
Nascent Adhesion Clustering: Integrin-Integrin and Integrin-Substrate Interactions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Nascent Adhesion Clusters: Background
Finer Points Regarding the Background on Nascent Clusters
3. Clustering Model: Outline
3.1. Specific Integrin Elastic Model
3.2. Simulation Model: General Set Up
3.3. Simulation Model: Force Mediated Diffusion
3.4. Ligation vs. Bond Rupture Kinetics
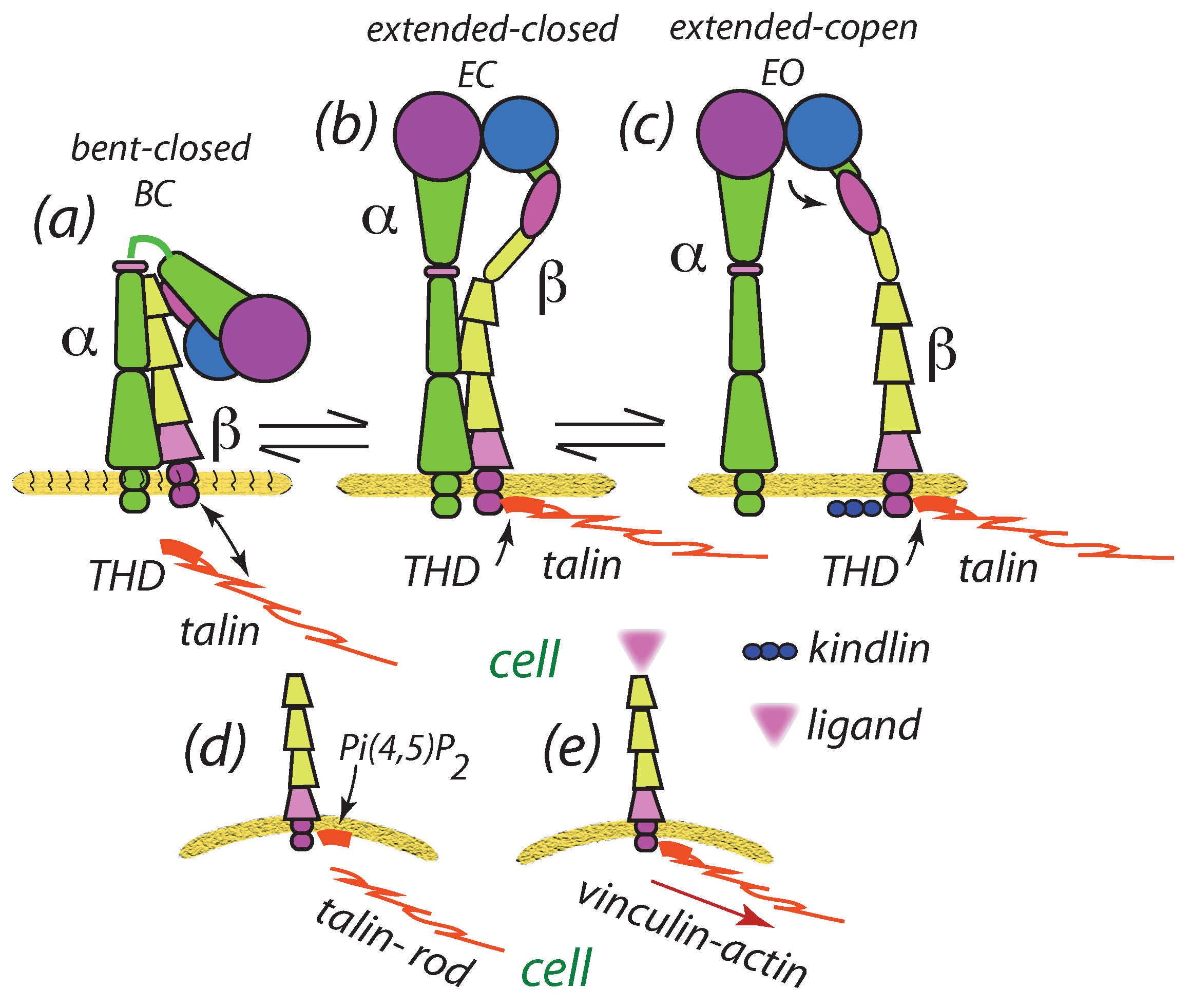
3.5. Scaffolding Proteins Affect Clustering: Talin
4. Results
4.1. Effects of Parameters
- Ligation distance, : As the force, f, within an integrin depends on , as does the integrin’s displacement field, we expect this parameter to have a first order effect on integrin migration and ligation binding lifetime. These effects are, however, complex due to the cooperative nature of integrin interaction as well as the stochastic nature of integrin migration and ligation and un-ligation behavior. We further note that the interaction forces are related to combinations of terms such as as in Equation (7), and as and , the interaction force between integrins may be expected to dominated by . However, as the internal force, f, depends on E, the rate of un-ligation will depend directly on stiffness E. Hence the effects of E, per se, are not easily anticipated a priori. For example, due to the attractive forces among the integrins we expect that increased mobilities will tend to reduce time scales for cluster formation and perhaps affect cluster size.
- Integrin diffusion coefficient, D: Both the rate of diffusion and viscous motion depend on the magnitude of D since the viscous mobility, , and D are related as . Hence we expect that D will have a first order effect in promoting clustering.
- Cell membrane-substrate stiffness, E: The effect of stiffness has already been noted and we expect that its main effect will be in determining whether clusters are stable or not; this is expected due to the effect of large integrin internal forces on the rate of un-ligation, i.e., . In terms of interaction force, the effect of E becomes more difficult to anticipate in such a stochastic process as diffusion-interaction driven clustering. Hence, we might say “we’ll see what happens and try to rationalize after”.
4.2. Simulation Results
4.2.1. Cluster-Cluster Interaction
5. Discussion
- We have demonstrated that nascent clusters should form on substrates of all rigidities, a claim made in the title of Changede and Sheetz [32,33]. This basis for their finding becomes clear upon the realization that a driving force for clustering stems from the energy reductions that follow clustering as illustrated by attractive pairwise force between integrins given in, e.g., Equation (7). As noted earlier, this force scales directly with terms such as where and , respectively, scale as E and , E being the system rigidity (stiffness). Hence, the this force does not depend strongly on system rigidity; the internal force within an integrin, however, does depend on system rigidity and that will affect ligation bond survival time. Of course, this basis may also be appreciated by the simulations of Reynar et al. [58,59] noted above; in those cases there was no substrate, but only the free energy of the deformed membrane.
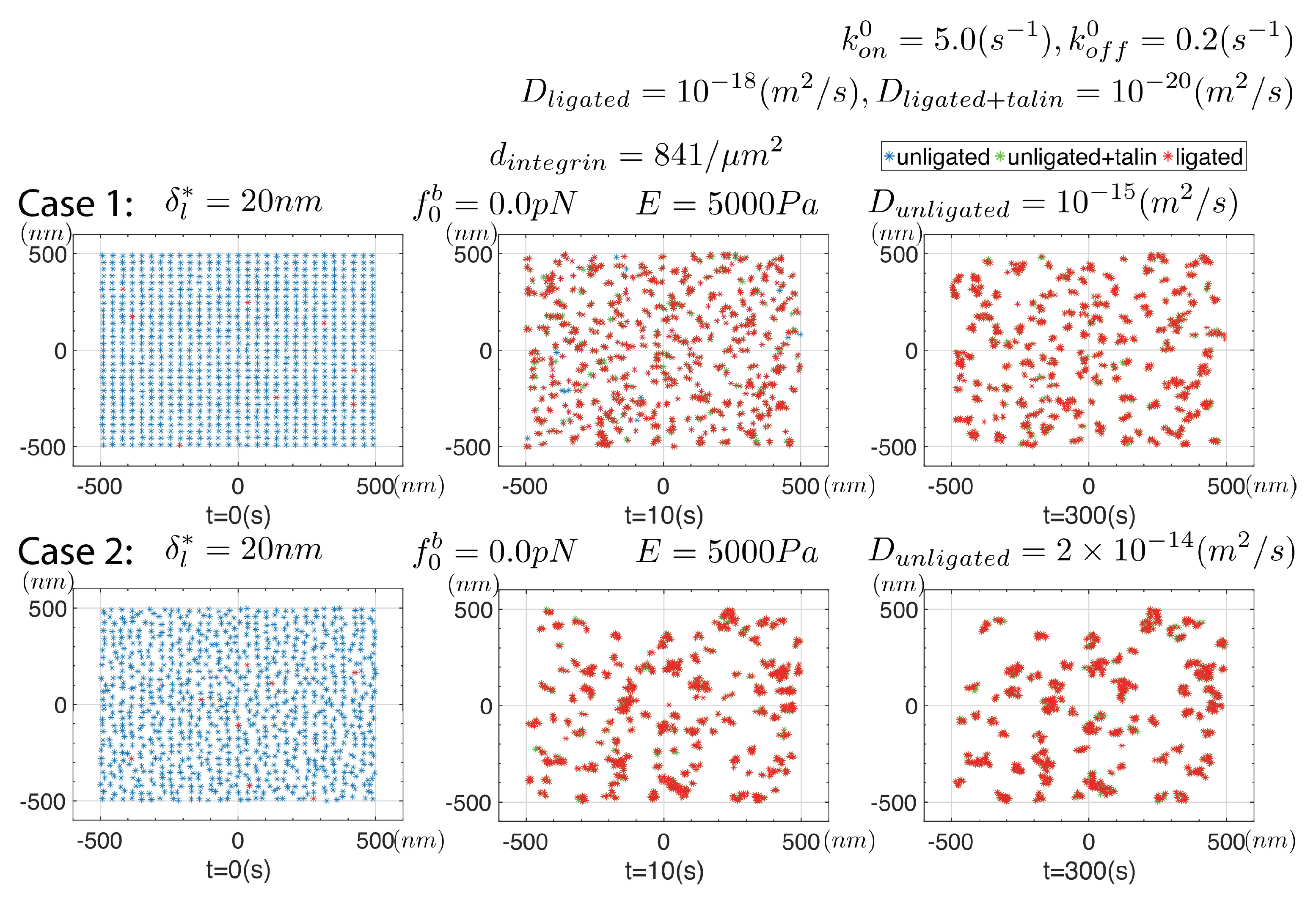
- Our simulations reveal important quantitative and qualitative effects of integrin mobility, on clustering as illustrated in Figure 10, and by comparing Figure 10 with Figure 9, as examples. First, we observe that clusters tend to be larger when the mobility of unligated integrins is increased; in the specific case studies clusters were in the size range (in Figure 9) and with a factor of 5 increase in were more in the range of (in Figure 10) in diameter. Moreover, if ligated integrins were ascribed a somewhat increased mobility, clusters were observed to “drift” so that they aggregated. To visualize why this may happen, we noted that clusters that stand apart by some distance from each other appear as “large point forces” and hence attract with a force similar in kind to that described by Equation (7) for single integrin pairs; this is the thermodynamic force driving the drifting motion. The actual mechanism for this drifting involves integrin unligation and ligation at the juxtaposed peripheries of merging clusters as illustrated in the simulation frames shown for Case #2 of Figure 10. But the question arises: what accounts for this enhanced mobility of ligated integrins? In the cases studied by Changede and Sheetz [32], involving cells on supported bilipid layers (SBLs), they reported that the ligands on SBLs were mobile. For cells adhered to an ECM or an organic substrate this would remain an open question. Nonetheless, if clusters are observed to aggregate, this offers a potential mechanism. Clearly, this sort of observation, among others, suggests that integrin mobility is an important determinant for deciphering cluster dynamics and hence experimental studies of clustering will be deficient without such information.
- On the question of integrin mobility we note the study of Rossier et al. [93] as a noteworthy example. Their study was particularly concerned with understanding the role of, in particular and containing, integrins dynamics on FAs in fibroblasts on fibronectin substrates. We have used these studies to motivate our variations in diffusion coefficient as explained in our case studies above. Accordingly, we “immobilize” integrins upon ligation where we have assumed that the integrins bind talin, and possibly kindlins as well; this is done by assigning a nearly vanishingly small value of . Rossier et al. [93] defined integrin confinement within FAs in terms of a radius, , and with that specified diffusion coefficients accordingly. This sort of detail may be incorporated into a model such as ours for further refinement.
- Finally we add that our model helps explain the patterns of integrin clustering observed in the experiments we have cited, e.g., [32,33,34], that are some of the most comprehensive conducted to date. In particular, we find that the numbers of integrins that appear in clusters appears to lie in the range of 30–50 which is again typical to what is observed experimentally [32,33]; this in itself is a noteworthy and not obvious trend. However we also note that depending on factors such as the initial density of integrins and the magnitude of diffusion coefficients numbers outside this range are indeed possible. For this reason we have made special note that future studies of nascent clustering should document the mobility of integrins as well as their expressed numbers.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Janmey, P.A.; Winer, J.P.; Murray, M.E.; Wen, Q. The hard life of soft cells. Cell Motil. Cytoskelet. 2009, 66, 597–605. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell linkage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Lo, C.M.; Wang, H.B.; Dembro, M.; Wang, Y.L. Cell movement is guided by the rigidity of the substrate. Biophys. J. 2000, 79, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Yeung, T.; Georges, P.C.; Flanagan, L.A.; Marg, B.; Ortiz, M.; Funaki, M.; Zahir, N.; Ming, W.; Weaver, V.; Janney, P.A. Effects of substrate stiffness on cell morphology, cytoskeleton structure, and adhesion. Cell Motil. Cytoskelet. 2005, 60, 24–34. [Google Scholar] [CrossRef]
- Thomas, W.E.; Discher, D.E.; Shastri, V.P. Mechanical regulation of cells by materials and tissues. MRS Bull. 2010, 35, 578–583. [Google Scholar] [CrossRef]
- Engler, A.J.; Carag-Krieger, C.; Johnson, C.P.; Raab, M.; Tang, H.Y.; Speicher, D.W.; Sanger, J.W.; Sanger, J.M.; Discher, D.E. Embryonic cardiomyocytes beat best on a matrix with heart-like elasticity: Scar-like rgiidity inhibits beating. J. Cell Sci. 2008, 121, 3794–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipski, A.M.; Pino, C.J.; Haseiton, F.R.; Chen, I.W.; Shastri, V.P. The effect of silica nanoscale-modified surfaces on cell morphology, cytoskeletal organization and function. Biomaterials 2008, 29, 3836–3846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lord, M.S.; Foss, M.; Besenbacher, F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today 2010, 5, 66–78. [Google Scholar] [CrossRef]
- Maheshwari, G.; Brown, G.; Lauffenburger, D.A.; Wells, A.; Griffith, L.G. Cell adhesion and motility depend on nanoscale RGD clustering. J. Cell Sci. 2000, 113, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Dalby, M.J.; Riehle, M.O.; Sutherland, D.S.; Agheli, H.; Curtis, A.S.G. Use of nanotopography to study mechanotransduction in fibroblasts—Methods and perspectives. Eur. J. Cell Biol. 2004, 83, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Jean, R.P.; Tan, J.L.; Liu, W.F.; Sniadecki, N.J.; Spector, A.A.; Chen, C.S. Emergent patterns of growth controlled by multicellular form and mechanics. Proc. Natl. Acad. Sci. USA 2005, 102, 11594–11599. [Google Scholar] [CrossRef] [Green Version]
- Gorfinkiel, N.; Blanchard, G.B.; Adams, R.J.; Arias, A.M. Mechanical control of global cell behavior during dorsal closure in Drosophila. Development 2009, 136, 1889–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammoto, T.; Ingber, D.E. Mechanical control of tissue and organ development. Development 2010, 137, 1407–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Weaver, V.M. Mechanics, malignancy, and metatasis: The force journey of a tumor cell. Cancer Matastasis Rev. 2009, 28, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Paszek, M.J.; Zahir, N.; Johnson, K.R.; Lakins, J.N.; Rozenberg, G.I.; Gefen, A.; Reinhart-King, C.A.; Margulies, S.S.; Dembo, M.; Boettiger, D.; et al. Tensional homeostasis and the malignant phenotype. Cancer Cell 2005, 8, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.S.; Mrksich, M.; Huang, S.; Whitesides, G.M.; Ingber, D.E. Geometric control of cell life and death. Science 1997, 276, 1425–1428. [Google Scholar] [CrossRef] [Green Version]
- Chrzanowska-Wodicka, M.; Burridge, K. Rho-stimulated contractility drives the formation of stress fibers and focal adhesions. J. Cell Biol. 1996, 133, 1403–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folkman, J.; Moscona, A. Role of cell shape in growth control. Nature 1978, 273, 354–359. [Google Scholar] [CrossRef]
- Fenteany, G.; Janmey, P.A.; Stossel, T.P. Signaling pathways and cell mechanics involved in wound closure by epithelial cell sheets. Curr. Biol. 2000, 10, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Bruguès, A.; Ester, A.; Conte, V.; Veldhuis, J.H.; Gupta, M.; Colombelli, J.; Munoz, J.J.; Brodland, G.W.; Ladoux, B.; Trepat, X. Forces driving epithelial wound healing. Nat. Phys. 2014, 10, 683–690. [Google Scholar]
- Balaban, N.Q.; Schwarz, U.S.; Riveline, D.; Goichberg, P.; Tzur, G.; Sabanay, D.; Mahalu, S.; Safran, S.; Bershadsky, A.; Addadi, L.; et al. Force and focal adhesion: A close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 2001, 2, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Beningo, K.A.; Dembro, M.; Kaverina, I.; Small, J.V.; Wang, Y.L. Nascent focal adhesions are responsible for the generation of strong propulsive forces in migrating fibroblasts. J. Cell Biol. 2001, 153, 881–888. [Google Scholar] [CrossRef] [Green Version]
- Bershadsky, A.D.; Balaban, N.Q.; Geiger, B. Adhesion-dependent cell mechanosensitivity. Annu. Cell Dev. Biol. 2003, 19, 677–695. [Google Scholar] [CrossRef] [Green Version]
- Geiger, B.; Spatz, J.P.; Bershadsky, A.D. Environmental sensing through focal adhesions. Nat. Rev. Mol. Cell Biol. 2009, 10, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Winograd-Katz, S.E.; Fassier, R.; Geiger, R.; Legate, K.R. The integrin adhesome: From genes and proteins to human disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Maartens, A.P.; Brown, N.H. Anchors and signals: The diverse roles of integrins in development. Curr. Top. Dev. Biol. 2015, 112, 233–272. [Google Scholar]
- Horton, E.R.; Humphries, J.D.; James, J.D.; Jones, M.C.; Askari, J.A.; Humphries, M.J. The integrin adhessome at a glance. J. Cell Sci. 2016, 129, 4159–4163. [Google Scholar]
- Zaidel-Bar, R.; Itzkovitz, S.; Ma’ayan, A.; Iyengar, R.; Geiger, B. Functional atlas of the integrin adesome. Nat. Cell Biol. 2007, 9, 858–867. [Google Scholar] [CrossRef]
- Roca-Cusachs, P.; Iskratsch, T.; Sheetz, M.P. Finding the weakest link—Exploring integrin-mediated mechanical molecular pathways. J. Cell Biol. 2012, 125, 3025–3038. [Google Scholar] [CrossRef] [Green Version]
- Klapholz, B.; Brown, N.H. Talin—The master of integrin adhesions. J. Cell Sci. 2017, 130, 2435–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atherton, P.; Sttchbury, B.; Wang, D.Y.; Jethwa, D.; Tsang, R.; Meiler-Rodriqhez, E.; Wang, P.; Bate, N.; Zent, R.; Barsukov, I.L.; et al. Vinculin controls talin engagement with the actomyosin machinery. Nat. Commun. 2015, 6, 10038. [Google Scholar] [CrossRef] [Green Version]
- Changede, R.; Xu, X.; Margadant, F.; Sheetz, M.P. Nascent integrin adhesions form on all matrix rigidities after integrin activation. Dev. Cell 2015, 35, 614–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changede, R.; Sheetz, M. Integrin and cadherin clusters: A robust way to organize adhesions foe cell mechanics. Bioessays 2016, 39, 1–12. [Google Scholar] [CrossRef]
- Choi, C.K.; Vicente-Manzanares, M.; Zareno, J.; Whitmore, L.A.; Mogilner, A.; Horwitz, A.R. Actin and α-actinin orchestrate the assembly anf maturation of nascent adhesions in a myosin II motor-independent manner. Nat. Cell Biol. 2008, 10, 1039–1050. [Google Scholar] [CrossRef]
- Sun, Z.; Costell, M.; Fässler, R. Integrin activation by talin, kindlin and mechanical forces. Nat. Cell Biol. 2019, 21, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Cluzel, C.; Saltel, F.; Lussi, J.; Paulhe, F.; Imhof, B.A.; Wehrle-Haller, B. The mechanisms and dynamics of αvβ3 integrin clustering in livibg cells. J. Cell Biol. 2005, 171, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schvartzman, M.; Palma, M.; Sable, J.; Abramson, J.; Hu, X.; Sheetz, M.P.; Wind, S.J. Nanolithographic control of the spatial organization of cellular adhesion receptors at the single-molecule level. Nano Lett. 2011, 11, 1306–1312. [Google Scholar] [CrossRef] [Green Version]
- Cavalcanti-Adam, E.A.; Aydin, D.; Hirechfeld-Warneken, V.C.; Spatz, J.P. Cell adhesion and response to synthetic nanopatterned environments by steering clustering and spatial location. HFSP J. 2006, 2, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Ballestram, C.; Hinz, B.; Imhof, B.E.; Wehrle-Haller, B. Marching at the front and dragging behind: Differential αVβ3-integrin turnover regulates focal adhesion behavior. J. Cell Biol. 2001, 155, 1319–1332. [Google Scholar] [CrossRef]
- Kucik, D.F. Rearrangement of integrins in avidity regulation by leukocytes. Immunol. Res. 2002, 26, 199–206. [Google Scholar] [CrossRef]
- Martino, F.; Perestrelo, A.R.; Vinarsky, V.; Pagliari, S.; Forte, G. Cellular mechanotranduction: From tension to function. Front. Physiol. 2018, 9, 824. [Google Scholar] [CrossRef]
- Wolfenson, H.; Yang, B.; Sheetz, M.P. Steps in mechanotransduction pathways that control cell morphology. Annu. Rev. Physiol. 2019, 81, 585–605. [Google Scholar] [CrossRef]
- Dorland, Y.L.; Huveneers, S. Cell-cell junctional mechanotransduction in endothelial remodeling. Cell. Mol. Life Sci. 2017, 74, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Kanyo, N.; Kovacs, K.D.; Saftics, A.; Szekacs, I.; Peter, B.; Santa-Maria, A.R.; Walter, F.R.; Dér, A.; Horvath, R. Glycocalyx regulates the strength and kinetics of cancer cell adhesion revealed by viophysical models based on high resolution label-free optical data. Sci. Rep. 2020, 10, 22422. [Google Scholar] [CrossRef]
- Shattil, S.J.; Kim, C.; Ginsberg, M.H. The final steps of integrin activation: The end game. Nat. Rev. Mol. Cell Biol. 2010, 11, 288–300. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, D.R.; Zent, R.; Grant, R.; Jasper, D.; Rees, G.; Hynes, R.O.; Ginsberg, M.H. The talin head domain binds to integrin β subunit cytoplasmic tails and regulates integrin activation. J. Biol. Chem. 1999, 274, 28071–28074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Alvarez, B.; de Pereda, J.M.; Calderwood, D.A.; Ulmer, T.S.; Critchley, D.A.; Campbell, I.D.; Ginsberg, M.H.; Liddington, R.C. Structural determinants of integrin recognition by talin. Mol. Cell 2003, 11, 49–58. [Google Scholar] [CrossRef]
- Ye, F.; Hu, G.; Taylor, D.; Ratnikov, B.; Bobkov, A.A.; McLean, M.A.; Sligar, S.G.; Taylor, K.A.; Ginsberg, M.H. Recreation of the terminal events in physilogical integrin activation. J. Cell Biol. 2010, 188, 157–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderwood, D.A.; Campbell, I.D.; Critchley, D.R. Talin and kindlins: Partners in integrin-mediated adhesion. Nat. Rev. Mol. Biol. 2013, 14, 503–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carman, C.V.; Springer, T.A. Integrin avidity regulation: Are changes in affinity and conformation underemphasized? Curr. Opin. Cell Biol. 2003, 15, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Su, Y.; Xia, W.; Humphries, M.J.; Vestweber, D.; Cabañs, C.; Lu, C.; Springer, T.A. Conformational equlibria and intrinsic affinities define integrin activation. EMBO J. 2017, 36, 629–645. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, D.A. Talin controls integrin activation. Biochem. Soc. Trans. 2004, 32, 434–437. [Google Scholar] [CrossRef]
- Kim, C.; Ye, F.; Xiaohui, H.; Ginsberg, M.H. Talin activates integrins by altering the topology of the β transmembrane domain. J. Cell Biol. 2011, 197, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Ginsberg, M.H. Integrin Activation. BMB Rep. 2014, 47, 655–659. [Google Scholar] [CrossRef]
- Du, X.; Gu, M.; Weisel, J.W.; Nagaswami, C.; Bennett, J.S.; Bowditch, R.; Ginsberg, M.H. Long range propagation of conformational changes in integrin αIIIβ3. J. Biol. Chem. 1993, 268, 23087–23092. [Google Scholar] [CrossRef]
- Theodosiou, M.; Widmaier, M.; Börrcher, R.T.; Rognoni, E.; Veeiders, M.; Bharadwaj, M.; Lambacher, A.; Austen, K.; Muller, D.J.; Zent, R.; et al. Kindlin-2 cooperates with talin to activate integrins and induces cell spreading by directly binding paxillin. eLife 2016, 5, e10130. [Google Scholar] [CrossRef]
- Saltel, F.; Martier, E.; Hytönen, V.P.; Jacqier, M.C.; Zimmermann, P.; Vogel, V.; Liu, W.; Wehrle-Haller, B. New PI (4, 5) P2-and membrane proximal integrin–binding motifs in the talin head control β3-integrin clustering. J. Cell Biol. 2009, 187, 715–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynwar, B.; Illya, G.; Hamandaris, V.A.; Muller, M.M.; Krermer, K.; Deserno, M. Aggregation and vesiculation of membrane proteins by curvature-mediated interactions. Nature 2007, 447, 461–464. [Google Scholar] [CrossRef]
- Reynwar, B.; Deserno, M. Membrane-mediated interactions between circular particles in strongly curved regieme. Soft Matter 2011, 7, 8567. [Google Scholar] [CrossRef]
- Van der Wel, C.; Vahid, A.; Šarić, A.; Idema, T.; Heinrich, D.; Kraft, D.J. Lipid membrane-mediated attraction between curvature inducing objects. Sci. Rep. 2016, 6, 32825–32834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingras, A.R.; Ginsberg, M.H. Signal transduction: Physical deformation of the membrane activates integrins. Curr. Biol. 2020, 30, R397–R400. [Google Scholar] [CrossRef]
- Chen, X.; Xie, C.; Nishida, N.; Li, Z.; Walz, T.; Springer, T.A. Requirement of open headpiece conformation for activation of leukocyte integrin αxβ2. Proc. Natl. Acad. Sci. USA 2010, 107, 14727–14732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lee, J.; Jang, J.; Ye, F.; Hong, S.J.; Petrich, B.G.; Ulmer, T.S.; Kim, C. Toplological adaptation of transmembrane domains to force-modulated lipid bilayer is a basis of sensing mechanical force. Curr. Biol. 2020, 30, 1614–1625. [Google Scholar] [CrossRef] [Green Version]
- Son, S. The Role of Membrane in Integrin-Mediated Adhesion of Cells to Surfaces. Ph.D. Thesis, Penn State University, State College, PA, USA, 2018. [Google Scholar]
- Kammerer, P.; Aretz, J.; Fässler, R. Lucky kindlin: A cloverleaf at the integrin tail. Proc. Natl. Acad. Sci. USA 2017, 114, 234–236. [Google Scholar] [CrossRef] [Green Version]
- Bachir, A.; Zareno, J.; Moissoglu, K.; Plow, E.; Gratton, E.; Horwitz, A.R. Integrin-associated complexes form hierarchically with variable stochiometry during nascent adhesion formation. Curr. Biol. 2014, 24, 1845–1853. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.L. Contact Mechanics; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Goennenwein, G.; Tanaka, M.; Hu, B.; Moroder, L.; Sackmann, E. Functional incorporation of integrins into solid supported membranes on unltrathin films of cellulose: Impact on adhesion. Biophys. J. 2003, 85, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Bagi, Z.; Couch, Y.; Broskova, Z.; Perez-Balderas, F.; Yeo, T.; Davis, S.; Fischer, R.; Sibson, N.R.; Davis, B.G.; Anthony, D.C. Extracellular vesicle integrins act as a nexus for platelet adhesion in cerebral microvessels. Sci. Rep. 2019, 9, 15847. [Google Scholar] [CrossRef] [Green Version]
- Heitjans, P.; Kärger, J. Diffusion in Condensed Matter; Springer: Heidelberg, Germany, 2005. [Google Scholar]
- Brànka, A.C.; Heyes, D.M. Algorithms for Brownian dynamics computer simulations: Multivariable case. Phys. Rev. E 1999, 60, 2381–2387. [Google Scholar] [CrossRef]
- Sun, L.; Cheng, Q.H.; Gao, H.; Zhang, Y.W. Computational modeling for cell spreading on a substrate mediated by specific interactions, long-range recruiting interactions, and diffusion of binders. Phys. Rev. E 2009, 79, 061907. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, T.; Schwarz, U.S. Stochastic dynamics of adhesion clusters under shared constant force and with rebinding. J. Chem. Phys. Lett. 2004, 121, 8997–9017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, G.I. Models for the specific adhesion of cells to cells. Science 1978, 200, 618–627. [Google Scholar] [CrossRef]
- Bell, G.I.; Dembro, M.; Bongrand, P. Competition between nonspecific and specific bonding. Biophys. J. 1984, 45, 1051–1064. [Google Scholar] [CrossRef] [Green Version]
- Dembo, M.; Torney, D.C.; Saxaman, K.; Hammer, D.A. The reaction-limited kinetics of membrane-to-surface adhesion and detachment. Proc. R. Soc. Lond. B 1988, 234, 55–83. [Google Scholar]
- Thomas, W.; Forero, M.; Yakovenko, O.; Nilsson, L.; Vicini, P.; Sokurenko, E.; Vogel, V. Catch-bond model derived from allostery explains force-activated bacterial adhesion. Biophys. J. 2006, 90, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Thomas, W. Catch bond in adhesion. Annu. Rev. Biomed. Eng. 2008, 10, 39–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaro, R.J.; Lin, K.; Zhu, Q. Mechanosensitivity occurs along the adhesome’s force train and affects traction stress. Biophys. J. 2019, 117, 1599–1614. [Google Scholar] [CrossRef]
- Paszek, M.J.; Boettiger, D.; Weaver, V.M.; Hammer, D.A. Integrin clustering is driven by mechanical resistence from the glycocalyx and the substrate. PLoS Comput. Biol. 2009, 5, e10000604. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Cheng,, Q.H.; Gao, H.J.; Zhang, Y.W. Effect of loading conditions on the dissociation behavior of catch bond clusters. J. R. Soc. Interface 2011, 9, 928–937. [Google Scholar] [CrossRef]
- Paszek, M.J.; DuFort, C.C.; Rossier, O.; Bainer, R.; Mouw, J.K.; Godula, K.; Hudak, J.E.; Lakins, J.N.; Wijekoon, A.C.; Cassereau, L.; et al. The cancer glycocalyx mechaincally primes integrin-mediated growth and survival. Nature 2014, 511, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Gao, H. Soft matrices suppress cooperation behaviors among receptor-ligand bonds in cell adhesions. PLoS ONE 2010, 5, e12342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdmann, T.; Schwartz, U.S. Stability of adhesion clusters under constant force. Phys. Rev. Lett. 2004, 92, 108102. [Google Scholar] [CrossRef] [Green Version]
- Luo, B.H.; Strokovich, K.; Walz, T.; Springer, T.A. Allosteric β1 integrin antibodies that stabilize the low affinity state by preventing the swingout of the hybrid domain. J. Biol. Chem. 2004, 279, 27466–27471. [Google Scholar] [CrossRef] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Weinbaum, S.; Tarbell, J.M.; Damiano, E.R. The structure and function of the endothelial glycocalyx. Annu. Rev. Biomed. Eng. 2007, 9, 121–167. [Google Scholar] [CrossRef]
- Soler, J.M.; Desplat-Jego, S.; Vacher, B.; Ponsonnet, L.; Fraterno, M.; Bongrand, P.; Martin, J.-M.; Foa, C. Adhesion-related glycocalyx study: Quantitative approach with imaging-spectrum in the energy filtering transmission microscope (EFTEM). FEBS Lett. 1998, 429, 89–94. [Google Scholar] [CrossRef]
- Squire, J.M.; Chew, M.; Nneji, G.; Neal, C.; Barry, J.; Michel, C. Quasi-periodic substructure in the microvessel endothelial glycocalyx: A possible explanation for molecular filtering. J. Struct. Biol. 2001, 136, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Lepzeiter, D.; Zaman, M.H. Clustered diffusion of integrins. Biophys. J. 2010, 99, L106–L108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, G.J.; Eich, C.; Torreno-Pina, J.A.; Diez-Ahedo, R.; Perez-Samper, G.; van Zanten, T.S.; Figdor, C.G.; Cambi, A.; Garcia-Parajo, M.F. Lateral mobility of individual integrin nanoclusters orchestrates the onset for leukocyte adhesion. Proc. Natl. Acad. Sci. USA 2012, 109, 4869–4874. [Google Scholar] [CrossRef] [Green Version]
- Yauch, R.L.; Felsenfeld, D.P.; Kraeft, S.K.; Chen, L.B.; Sheetz, M.P.; Hemler, M.E. Mutational evidence for control of adhesion through diffusion/Clustering Independent of ligand binding. J. Exp. Med. 1997, 186, 1347–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossier, O.; Octaeu, V.; Sibarita, J.-B.; Leduc, C.; Tessier, B.; Nair, D.; Gatterdam, V.; Destaing, O.; Albiges-Rizo, C.; Tampe, R.; et al. Integrins β1β3 exhibit distinct dynamic nanoscale organizations inside focal adhesions. Nat. Cell Biol. 2012, 14, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Chang, B.H.; Bae, Y.C. Salting out in the aqeous single-protein solution: The effect of shape factor. Biophys. Chem. 2003, 104, 523–533. [Google Scholar] [CrossRef]
- Docherty, H.; Galindo, A.; Sanz, E.; Vega, C. Investigation of the slating out of methane from aqueous electrolyte solutions using computer simulations. J. Phys. Chem. 2007, 111, 8993–9000. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, K.; Asaro, R.J. Nascent Adhesion Clustering: Integrin-Integrin and Integrin-Substrate Interactions. Biophysica 2022, 2, 34-58. https://doi.org/10.3390/biophysica2010004
Lin K, Asaro RJ. Nascent Adhesion Clustering: Integrin-Integrin and Integrin-Substrate Interactions. Biophysica. 2022; 2(1):34-58. https://doi.org/10.3390/biophysica2010004
Chicago/Turabian StyleLin, Kuanpo, and Robert J. Asaro. 2022. "Nascent Adhesion Clustering: Integrin-Integrin and Integrin-Substrate Interactions" Biophysica 2, no. 1: 34-58. https://doi.org/10.3390/biophysica2010004