
Inactivating Host Bacteria for Characterization and Use of Phages



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Phage Strains: Purification of Phages
2.2. Counting of Bacteria and Phages
2.3. Effects of PrimeStore-Based Inactivation Reagent on Bacterial and Phage Viability
2.4. Electron Microscopy (EM) of Bacteria Propagated in Liquid Culture
2.5. Electron Microscopy of Phage T4 Plaques
2.6. Detection of Endotoxin
3. Results
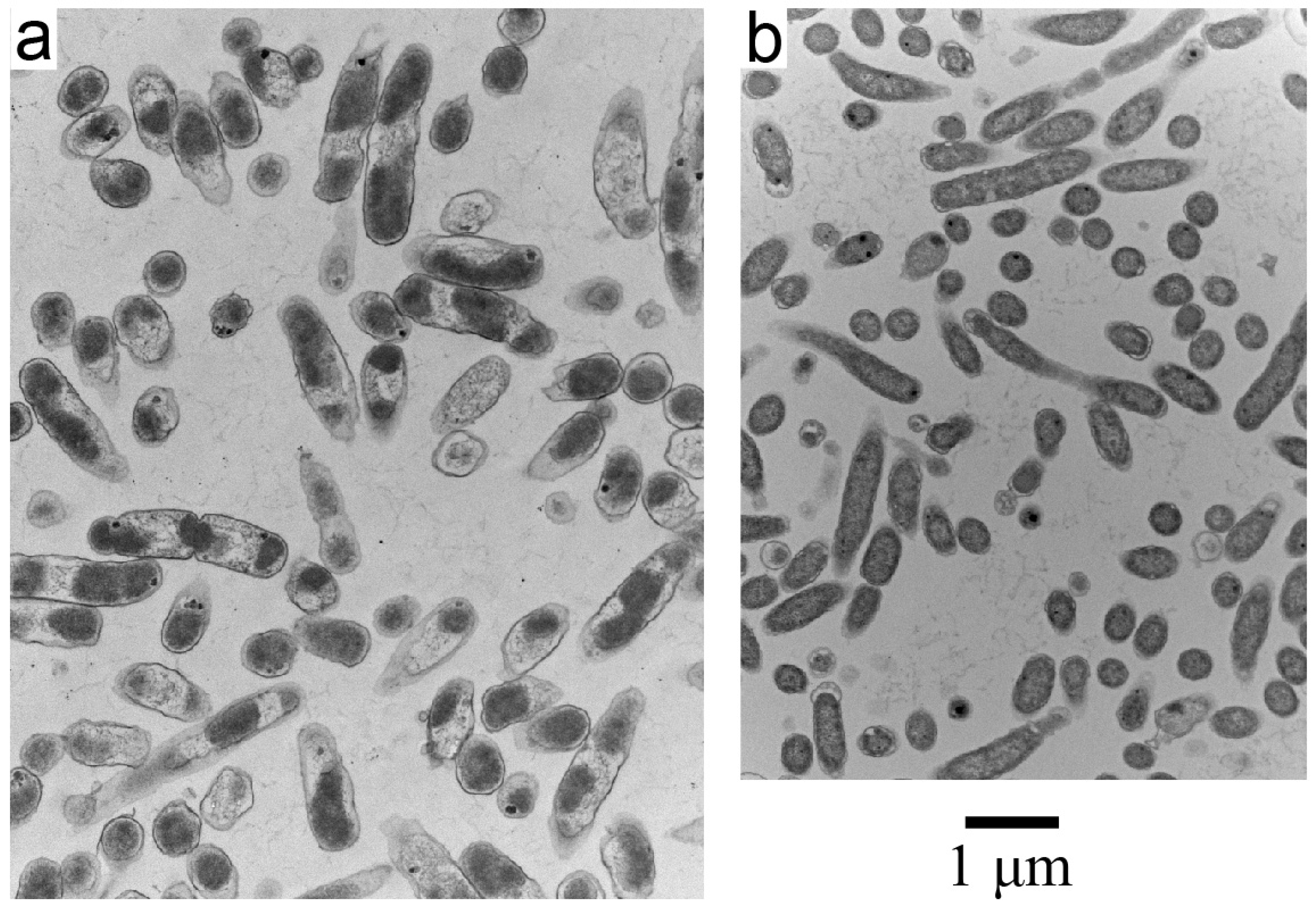
3.1. Inactivation of Gram-Negative and Gram-Positive Bacteria in Growth Medium
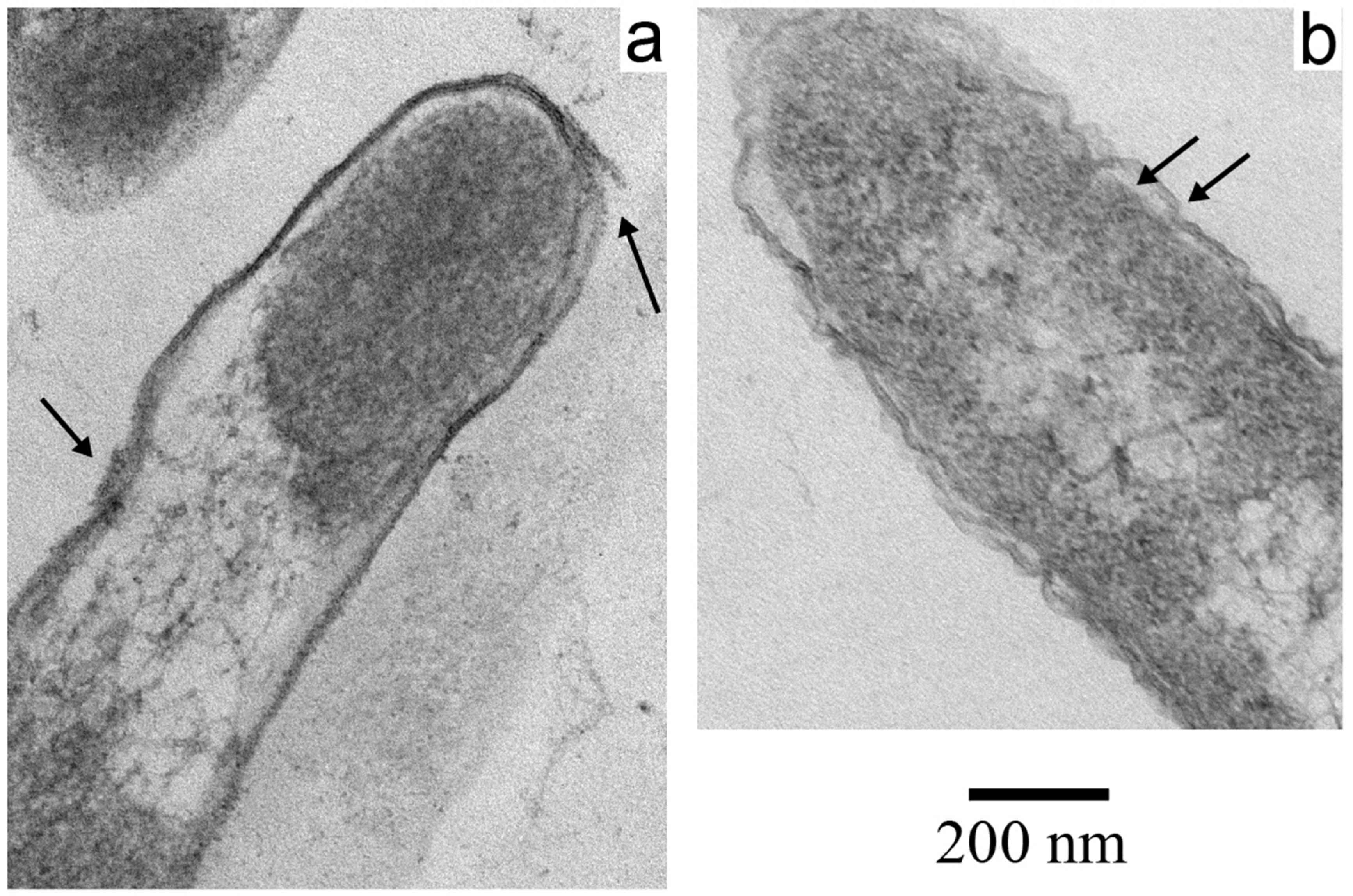
3.2. Electron Microscopy of Inactivated E. coli
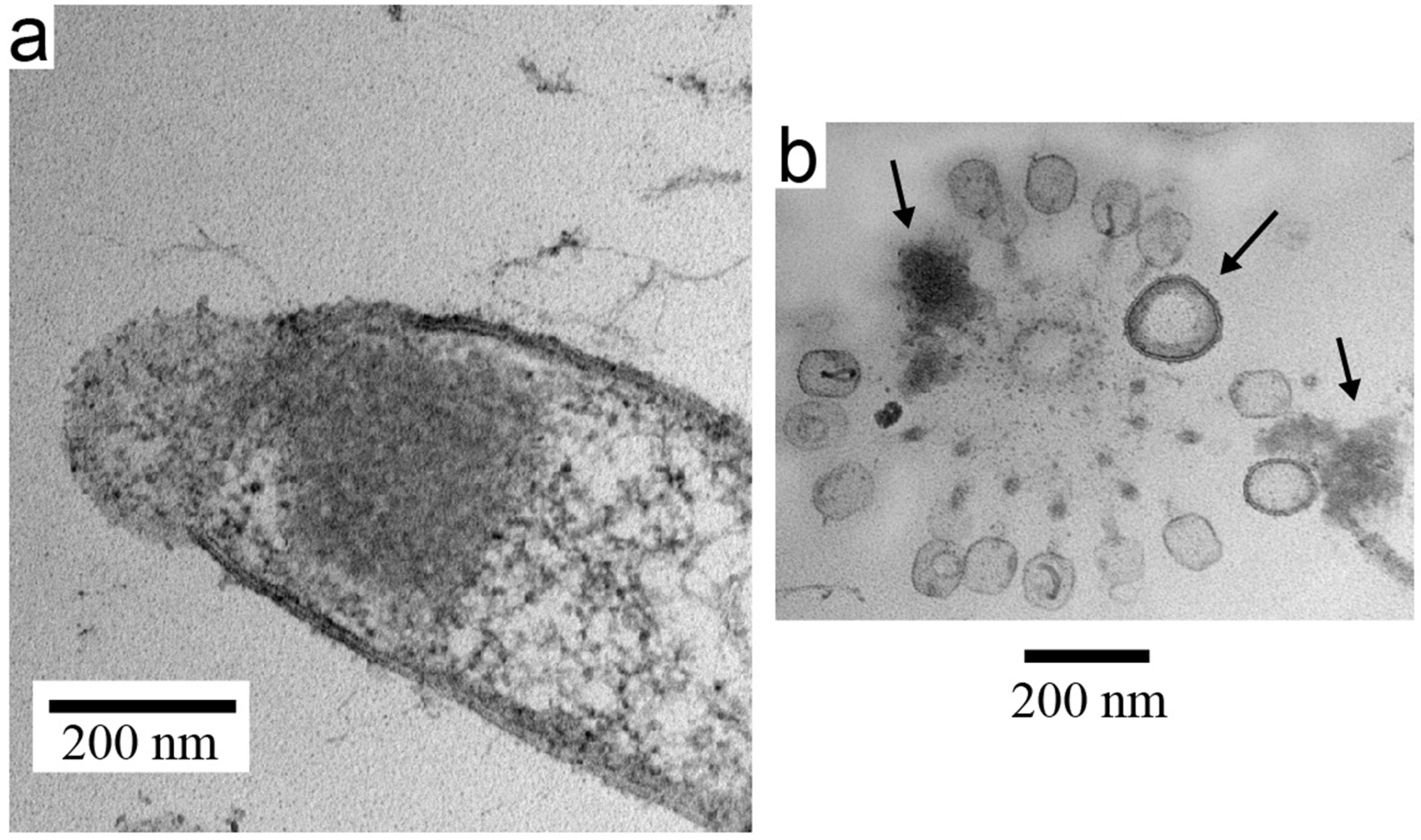
3.3. Electron Microscopy of Inactivated B. thuringiensis
3.4. Margin for Preservation of Phage Infectivity
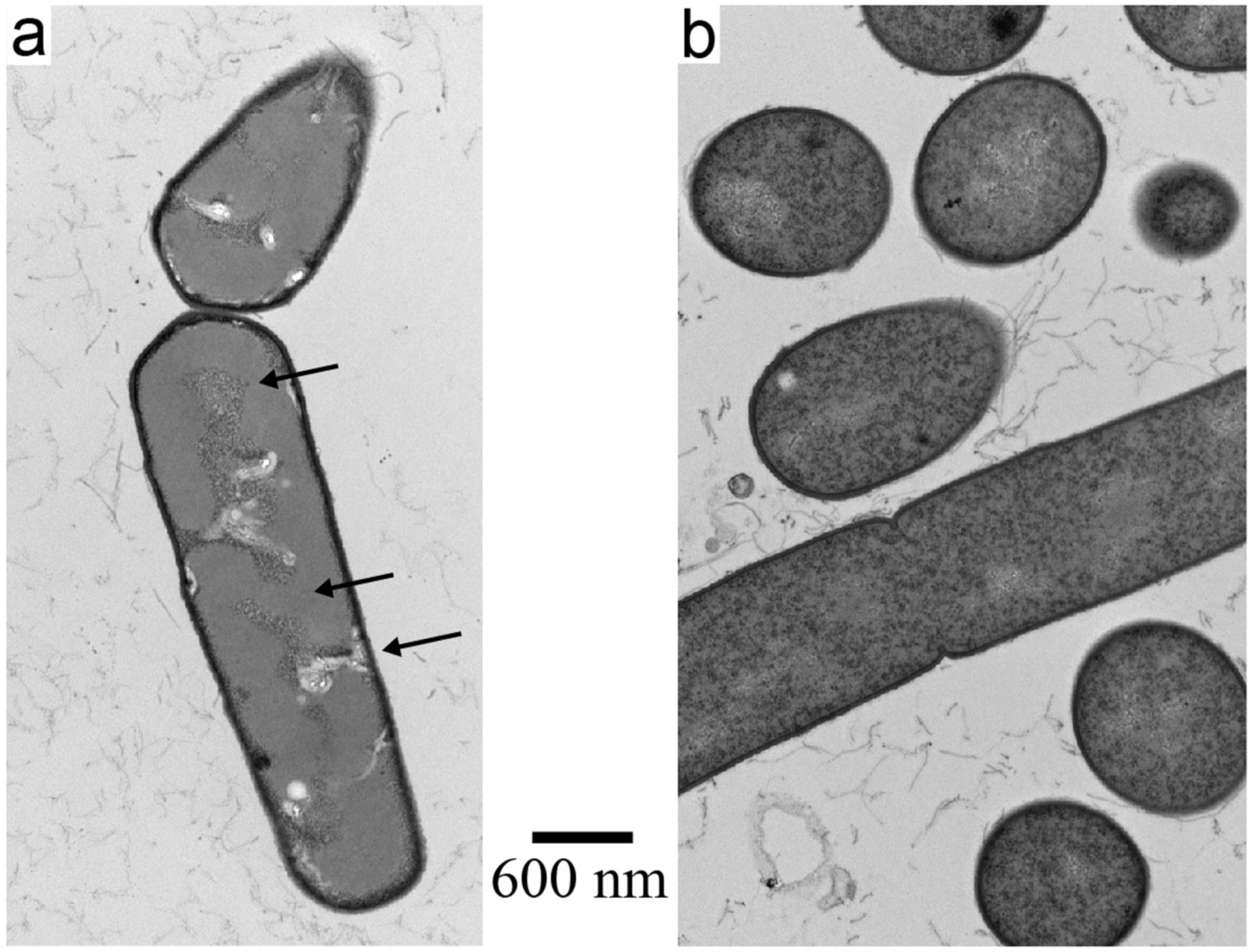
3.5. Effect of Inactivation on Release of Endotoxin
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hatfull, G.F.; Dedrick, R.M.; Schooley, R.T. Phage therapy for antibiotic-resistant bacterial infections. Annu. Rev. Med. 2022, 73, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Strathdee, S.A.; Hatfull, G.F.; Mutalik, V.K.; Schooley, R.T. Phage therapy: From biological mechanisms to future directions. Cell 2023, 186, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Żaczek, M.; Górski, A.; Grzybowski, A.; Pasternak, E.; Weber-Dąbrowska, B.; Międzybrodzki, R. How interest in phages has bloomed into a leading medical research activity in Poland. Viruses 2022, 14, 2617. [Google Scholar] [CrossRef]
- Górski, A.; Borysowski, J.; Międzybrodzki, R. Phage therapy: Towards a successful clinical trial. Antibiotics 2020, 9, 827. [Google Scholar] [CrossRef] [PubMed]
- Diallo, K.; Dublanchet, A. A century of clinical use of phages: A literature review. Antibiotics 2023, 12, 751. [Google Scholar] [CrossRef]
- Serwer, P.; Wright, E.T.; De La Chapa, J.; Gonzales, C.B. Basics for improved use of phages for therapy. Antibiotics 2021, 10, 723. [Google Scholar] [CrossRef]
- Roberts, S.M.; Aldis, M.; Wright, E.T.; Gonzales, C.B.; Lai, Z.; Weintraub, S.T.; Hardies, S.C.; Serwer, P. Siphophage 0105phi7-2 of Bacillus thuringiensis: Novel propagation, DNA, and genome-implied assembly. Int. J. Mol. Sci. 2023, 24, 8941. [Google Scholar] [CrossRef]
- Goswami, K.P.; Pugazhenthi, G. Credibility of polymeric and ceramic membrane filtration in the removal of bacteria and virus from water: A review. J. Environ. Manag. 2020, 268, 110583. [Google Scholar] [CrossRef]
- Cevallos-Mendoza, J.; Amorim, C.G.; Rodríguez-Díaz, J.M.; Montenegro, M.d.C.B.S.M. Removal of contaminants from water by membrane filtration: A review. Membranes 2022, 12, 570. [Google Scholar] [CrossRef]
- Mortimer, M.; Petersen, E.J.; Buchholz, B.A.; Holden, P.A. Separation of bacteria, protozoa and carbon nanotubes by density gradient centrifugation. Nanomaterials 2016, 6, 181. [Google Scholar] [CrossRef]
- Hyman, P. Phages for phage therapy: Isolation, characterization, and host range breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Karczewska, M.; Strzelecki, P.; Szalewska-Pałasz, A.; Nowicki, D. How to tackle bacteriophages: The review of approaches with mechanistic insight. Int. J. Mol. Sci. 2023, 24, 4447. [Google Scholar] [CrossRef] [PubMed]
- Farber, F.E.; Gradwohl, S.E.; Sanford, P.B.; Tobin, M.J.; Lee, K.J.; Gerba, C.P. Bacteriophage concentration from water by filter chromatography. J. Virol. Meth. 1983, 7, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Daum, L.T.; Worthy, S.A.; Yim, K.C.; Nogueras, M.; Schuman, R.F.; Choi, Y.W.; Fischer, G.W. A clinical specimen collection and transport medium for molecular diagnostic and genomic applications. Epidemiol. Infect. 2011, 139, 1764–1773. [Google Scholar] [CrossRef]
- Daum, L.T.; Fischer, G.W. Rapid and safe detection of SARS-CoV-2 and influenza virus RNA using onsite quantitative PCR diagnostic testing from clinical specimens collected in molecular transport medium. J. Appl. Lab. Med. 2021, 6, 1409–1416. [Google Scholar] [CrossRef]
- Clarke, C.; Smith, K.; Goldswain, S.J.; Helm, C.; Cooper, D.V.; Kerr, T.J.; Kleynhans, L.; van Helden, P.D.; Warren, R.M.; Miller, M.A.; et al. Novel molecular transport medium used in combination with Xpert MTB/RIF ultra provides rapid detection of Mycobacterium bovis in African buffaloes. Sci. Rep. 2021, 11, 7061. [Google Scholar] [CrossRef]
- Fischer, G.W.; Daum, L.T. Biological Specimen Collection/Transport Compositions and Methods. U.S. Patent 8,080,645 B2, 20 December 2011. [Google Scholar]
- Serwer, P.; Wright, E.T.; Liu, Z.; Jiang, W. Length quantization of DNA partially expelled from heads of a bacteriophage T3 mutant. Virology 2014, 456–457, 157–170. [Google Scholar] [CrossRef]
- Serwer, P.; Wright, E.T. Gated ethidium- and bleomycin-loading in phage T4 that is subsequently purified leak-free. Biophysica 2022, 2, 366–380. [Google Scholar] [CrossRef]
- Serwer, P.; Hunter, B.; Wright, E.T. Electron microscopy of in-plaque phage T3 assembly: Proposed analogs of neurodegenerative disease triggers. Pharmaceuticals 2020, 13, 18. [Google Scholar] [CrossRef]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef]
- Bahadur, R.; Chodisetti, P.K.; Reddy, M. Cleavage of Braun’s lipoprotein Lpp from the bacterial peptidoglycan by a paralog of l,d-transpeptidases, LdtF. Proc. Natl. Acad. Sci. USA 2021, 118, e2101989118. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.-L.; Sánchez, A.; Fernández-de-Bobadilla, M.D.; San-Millán, A.; Rodríguez-Beltrán, J. Bacterial subcellular architecture, structural epistasis, and antibiotic resistance. Biology 2023, 12, 640. [Google Scholar] [CrossRef]
- Fux, A.C.; Casonato Melo, C.; Michelini, S.; Swartzwelter, B.J.; Neusch, A.; Italiani, P.; Himly, M. Heterogeneity of lipopolysaccharide as source of variability in bioassays and LPS-binding proteins as remedy. Int. J. Mol. Sci. 2023, 24, 8395. [Google Scholar] [CrossRef]
- Correa, W.; Brandenburg, K.; Zähringer, U.; Ravuri, K.; Khan, T.; Von Wintzingerode, F. Biophysical analysis of lipopolysaccharide formulations for an understanding of the low endotoxin recovery (LER) phenomenon. Int. J. Mol. Sci. 2017, 18, 2737. [Google Scholar] [CrossRef]
- Mueller, M.; Lindner, B.; Kusumoto, S.; Fukase, K.; Schromm, A.B.; Seydel, U. Aggregates are the biologically active units of endotoxin. J. Biol. Chem. 2004, 279, 26307–26313. [Google Scholar] [CrossRef]
- Gioannini, T.L.; Teghanemt, A.; Zhang, D.; Levis, E.N.; Weiss, J.P. Monomeric endotoxin:protein complexes are essential for TLR4-dependent cell activation. J. Endotoxin Res. 2005, 11, 117–123. [Google Scholar] [CrossRef]
- Vermassen, A.; Leroy, S.; Talon, R.; Provot, C.; Popowska, M.; Desvaux, M. Cell wall hydrolases in bacteria: Insight on the diversity of cell wall amidases, glycosidases and peptidases toward peptidoglycan. Front. Microbiol. 2019, 10, 331. [Google Scholar] [CrossRef]
- Frirdich, E.; Vermeulen, J.; Biboy, J.; Vollmer, W.; Gaynor, E.C. Multiple Campylobacter jejuni proteins affecting the peptidoglycan structure and the degree of helical cell curvature. Front. Microbiol. 2023, 14, 1162806. [Google Scholar] [CrossRef]
- Shehadul Islam, M.; Aryasomayajula, A.; Selvaganapathy, P.R. A review on macroscale and microscale cell lysis methods. Micromachines 2017, 8, 83. [Google Scholar] [CrossRef]
- Grigorov, E.; Kirov, B.; Marinov, M.B.; Galabov, V. Review of microfluidic methods for cellular lysis. Micromachines 2021, 12, 498. [Google Scholar] [CrossRef]
- Welch, S.R.; Davies, K.A.; Buczkowski, H.; Hettiarachchi, N.; Green, N.; Arnold, U.; Jones, M.; Hannah, M.J.; Evans, R.; Burton, C.; et al. Analysis of inactivation of SARS-CoV-2 by specimen transport media, nucleic acid extraction reagents, detergents, and fixatives. J. Clin. Microbiol. 2020, 58, e01713–e01720. [Google Scholar] [CrossRef]
- Olejnik, J.; Hume, A.J.; Ross, S.J.; Scoon, W.A.; Seitz, S.; White, M.R.; Slutzky, B.; Yun, N.E.; Mühlberger, E. Art of the kill: Designing and testing viral inactivation procedures for highly pathogenic negative sense RNA viruses. Pathogens 2023, 12, 952. [Google Scholar] [CrossRef]
- Sali, W.; Patoli, D.; Pais de Barros, J.P.; Labbé, J.; Deckert, V.; Duhéron, V.; Le Guern, N.; Blache, D.; Chaumont, D.; Lesniewska, E.; et al. Polysaccharide chain length of lipopolysaccharides from Salmonella minnesota is a determinant of aggregate stability, plasma residence time and proinflammatory propensity in vivo. Front. Microbiol. 2019, 10, 1774. [Google Scholar] [CrossRef]
- Serwer, P.; Hayes, S.J.; Zaman, S.; Lieman, K.; Rolando, M.; Hardies, S.C. Improved isolation of undersampled bacteriophages: Finding of distant terminase genes. Virology 2004, 329, 412–424. [Google Scholar] [CrossRef]
- Serwer, P.; Hayes, S.J.; Thomas, J.A.; Hardies, S.C. Propagating the missing bacteriophages: A large bacteriophage in a new class. Virol. J. 2007, 4, 21. [Google Scholar] [CrossRef]
- Serwer, P.; Hayes, S.J.; Thomas, J.A.; Griess, G.A.; Hardies, S.C. Rapid determination of genomic DNA length for new bacteriophages. Electrophoresis 2007, 28, 1896–1902. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a. | Control-CFU/mL | Treat-(5 %) CFU/mL | Treat-(10 %) CFU/mL |
| Bacteria | |||
| E. coli | 9.0 × 1010 | <101 | <101 |
| B. thuringiensis | 3.5 × 109 | <101 | <101 |
| b. | |||
| Phages | Control-PFU/mL | Treat-(10 %) PFU/mL | |
| T3 | 1.8 × 1010 | 1.8 × 1010 | |
| T4 | 6.6 × 109 | 7.0 × 109 | |
| 0105phi 7-2 | 2.2 × 108 | 2.2 × 108 | |
| a. Inactivation (1 h) | b. Inactivation (8 h) | ||||
| Phages | Control | Treat | Phages | Control | Treat |
| T3 | 1.0 × 107 | 1.0 × 107 | T3 | 1.1 × 107 | 6.6 × 106 |
| T4 | 8.4 × 106 | 8.6 × 106 | T4 | 7.0 × 106 | 5.3 × 106 |
| 0105phi 7-2 | 5.1 × 106 | 5.2 × 106 | 0105phi 7-2 | 3.5 × 106 | 2.1 × 106 |
| c. Inactivation (2.5% PS) | d. Inactivation (5.0% PS) | ||||
| Phages | Control | Treat | Phages | Control | Treat |
| T3 | 1.4 × 107 | 1.5 × 107 | T3 | 1.2 × 107 | 1.1 × 107 |
| T4 | 8.9 × 106 | 9.0 × 106 | T4 | 9.2 × 106 | 9.1 × 106 |
| 0105phi 7-2 | 3.8 × 106 | 3.7 × 106 | 0105phi 7-2 | 3.4 × 106 | 3.2 × 106 |
| e. Effect of PS Dilution | |||||
| Phage | Control | Treat | 1:10 | ||
| T4 | 2.1 × 107 | 1.1 × 105 | 1.5 × 107 | ||
| Pre-Dialysis | Post-Dialysis | ||
|---|---|---|---|
| Control | Treat | Control | Treat |
| 2.7 × 108 | 1.1 × 109 | 6.9 × 107 | 1.9 × 106 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chambers, J.P.; Wright, E.T.; Hunter, B.; Serwer, P. Inactivating Host Bacteria for Characterization and Use of Phages. Biophysica 2023, 3, 558-568. https://doi.org/10.3390/biophysica3040038
Chambers JP, Wright ET, Hunter B, Serwer P. Inactivating Host Bacteria for Characterization and Use of Phages. Biophysica. 2023; 3(4):558-568. https://doi.org/10.3390/biophysica3040038
Chicago/Turabian StyleChambers, James P., Elena T. Wright, Barbara Hunter, and Philip Serwer. 2023. "Inactivating Host Bacteria for Characterization and Use of Phages" Biophysica 3, no. 4: 558-568. https://doi.org/10.3390/biophysica3040038