
A Molecular View of the Surface Pressure/Area Per Lipid Isotherms Assessed by FTIR/ATR Spectroscopy
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lipids and Chemicals
2.2. Surface Pressure vs. Area per Molecule of Lipid Monolayers
2.3. ATR-FTIR Spectroscopy
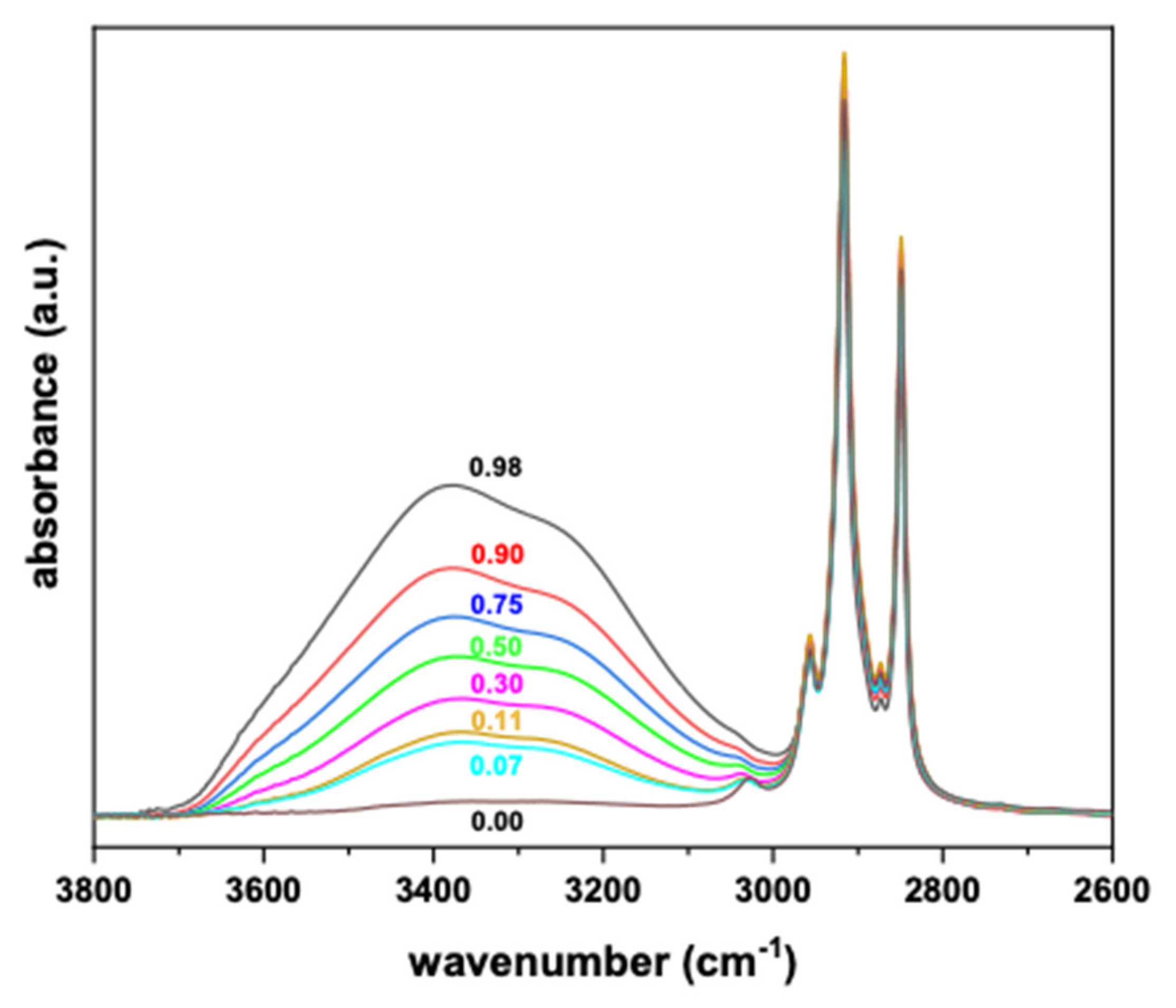
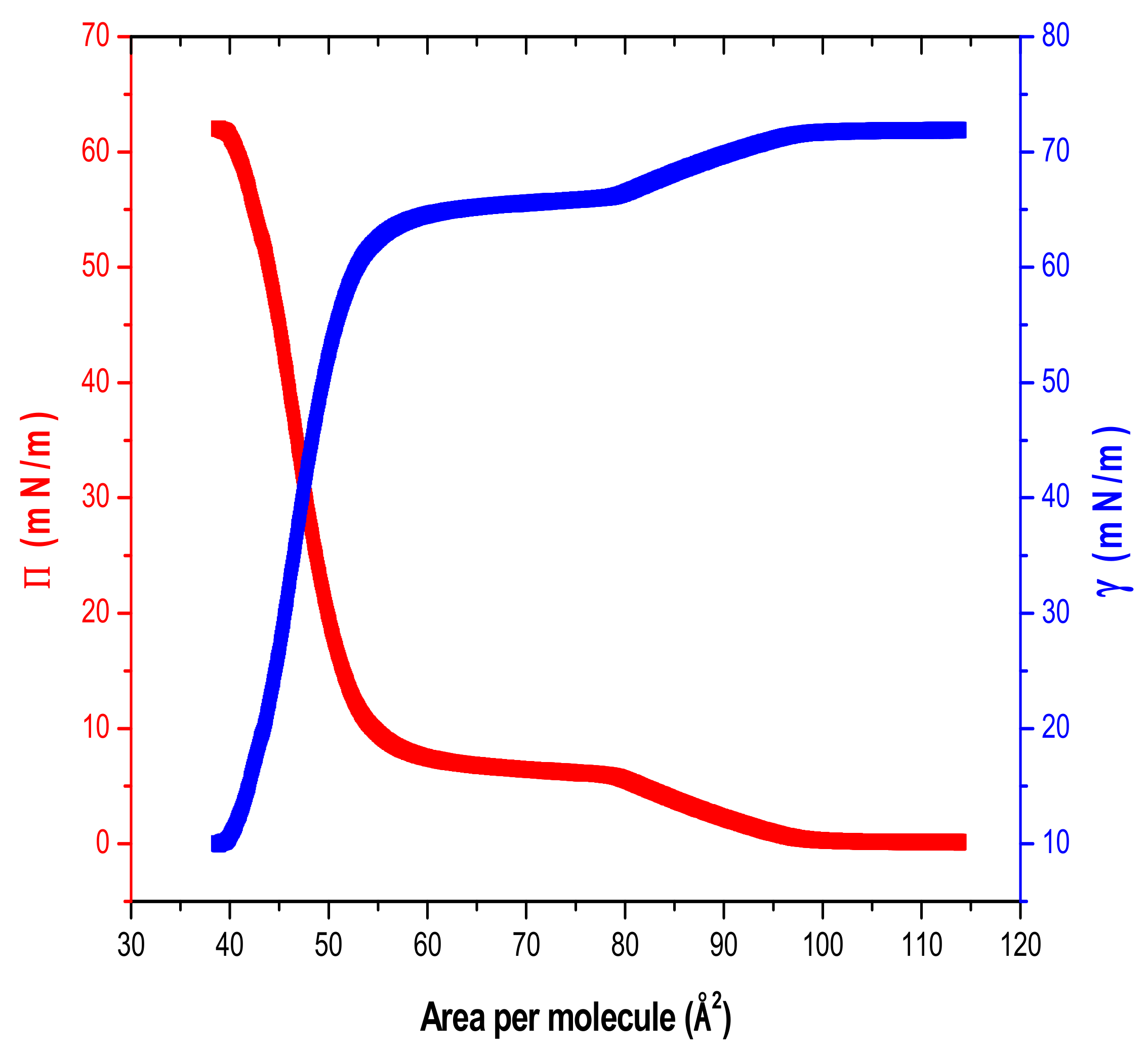
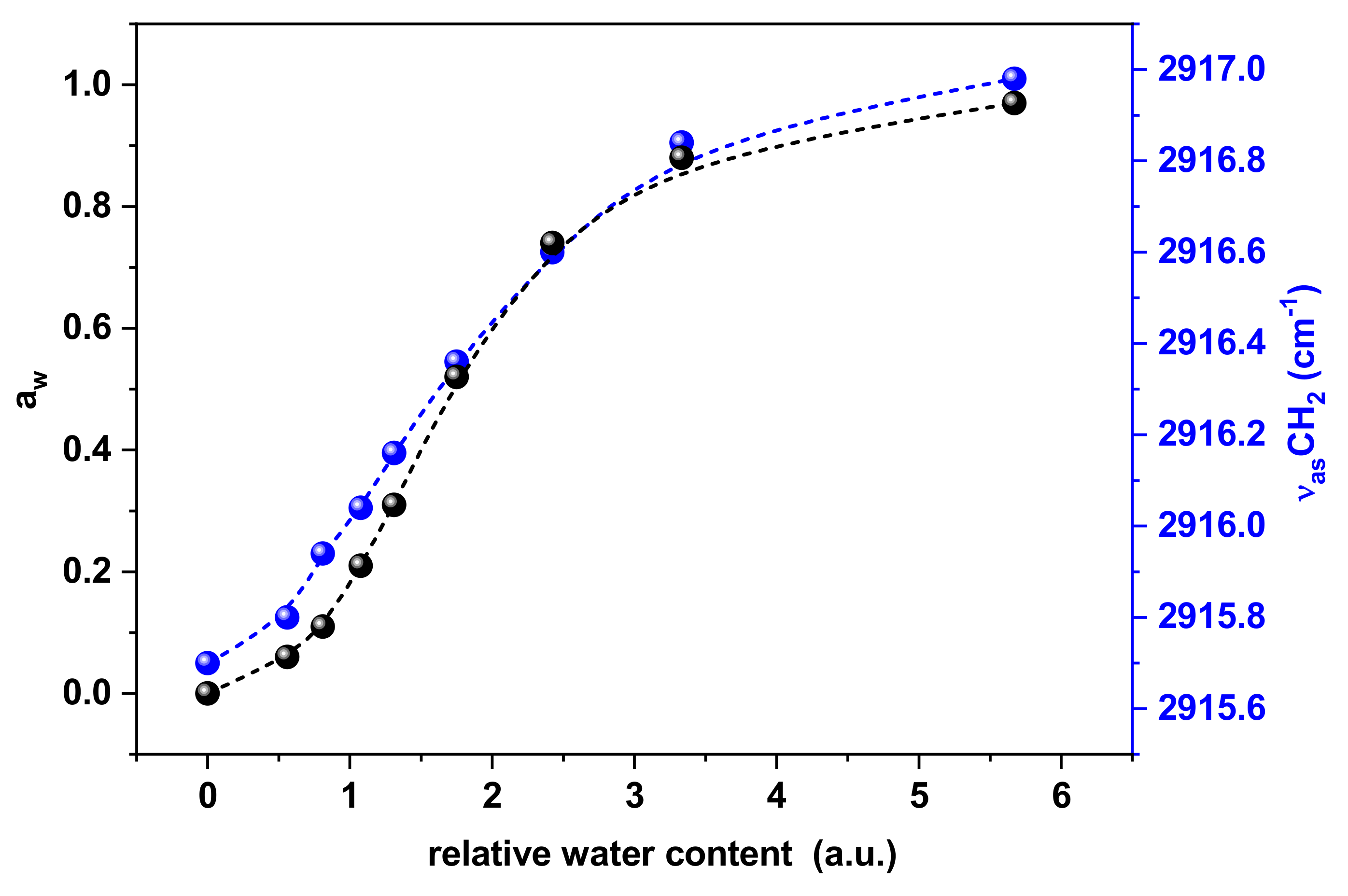
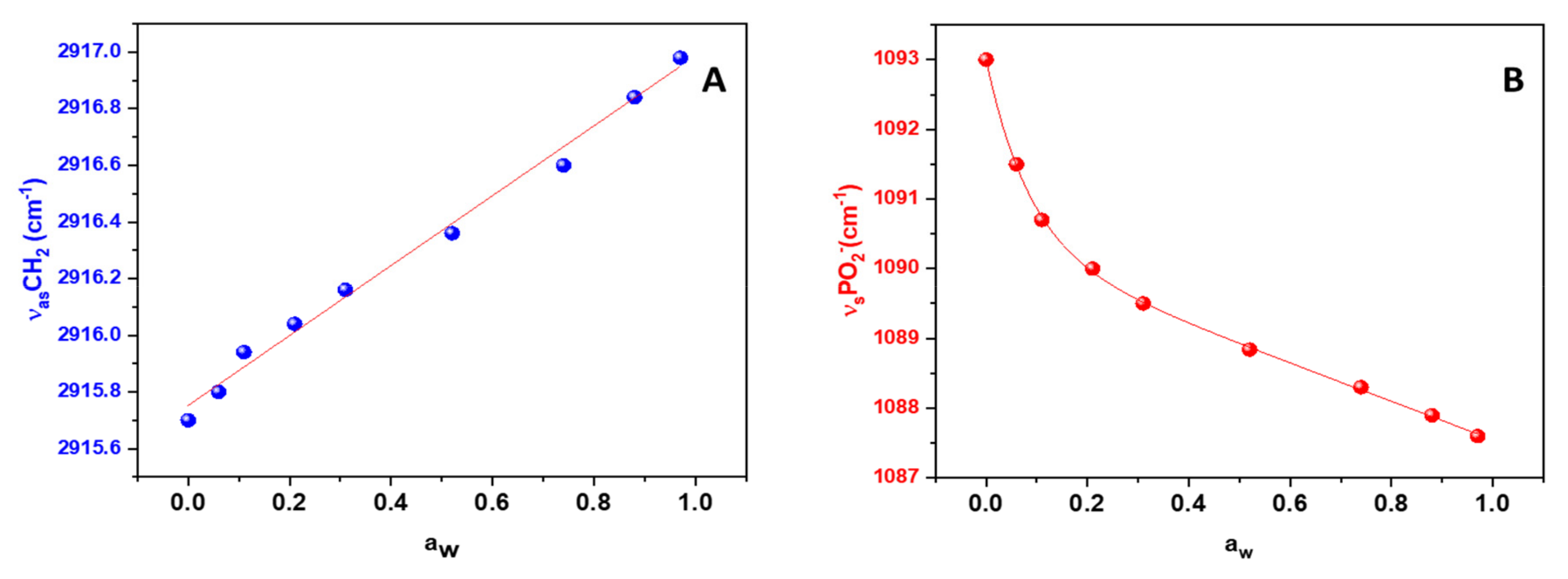
3. Results
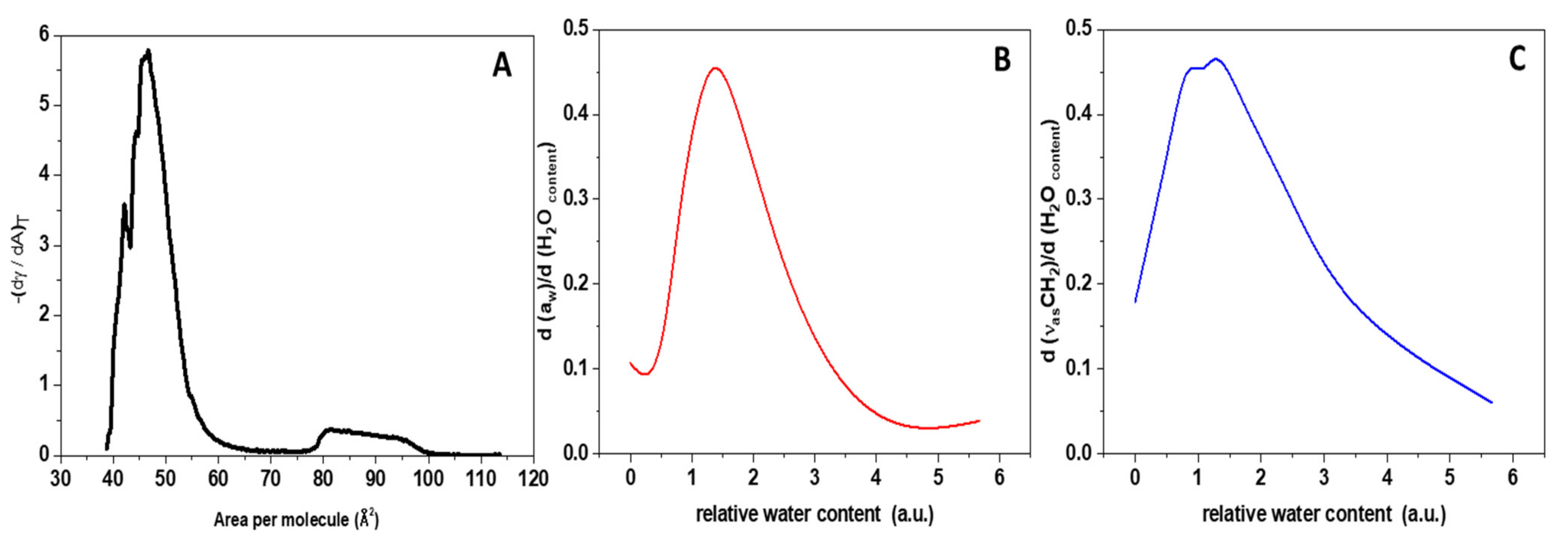
Surface Tension and Water Activity Curves
4. Discussion
Surface Tension and CH2 Exposure:
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Marsh, D. Lateral pressure in membranes. Biochim. Biophys. Acta-Rev. Biomembr. 1996, 1286, 183–223. [Google Scholar] [CrossRef]
- Duncan, S.L.; Larson, R.G. Comparing experimental and simulated pressure-area isotherms for DPPC. Biophys. J. 2008, 94, 2965–2986. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, D.H.; Brockman, H.L. Regulation of the surface pressure of lipid monolayers and bilayers by the activity of water: Derivation and application of an equation of state. Proc. Natl. Acad. Sci. USA 1988, 85, 4285–4289. [Google Scholar] [CrossRef] [Green Version]
- Frias, M.; Disalvo, E. Breakdown of classical paradigms in relation to membrane structure and functions. Biochim. Biophys. Acta-Biomembr. 2020, 1863, 183512. [Google Scholar] [CrossRef]
- Pinto, O.; Disalvo, E. A new model for lipid monolayer and bilayers based on thermodynamics of irreversible processes. PLoS ONE 2019, 14, e0212269. [Google Scholar] [CrossRef] [Green Version]
- Pinto, O.A.; Bouchet, A.M.; Frias, M.A.; Disalvo, E.A. Microthermodynamic interpretation of fluid states from FTIR measurements in lipid membranes: A Monte Carlo study. J. Phys. Chem. B 2014, 118, 10436–10443. [Google Scholar] [CrossRef]
- Calero, C.; Franzese, G. Membranes with different hydration levels: The interface between bound and unbound hydration water. J. Mol. Liq. 2019, 273, 488–496. [Google Scholar] [CrossRef]
- Tristram-Nagle, S. Use of X-ray and neutron scattering methods with volume measurements to determine lipid bilayer structure and number of water molecules/lipid. In Membrane Hydration; Springer: Berlin, Germany, 2015; pp. 17–43. [Google Scholar]
- Nickels, J.D.; Katsaras, J. Water and Lipid Bilayers. In Membrane Hydration; Springer: Berlin, Germany, 2015; pp. 45–67. [Google Scholar]
- Rosa, A.S.; Cejas, J.P.; Disalvo, E.A.; Frías, M.A. Correlation between the hydration of acyl chains and phosphate groups in lipid bilayers: Effect of phase state, head group, chain length, double bonds and carbonyl groups. Biochim. Biophys. Acta-Biomembr. 2019, 1861, 1197–1203. [Google Scholar] [CrossRef]
- Ball, P. Water as an active constituent in cell biology. Chem. Rev. 2008, 108, 74–108. [Google Scholar] [CrossRef]
- Jendrasiak, G.L. The hydration of phospholipids and its biological significance. J. Nutr. Biochem. 1996, 7, 599–609. [Google Scholar] [CrossRef]
- Milhaud, J. New insights into water–phospholipid model membrane interactions. Biochim. Biophys. Acta-Biomembr. 2004, 1663, 19–51. [Google Scholar] [CrossRef]
- Disalvo, E.A.; Bouchet, A.M.; Frias, M.A. Connected and isolated CH2 populations in acyl chains and its relation to pockets of confined water in lipid membranes as observed by FTIR spectrometry. Biochim. Biophys. Acta 2013, 1828, 1683–1689. [Google Scholar] [CrossRef] [Green Version]
- Young, J.F. Humidity control in the laboratory using salt solutions—A review. J. Appl. Chem. 1967, 17, 241–245. [Google Scholar] [CrossRef]
- O’brien, F. The control of humidity by saturated salt solutions. J. Sci. Instrum. 1948, 25, 73. [Google Scholar] [CrossRef]
- Rosa, A.S.; Disalvo, E.A.; Frias, M.A. Water behaviour at the phase transition of phospholipid matrixes assesed by FTIR. J. Phys. Chem. B 2020, 124, 6236–6244. [Google Scholar] [CrossRef]
- Rosa, A.S.; Cutro, A.C.; Frias, M.A.; Disalvo, E.A. Interaction of Phenylalanine with DPPC Model Membranes: More Than a Hydrophobic Interaction. J. Phys. Chem. B 2015, 119, 15844–15847. [Google Scholar] [CrossRef]
- Wolkers, W.F.; Oldenhof, H.; Glasmacher, B. Dehydrating phospholipid vesicles measured in real-time using ATR Fourier transform infrared spectroscopy. Cryobiology 2010, 61, 108–114. [Google Scholar] [CrossRef]
- Disalvo, E.A.; Pinto, O.A.; Martini, M.F.; Bouchet, A.M.; Hollmann, A.; Frias, M.A. Functional role of water in membranes updated: A tribute to Trauble. Biochim. Biophys. Acta 2015, 1848, 1552–1562. [Google Scholar] [CrossRef] [Green Version]
- Defay, R.; Prigogine, I.; Bellemans, A. Surface Tension and Adsorption; Wiley: Hoboken, NJ, USA, 1966. [Google Scholar]
- Marčelja, S.; Radić, N. Repulsion of interfaces due to boundary water. Chem. Phys. Lett. 1976, 42, 129–130. [Google Scholar] [CrossRef]
- Alarcón, L.M.; Frías, M.d.; Morini, M.A.; Sierra, M.B.; Appignanesi, G.A.; Disalvo, E.A. Water populations in restricted environments of lipid membrane interphases. Eur. Phys. J. E 2016, 39, 94. [Google Scholar] [CrossRef]
- Miyazaki, K.; Schweizer, K.; Thirumalai, D.; Tuinier, R.; Zaccarelli, E. The Asakura–Oosawa theory: Entropic forces in physics, biology, and soft matter. J. Chem. Phys. 2022, 156, 080401. [Google Scholar] [CrossRef]
- Chattopadhyay, M.; Krok, E.; Orlikowska, H.; Schwille, P.; Franquelim, H.G.; Piatkowski, L. Hydration Layer of Only a Few Molecules Controls Lipid Mobility in Biomimetic Membranes. J. Am. Chem. Soc. 2021, 143, 14551–14562. [Google Scholar] [CrossRef]
- Wiggins, P. Life depends upon two kinds of water. PLoS ONE 2008, 3, e1406. [Google Scholar] [CrossRef]
- Disalvo, E.; Lairion, F.; Martini, F.; Almaleck, H.; Diaz, S.; Gordillo, G. Water in biological membranes at interfaces: Does it play a functional role? Anales de la Asociación Química Argentina. Asoc. Química Argent. 2004, 92, 1–22. [Google Scholar]
- Disalvo, E.A.; Frias, M.A. Water state and carbonyl distribution populations in confined regions of lipid bilayers observed by FTIR spectroscopy. Langmuir 2013, 29, 6969–6974. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Disalvo, E.A.; Rosa, A.S.; Cejas, J.P.; Frias, M.A. A Molecular View of the Surface Pressure/Area Per Lipid Isotherms Assessed by FTIR/ATR Spectroscopy. Colloids Interfaces 2022, 6, 54. https://doi.org/10.3390/colloids6040054
Disalvo EA, Rosa AS, Cejas JP, Frias MA. A Molecular View of the Surface Pressure/Area Per Lipid Isotherms Assessed by FTIR/ATR Spectroscopy. Colloids and Interfaces. 2022; 6(4):54. https://doi.org/10.3390/colloids6040054
Chicago/Turabian StyleDisalvo, E. Anibal, Antonio Sebastian Rosa, Jimena P. Cejas, and Maria A. Frias. 2022. "A Molecular View of the Surface Pressure/Area Per Lipid Isotherms Assessed by FTIR/ATR Spectroscopy" Colloids and Interfaces 6, no. 4: 54. https://doi.org/10.3390/colloids6040054