. Condor 2014, 116, 8–23. [Google Scholar] [CrossRef]
- Rich, C.; Longcore, T. Ecological Consequences of Artificial Night Lighting; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Boyes, D.H.; Evans, D.M.; Fox, R.; Parsons, M.S.; Pocock, M.J.O. Street lighting has detrimental impacts on local insect populations. Sci. Adv. 2021, 7, eabi8322. [Google Scholar] [CrossRef]
- Owens, A.C.S.; Cochard, P.; Durrant, J.; Farnworth, B.; Perkin, E.K.; Seymoure, B. Light pollution is a driver of insect declines. Biol. Conserv. 2020, 241, 108259. [Google Scholar] [CrossRef]
- Yang, R.; Nakamura, A. Cavity-dwelling ants tend to colonize close to artificial light. Biodivers. Sci. 2022, 30, 22067. [Google Scholar] [CrossRef]
- Liu, Y.; Speißer, B.; Knop, E.; Van Kleunen, M. The Matthew effect: Common species become more common and rare ones become more rare in response to artificial light at night. Glob. Chang. Biol. 2022, 28, 3674–3682. [Google Scholar] [CrossRef]
- Speißer, B.; Liu, Y.; van Kleunen, M. Biomass responses of widely and less-widely naturalized alien plants to artificial light at night. J. Ecol. 2021, 109, 1819–1827. [Google Scholar] [CrossRef]
- Onoda, Y.; Wright, I.; Evans, J.; Hikosaka, K.; Kitajima, K.; Niinemets, Ü.; Poorter, H.; Tosens, T.; Westoby, M. Physiological and structural tradeoffs underlying the leaf economics spectrum. New Phytol. 2017, 214, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Osnas, J.L.D.; Katabuchi, M.; Kitajima, K.; Wright, S.J.; Reich, P.B.; Van Bael, S.A.; Kraft, N.J.B.; Samaniego, M.J.; Pacala, S.W.; Lichstein, J.W. Divergent drivers of leaf trait variation within species, among species, and among functional groups. Proc. Natl. Acad. Sci. USA 2018, 115, 5480–5485. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Tupayachi, R.; Emerson, R.; Martinez, P.; Sinca, F.; Powell, G.V.N.; Wright, S.J.; Lugo, A.E. Taxonomy and remote sensing of leaf mass per area (LMA) in humid tropical forests. Ecol. Appl. 2011, 21, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Paik, I.; Huq, E. Plant photoreceptors: Multi-functional sensory proteins and their signaling networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Terashima, I.; Hanba, Y.T.; Tazoe, Y.; Vyas, P.; Yano, S. Irradiance and phenotype: Comparative eco-development of sun and shade leaves in relation to photosynthetic CO2 diffusion. J. Exp. Bot. 2006, 57, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Ackerly, D.; Knight, C.; Weiss, S.; Barton, K.; Starmer, K. Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: Contrasting patterns in species level and community level analyses. Oecologia 2002, 130, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Kull, O.; Tenhunen, J.D. Within-canopy variation in the rate of development of photosynthetic capacity is proportional to integrated quantum flux density in temperate deciduous trees. Plant Cell Environ. 2004, 27, 293–313. [Google Scholar] [CrossRef]
- Onoda, Y.; Schieving, F.; Anten, N.P. Effects of Light and Nutrient Availability on Leaf Mechanical Properties of Plantago major: A Conceptual Approach. Ann. Bot. 2008, 101, 727–736. [Google Scholar] [CrossRef]
- Baker, R.R.; Sadovy, Y. The distance and nature of the light-trap response of moths. Nature 1978, 276, 818–821. [Google Scholar] [CrossRef]
- Sotthibandhu, S.; Baker, R. Celestial orientation by the large yellow underwing moth, Noctua pronuba L. Anim. Behav. 1979, 27, 786–800. [Google Scholar] [CrossRef]
- Minnaar, C.; Boyles, J.G.; Minnaar, I.A.; Sole, C.L.; McKechnie, A.E. Stacking the odds: Light pollution may shift the balance in an ancient predator-prey arms race. J. Appl. Ecol. 2015, 52, 522–531. [Google Scholar] [CrossRef]
- Owens, A.C.S.; Lewis, S.M. The impact of artificial light at night on nocturnal insects: A review and synthesis. Ecol. Evol. 2018, 8, 11337–11358. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.J. Plant responses to nutrient addition experiments conducted in tropical forests. Ecol. Monogr. 2019, 89, e01382. [Google Scholar] [CrossRef]
- Behie, S.W.; Bidochka, M.J. Insects as a Nitrogen Source for Plants. Insects 2013, 4, 413–424. [Google Scholar] [CrossRef] [PubMed]
- De la Riva, E.G.; Olmo, M.; Poorter, H.; Ubera, J.L.; Villar, R. Leaf Mass per Area (LMA) and Its Relationship with Leaf Structure and Anatomy in 34 Mediterranean Woody Species along a Water Availability Gradient. PLoS ONE 2016, 11, e0148788. [Google Scholar] [CrossRef] [PubMed]
- Katabuchi, M.; Kurokawa, H.; Davies, S.J.; Tan, S.; Nakashizuka, T. Soil resource availability shapes community trait structure in a species-rich dipterocarp forest. J. Ecol. 2012, 100, 643–651. [Google Scholar] [CrossRef]
- Hernández-Vargas, G.; Sánchez-Velásquez, L.R.; López-Acosta, J.C.; Noa-Carrazana, J.C.; Perroni, Y. Relationship between soil properties and leaf functional traits in early secondary succession of tropical montane cloud forest. Ecol. Res. 2019, 34, 213–224. [Google Scholar] [CrossRef]
- Warren-Thomas, E.M.; Edwards, D.P.; Bebber, D.P.; Chhang, P.; Diment, A.N.; Evans, T.D.; Lambrick, F.H.; Maxwell, J.F.; Nut, M.; O’kelly, H.J.; et al. Protecting tropical forests from the rapid expansion of rubber using carbon payments. Nat. Commun. 2018, 9, 911. [Google Scholar] [CrossRef]
- Sadeghi, S.M.M.; Van Stan, J.T.; Pypker, T.G.; Friesen, J. Canopy hydrometeorological dynamics across a chronosequence of a globally invasive species, Ailanthus altissima (Mill., tree of heaven). Agric. For. Meteorol. 2017, 240–241, 10–17. [Google Scholar] [CrossRef]
- Sadeghi, S.M.M.; Van Stan, J.T.; Pypker, T.G.; Tamjidi, J.; Friesen, J.; Farahnaklangroudi, M. Importance of transitional leaf states in canopy rainfall partitioning dynamics. Eur. J. For. Res. 2018, 137, 121–130. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, X.; Chang, X.; Tie, L. Impacts of canopy structure on the sub-canopy solar radiation under a deciduous forest based on fisheye photographs. Res. Cold Arid. Reg. 2023, 15, 150–160. [Google Scholar] [CrossRef]
- Katabuchi, M. LeafArea: An R package for rapid digital image analysis of leaf area. Ecol. Res. 2015, 30, 1073–1077. [Google Scholar] [CrossRef]
- Maenpuen, P.; Katabuchi, M.; Onoda, Y.; Zhou, C.; Zhang, J.; Chen, Y. Sources and consequences of mismatch between leaf disc and whole-leaf leaf mass per area (LMA). Am. J. Bot. 2022, 109, 1242–1250. [Google Scholar] [CrossRef]
- Carpenter, B.; Gelman, A.; Hoffman, M.D.; Lee, D.; Goodrich, B.; Betancourt, M.; Brubaker, M.; Guo, J.; Li, P.; Riddell, A. Stan: A Probabilistic Programming Language. J. Stat. Soft. 2017, 76, 1. [Google Scholar] [CrossRef]
- Gelman, A.; Jakulin, A.; Pittau, M.G.; Su, Y.-S. A weakly informative default prior distribution for logistic and other regression models. Ann. Appl. Stat. 2008, 2, 1360–1383. [Google Scholar] [CrossRef]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Vehtari, A.; Gelman, A.; Simpson, D.; Carpenter, B.; Bürkner, P.-C. Rank-Normalization, Folding, and Localization: An Improved R^ for Assessing Convergence of MCMC (with Discussion). Bayesian Anal. 2021, 16, 667–718. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Landau, W.M. The targets R package: A dynamic Make-like function-oriented pipeline toolkit for reproducibility and high-performance computing. J. Open Source Softw. 2021, 6, 2959. [Google Scholar] [CrossRef]
- Yang, J.-C.; Huang, J.-H.; Pan, Q.-M.; Tang, J.-W.; Han, X.-G. Long-term impacts of land-use change on dynamics of tropical soil carbon and nitrogen pools. J. Environ. Sci. 2004, 16, 256–261. [Google Scholar]
- Fagan, W.F.; Siemann, E.; Mitter, C.; Denno, R.F.; Huberty, A.F.; Woods, H.A.; Elser, J.J. Nitrogen in Insects: Implications for Trophic Complexity and Species Diversification. Am. Nat. 2002, 160, 784–802. [Google Scholar] [CrossRef]
- Schulze, W.; Schulze, E.D.; Pate, J.S.; Gillison, A.N. The nitrogen supply from soils and insects during growth of the pitcher plants Nepenthes mirabilis, Cephalotus follicularis and Darlingtonia californica. Oecologia 1997, 112, 464–471. [Google Scholar] [CrossRef]
- Woods, H.A.; Fagan, W.F.; Elser, J.J.; Harrison, J.F. Allometric and phylogenetic variation in insect phosphorus content. Funct. Ecol. 2004, 18, 103–109. [Google Scholar] [CrossRef]
- Siefert, A.; Violle, C.; Chalmandrier, L.; Albert, C.H.; Taudiere, A.; Fajardo, A.; Aarssen, L.W.; Baraloto, C.; Carlucci, M.B.; Cianciaruso, M.V.; et al. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol. Lett. 2015, 18, 1406–1419. [Google Scholar] [CrossRef]
- Bai, Y.; Zha, X.; Zhang, J.; Chen, S. The threshold of nitrogen and phosphorus loss in runoff on degraded Ferralsols of Fujian province, southern China. Environ. Earth Sci. 2020, 79, 395. [Google Scholar] [CrossRef]
- Yi, B.; Zhang, Q.; Gu, C.; Li, J.; Abbas, T.; Di, H. Effects of different fertilization regimes on nitrogen and phosphorus losses by surface runoff and bacterial community in a vegetable soil. J. Soils Sediments 2018, 18, 3186–3196. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, P.; Chen, X.; Helmers, M.J.; Zhou, X. Runoff and sediment yield under simulated rainfall on hillslopes in the Loess Plateau of China. Soil Res. 2013, 51, 50–58. [Google Scholar] [CrossRef]
- Knight, S.L.; Mitchell, C.A. Effects of CO 2 and Photosynthetic Photon Flux on Yield, Gas Exchange and Growth Rate of Lactuca sativa L. ‘Waldmann’s Green. J. Exp. Bot. 1988, 39, 317–328. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Mean | 95% CI |
|---|---|---|
| Melastoma candidum | ||
| ALAN | −0.0434 | [−0.1147, 0.0278] |
| Daylight | 0.0006 | [−0.0736, 0.0768] |
| ALAN × Daylight | −0.0309 | [−0.0840, 0.0233] |
| Colocasia gigantea | ||
| ALAN | −0.1052 | [−0.1500, −0.0613] |
| Daylight | 0.0489 | [0.0036, 0.0934] |
| ALAN × Daylight | −0.0113 | [−0.0436, 0.0216] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, C.; Nakamura, A.; Song, X.; Katabuchi, M. Artificial Light at Night (ALAN) Influences Understory Plant Traits through Ecological Processes: A Two-Year Experiment in a Rubber Plantation in China. Ecologies 2023, 4, 704-713. https://doi.org/10.3390/ecologies4040046
Zhou C, Nakamura A, Song X, Katabuchi M. Artificial Light at Night (ALAN) Influences Understory Plant Traits through Ecological Processes: A Two-Year Experiment in a Rubber Plantation in China. Ecologies. 2023; 4(4):704-713. https://doi.org/10.3390/ecologies4040046
Chicago/Turabian StyleZhou, Cong, Akihiro Nakamura, Xiaoyang Song, and Masatoshi Katabuchi. 2023. "Artificial Light at Night (ALAN) Influences Understory Plant Traits through Ecological Processes: A Two-Year Experiment in a Rubber Plantation in China" Ecologies 4, no. 4: 704-713. https://doi.org/10.3390/ecologies4040046