
Relationship between Somatic Cell Score and Fat Plus Protein Yield in the First Three Lactations in Spanish Florida Goats

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotypic Data and Pedigree
2.2. Statistical Analysis
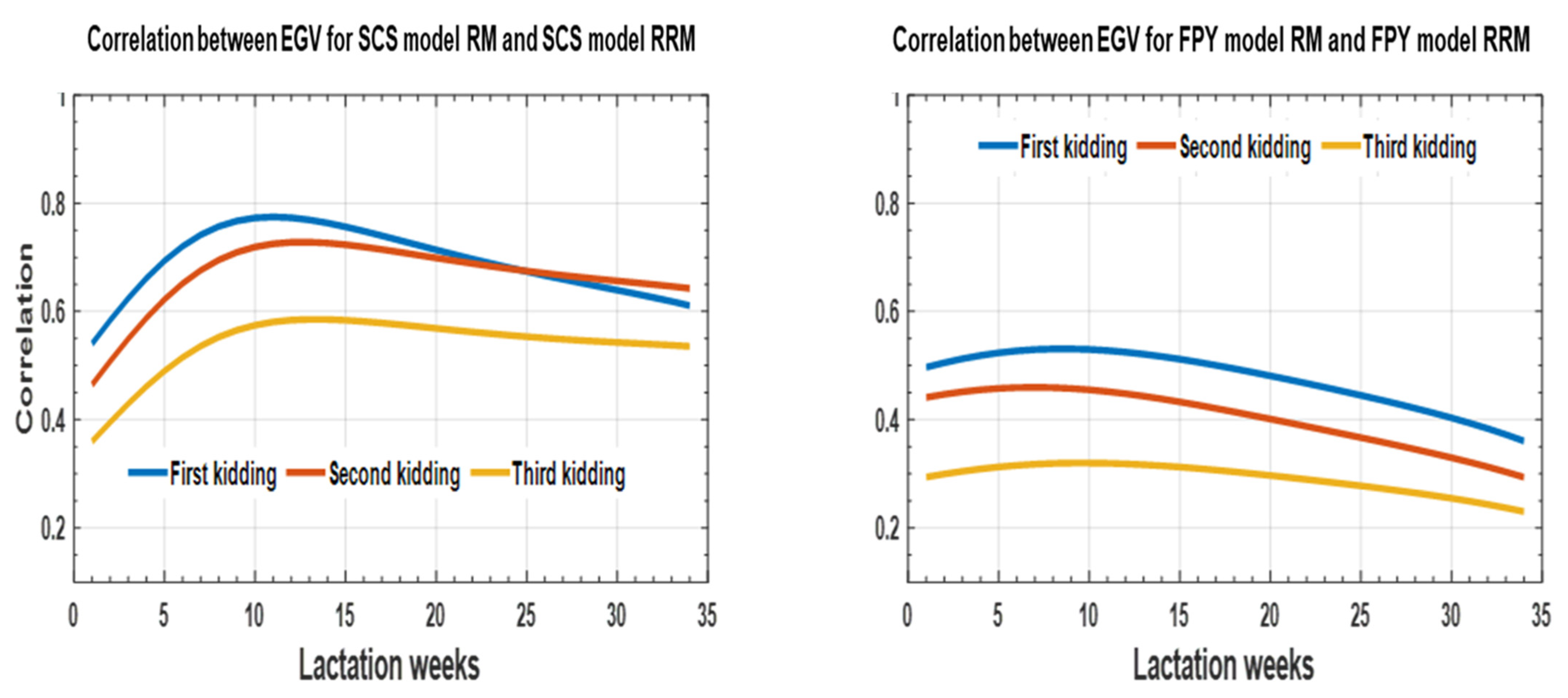
3. Results
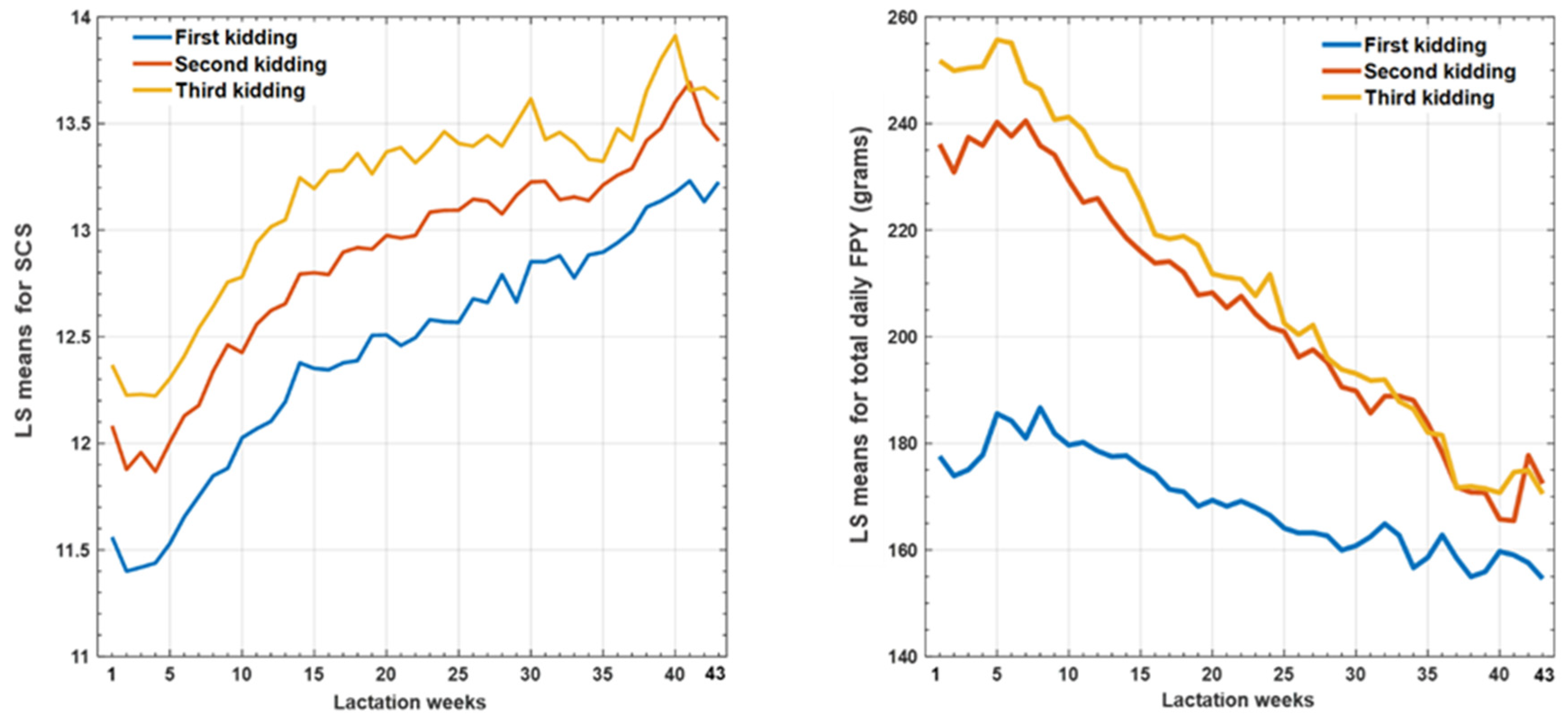
3.1. Phenotypic Parameters
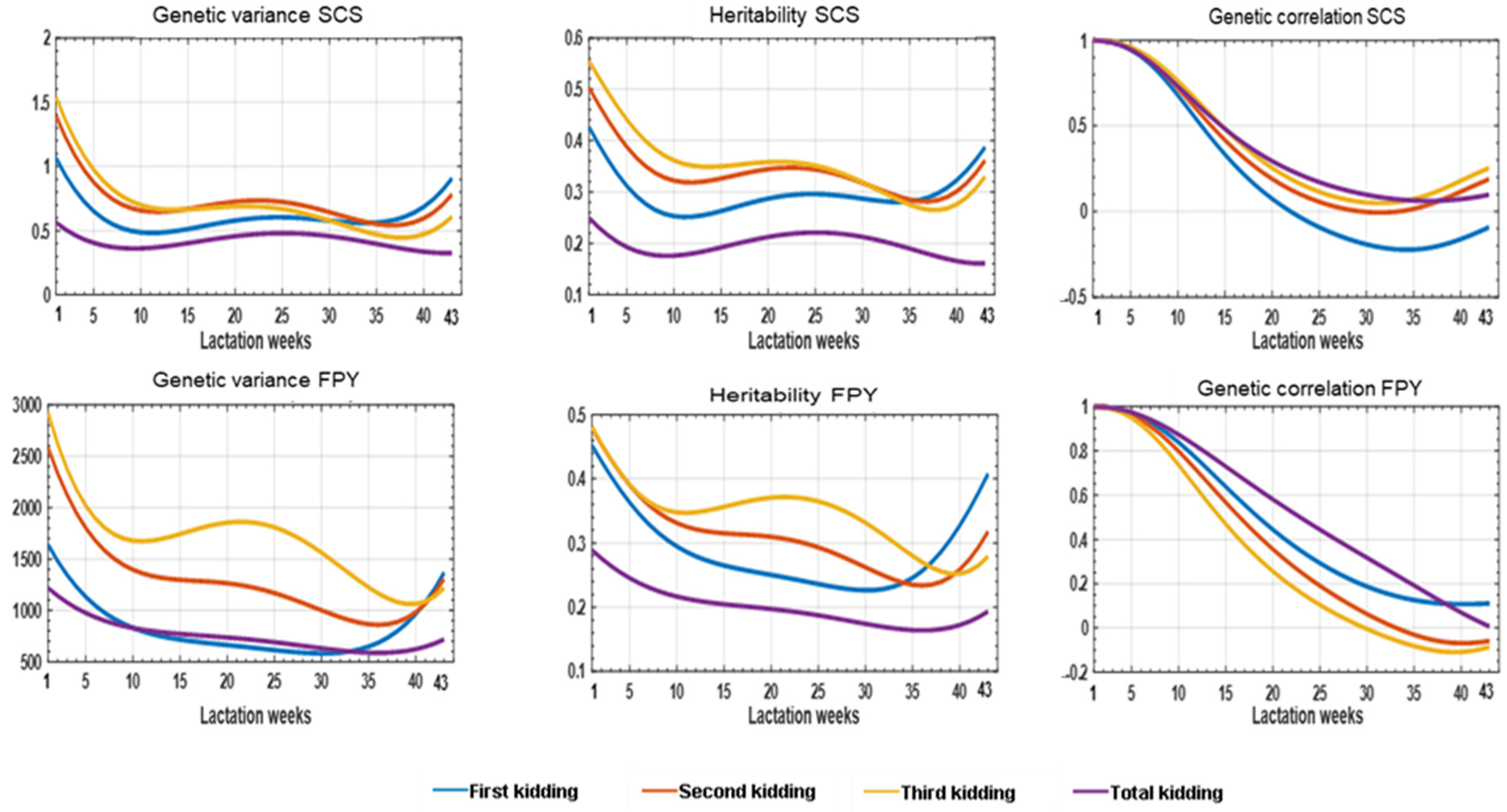
3.2. Genetic Parameters
3.2.1. Repeatability Model
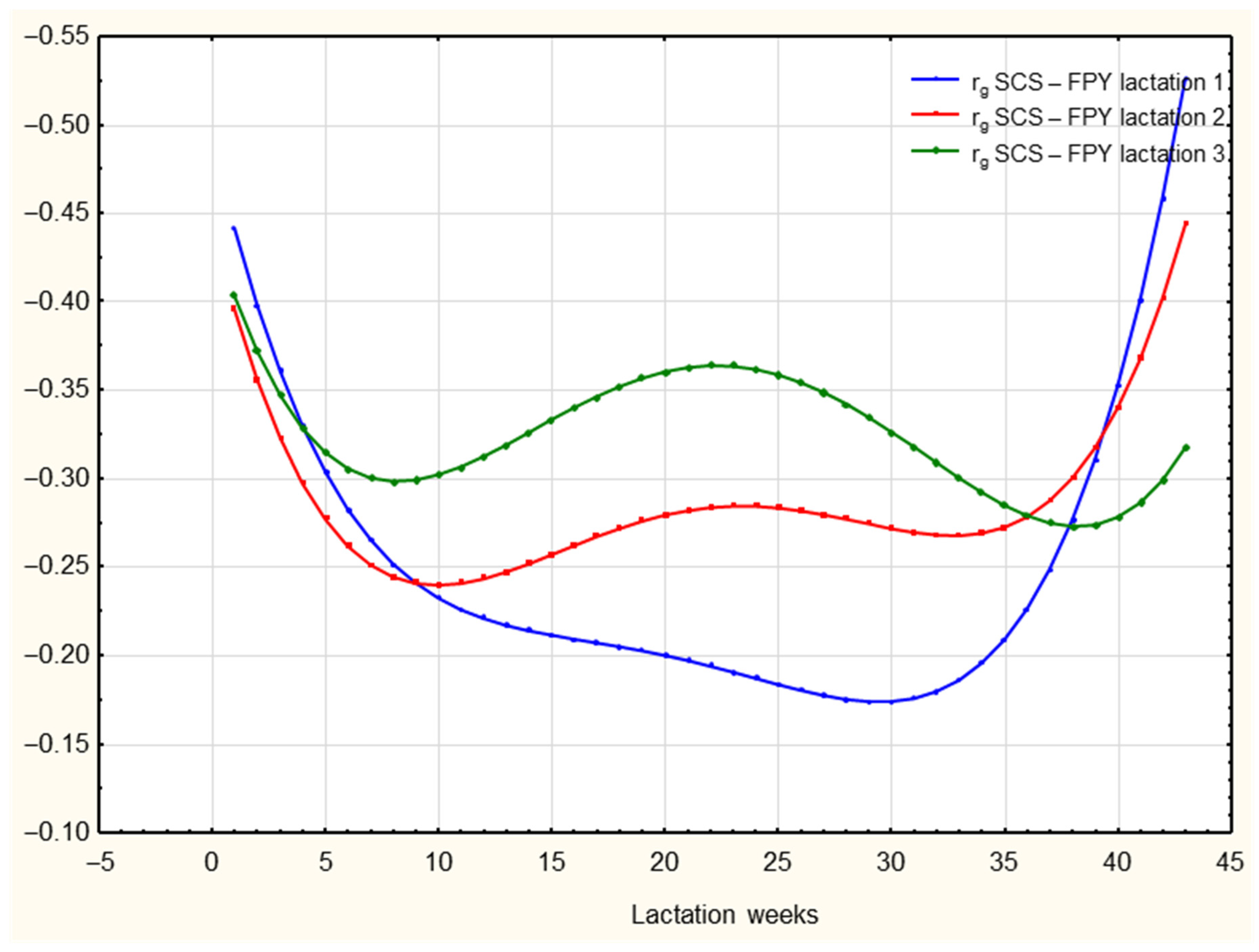
3.2.2. Random Regression Model
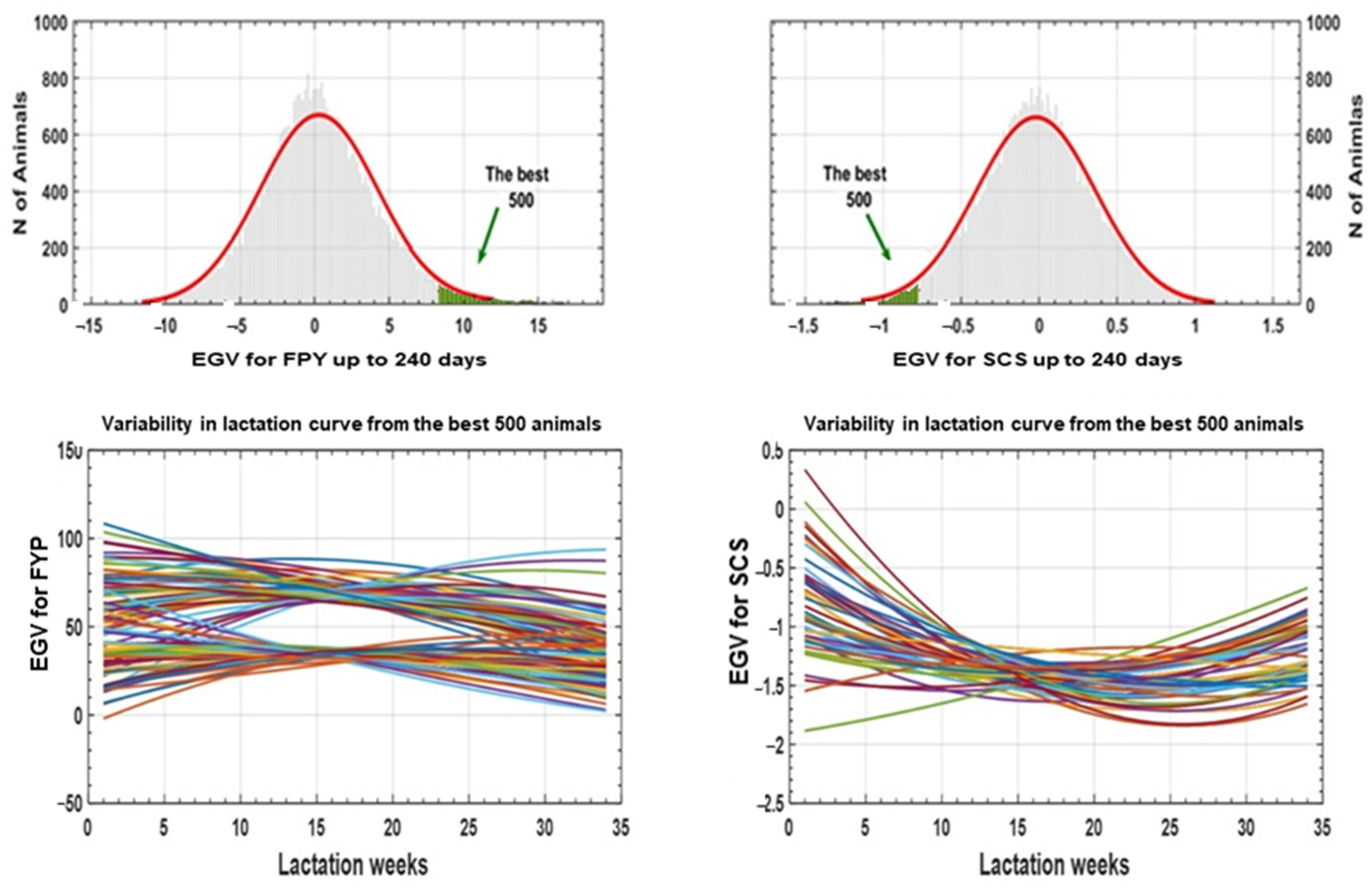
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, B.A.; Lu, C.D. Current status of global dairy goat production: An overview. Asian-Australas. J. Anim. Sci. 2019, 32, 1219. [Google Scholar] [CrossRef] [PubMed]
- Stergiadis, S.; Nørskov, N.P.; Purup, S.; Givens, I.; Lee, M.R. Comparative nutrient profiling of retail goat and cow milk. Nutrients 2019, 11, 2282. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.D.A.R.; Genís, J.M.C.; Guerrero, Y.M. Current status, challenges and the way forward for dairy goat production in Europe. Asian-Australas. J. Anim. Sci. 2019, 32, 1256. [Google Scholar] [CrossRef] [PubMed]
- Linker, R. The Goat Milk Cosmetics Market is Expected to Grow at a CAGR of over 7% during the Period 2019–2025. 2020. Available online: https://www.reportlinker.com/p05932562/Goat-Milk-Cosmetics-Market-Global-Outlook-and-Forecast.html?utm_source=GNW (accessed on 1 October 2023).
- Guerrero, A.; Abecia, J.A.; Sañudo, C. The Goat Sector in Spain: Situation, Local Breeds, and Production Systems. In Goats (Capra)-From Ancient to Modern; IntechOpen: London, UK, 2019. [Google Scholar]
- Muñoz-Mejías, M.; Menéndez-Buxadera, A.; Sánchez-Rodríguez, M.; Serradilla, J. Genetic progress attained in the selection program of Florida breed of goats in Spain. Option Méditerranéennes Ser. A 2013, 108, 134–139. [Google Scholar]
- Jimenez-Granado, R.; Molina, A.; Ziadi, C.; Sanchez, M.; Muñoz-Mejías, E.; Demyda-Peyrás, S.; Menendez-Buxadera, A. Genetic Parameters of Somatic Cell Score in Florida Goats Using Single and Multiple Traits Models. Animals 2022, 12, 1009. [Google Scholar] [CrossRef]
- Matuozzo, M.; Spagnuolo, M.S.; Hussein, H.A.; Gomaa, A.; Scaloni, A.; D’Ambrosio, C. Novel biomarkers of mastitis in goat milk revealed by MALDI-TOF-MS-based peptide profiling. Biology 2020, 9, 193. [Google Scholar] [CrossRef]
- Gómez-Gascón, L.; Galán-Relaño, A.; Cardoso-Toset, F.; Barrero-Domínguez, B.; Astorga, R.; Luque, I.; Tarradas, C.; Gómez-Laguna, J. Lactate dehydrogenase: Detecting high bacterial and somatic cells counts in goats from whole milk samples. Small Rumin. Res. 2022, 208, 106632. [Google Scholar] [CrossRef]
- Granado, R.J.; Rodríguez, M.S.; Arce, C.; Estévez, V.R. Factors affecting somatic cell count in dairy goats: A review. Span. J. Agric. Res. 2014, 12, 133–150. [Google Scholar] [CrossRef]
- Koop, G.; Nielen, M.; Van Werven, T. Bulk milk somatic cell counts are related to bulk milk total bacterial counts and several herd-level risk factors in dairy goats. J. Dairy Sci. 2009, 92, 4355–4364. [Google Scholar] [CrossRef]
- Molina, A.; Muñoz, E.; Díaz, C.; Menéndez-Buxadera, A.; Ramón, M.; Sánchez, M.; Carabaño, M.J.; Serradilla, J.M. Goat genomic selection: Impact of the integration of genomic information in the genetic evaluations of the Spanish Florida goats. Small Rumin. Res. 2018, 163, 72–75. [Google Scholar] [CrossRef]
- ARCA. Sistema Nacional De Información De Razas. Raza Caprina Florida, Ministerio De Agricultura, Alimentación Y Medio Ambiente. Gobierno De España. (mapa.gob.es). 2022. Available online: https://resolucionaprobacionflorida05_05_20_tcm30-537798.pdf (accessed on 31 July 2023).
- Bagnicka, E.; Lukaszewicz, M.; Ådnøy, T. Genetic parameters of somatic cell score and lactose content in goat s milk. J. Anim. Feed. Sci. 2016, 25, 210–215. [Google Scholar] [CrossRef]
- Scholtens, M.; Lopez-Villalobos, N.; Garrick, D.; Blair, H.; Lehnert, K.; Snell, R. Estimates of genetic parameters for lactation curves for milk, fat, protein and somatic cell score in New Zealand dairy goats. N. Z. J. Anim. Sci. Prod. 2019, 79, 177–182. [Google Scholar]
- Wiggans, G.; Shook, G. A lactation measure of somatic cell count. J. Dairy Sci. 1987, 70, 2666–2672. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, A.R.; Gogel, B.; Cullis, B.; Thompson, R.; Butler, D. ASReml User Guide (Release 3.0). 2009; VSN International Ltd.: Hemel Hempstead, UK, 2015. [Google Scholar]
- Falconer, D.S. Introduction to Quantitative Genetics; Longman Group Limited: London, UK, 1982. [Google Scholar]
- Jamrozik, J.; Schaeffer, L.R. Estimates of Genetic Parameters for a Test Day Model with Random Regressions for Yield Traits of First Lactation Holsteins. J. Dairy Sci. 1997, 80, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Weigel, K.A.; Shook, G.E. Genetic selection for mastitis resistance. Vet. Clin. Food Anim. Pract. 2018, 34, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Arnal, M.; Larroque, H.; Leclerc, H.; Ducrocq, V.; Robert-Granié, C. Estimation of genetic parameters for dairy traits and somatic cell score in the first 3 parities using a random regression test-day model in French Alpine goats. J. Dairy Sci. 2020, 103, 4517–4531. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Pérez-Guzmán, M.; Montoro, V.; Jurado, J. Genetic analysis of somatic cell count and milk traits in Manchega ewes: Mean lactation and test-day approaches. Livest. Prod. Sci. 2003, 84, 1–10. [Google Scholar] [CrossRef]
- Riggio, V.; Finocchiaro, R.; Van Kaam, J.; Portolano, B.; Bovenhuis, H. Genetic parameters for milk somatic cell score and relationships with production traits in primiparous dairy sheep. J. Dairy Sci. 2007, 90, 1998–2003. [Google Scholar] [CrossRef]
- Tolone, M.; Larrondo, C.; Yáñez, J.M.; Newman, S.; Sardina, M.T.; Portolano, B. Assessment of genetic variation for pathogen-specific mastitis resistance in Valle del Belice dairy sheep. BMC Vet. Res. 2016, 12, 158. [Google Scholar] [CrossRef]
- Windig, J.; Ouweltjes, W.; Ten Napel, J.; De Jong, G.; Veerkamp, R.; De Haas, Y. Combining somatic cell count traits for optimal selection against mastitis. J. Dairy Sci. 2010, 93, 1690–1701. [Google Scholar] [CrossRef]
- Rupp, R.; Clément, V.; Piacere, A.; Robert-Granié, C.; Manfredi, E. Genetic parameters for milk somatic cell score and relationship with production and udder type traits in dairy Alpine and Saanen primiparous goats. J. Dairy Sci. 2011, 94, 3629–3634. [Google Scholar] [CrossRef] [PubMed]
- Arnal, M.; Larroque, H.; Leclerc, H.; Ducrocq, V.; Robert-Granié, C. Genetic parameters for first lactation dairy traits in the Alpine and Saanen goat breeds using a random regression test-day model. Genet. Sel. Evol. 2019, 51, 43. [Google Scholar] [CrossRef] [PubMed]
- García-Peniche, T.; Montaldo, H.H.; Valencia-Posadas, M.; Wiggans, G.; Hubbard, S.; Torres-Vázquez, J.; Shepard, L. Breed differences over time and heritability estimates for production and reproduction traits of dairy goats in the United States. J. Dairy Sci. 2012, 95, 2707–2717. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Bustos, V.J.; Montaldo, H.H.; Torres-Hernández, G.; Pérez-Elizalde, S.; Valencia-Posadas, M.; Hernández-Mendo, O.; Shepard, L. Estimation of genetic parameters for productive life, reproduction, and milk-production traits in US dairy goats. J. Dairy Sci. 2014, 97, 2462–2473. [Google Scholar] [CrossRef]
- Stinchcombe, J.R.; Kirkpatrick, M. Genetics and evolution of function-valued traits: Understanding environmentally responsive phenotypes. Trends Ecol. Evol. 2012, 27, 637–647. [Google Scholar] [CrossRef]
- Zumbach, B.; Peters, K.; Emmerling, R.; Sölkner, J. Development of a test day model for milk sheep and goats under unfavourable structural conditions in Germany. In Proceedings of the Annual Meeting of EAAP, Bled, Slovenia, 5–9 September 2004; p. S4. [Google Scholar]
- Apodaca-Sarabia, C.; Lopez-Villalobos, N.; Blair, H.; Prosser, G. Genetic parameters for somatic cell score in dairy goats estimated by random regression. In Proceedings of the New Zealand Society of Animal Production, Canterbury, New Zealand, 24–26 June 2009; pp. 206–209. [Google Scholar]
- Oliveira, H.; Silva, F.; Siqueira, O.; Souza, N.; Junqueira, V.; Resende, M.; Borquis, R.; Rodrigues, M. Combining different functions to describe milk, fat, and protein yield in goats using Bayesian multiple-trait random regression models. J. Anim. Sci. 2016, 94, 1865–1874. [Google Scholar] [CrossRef]
- Valencia-Posadas, M.; Lechuga-Arana, A.A.; Ávila-Ramos, F.; Shepard, L.; Montaldo, H.H. Genetic parameters for somatic cell score, milk yield and type traits in Nigerian Dwarf goats. Anim. Biosci. 2022, 35, 377–384. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kidding Number | ||||
|---|---|---|---|---|
| First | Second | Third | Total | |
| Number of test-day records | 145,816 | 118,039 | 76,799 | 340,654 |
| Number of animals | 25,430 | 19,268 | 12,599 | 27,749 |
| Number of sires | 939 | 900 | 807 | 941 |
| Number of dams | 15,215 | 12,150 | 8406 | 16,243 |
| Average SCS (×103) | 11.86 (1.65) | 12.36 (1.61) | 12.72 (1.54) | 12.22 (1.65) |
| Average daily FPY (grams) | 168.1 (64.1) | 211.0 (79.1) | 219.9 (83.2) | 194.6 (77.6) |
| Lactation Number | ||||||||
|---|---|---|---|---|---|---|---|---|
| First | Second | Third | Total | |||||
| SCS | FPY | SCS | FPY | SCS | FPY | SCS | FPY | |
| Genetic variance | 0.283 | 452.9 | 0.291 | 539.0 | 0.273 | 522.4 | 0.264 | 497.8 |
| Phenotypic variance | 1.024 | 3275 | 1.066 | 4850 | 0.979 | 5287.5 | 1.051 | 4379.0 |
| Heritability | 0.276 ± 0.02 | 0.138 ± 0.01 | 0.272 ± 0.02 | 0.111 ± 0.01 | 0.279 ± 0.02 | 0.099 ± 0.01 | 0.246 ± 0.01 | 0.105 ± 0.01 |
| Genetic correlation | −0.304 ± 0.03 | −0.308 ± 0.04 | −0.477 ± 0.06 | −0.371 ± 0.04 | ||||
| Repeatibility | 0.486 ± 0.001 | 0.202 ± 0.01 | ||||||
| First | Second | Third | |
|---|---|---|---|
| First | −0.307 | 0.552 | 0.418 |
| Second | 0.366 | −0.423 | 0.609 |
| Third | 0.197 | 0.358 | −0.592 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Granado, R.; Molina, A.; Sánchez Rodríguez, M.; Ziadi, C.; Menéndez Buxadera, A. Relationship between Somatic Cell Score and Fat Plus Protein Yield in the First Three Lactations in Spanish Florida Goats. Dairy 2024, 5, 1-12. https://doi.org/10.3390/dairy5010001
Jiménez-Granado R, Molina A, Sánchez Rodríguez M, Ziadi C, Menéndez Buxadera A. Relationship between Somatic Cell Score and Fat Plus Protein Yield in the First Three Lactations in Spanish Florida Goats. Dairy. 2024; 5(1):1-12. https://doi.org/10.3390/dairy5010001
Chicago/Turabian StyleJiménez-Granado, Rocío, Antonio Molina, Manuel Sánchez Rodríguez, Chiraz Ziadi, and Alberto Menéndez Buxadera. 2024. "Relationship between Somatic Cell Score and Fat Plus Protein Yield in the First Three Lactations in Spanish Florida Goats" Dairy 5, no. 1: 1-12. https://doi.org/10.3390/dairy5010001