
Distribution and Abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Benin, West Africa

 , , , , , , and
, , , , , , and
Abstract
:1. Background
2. Methods
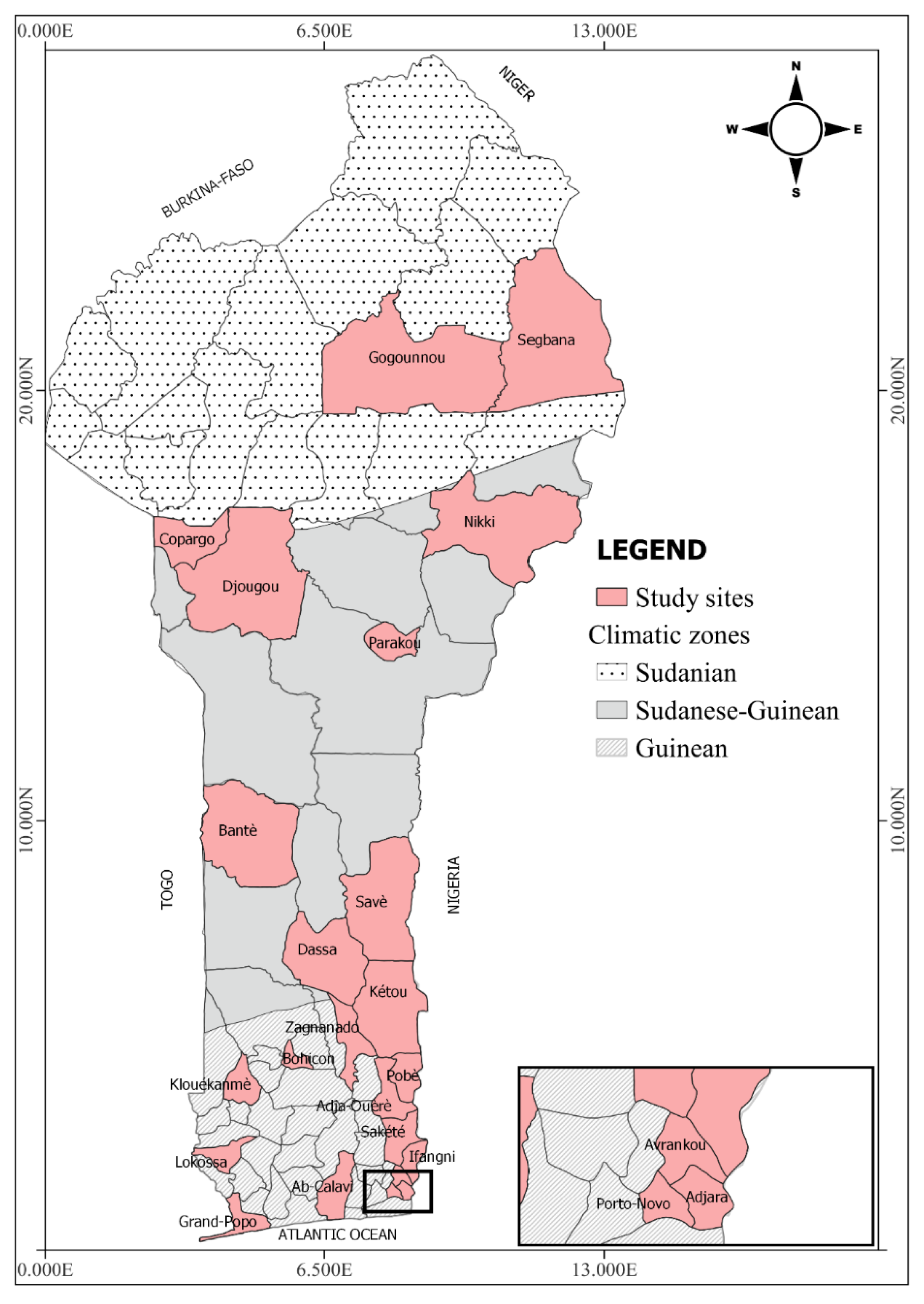
2.1. Study Area
2.1.1. Area of Degraded Forests with a Subequatorial Climate
2.1.2. Area of Savannas with a Sudano-Guinean Climate
2.1.3. Area of Savannahs with a Sudanian Climate
2.2. Mosquito Collection Techniques
2.2.1. Collection of Mosquito Immature Stages
- -
- Domestic containers: flowerpots, drums, cups, water storage containers (cement-made cisterns and earthen jars), buckets, garbage cans, pet water bowls, bottles, mortars and barrels.
- -
- Discarded containers: plastic bags, abandoned machinery (refrigerators, freezers) and tin cans.
- -
- Tires
- -
- Natural breeding sites: fruit shells, plant leaf axils, coconut and tree and root holes.
- -
- Others: wheelbarrows, abandoned canoes and cars, and pipes in the ground.
2.2.2. Using the Ovitrap Method
2.2.3. Human Landing Catch (HLC)
2.3. Morphological and Molecular Identification
2.4. Ethical Considerations
2.5. Data Analysis
3. Results
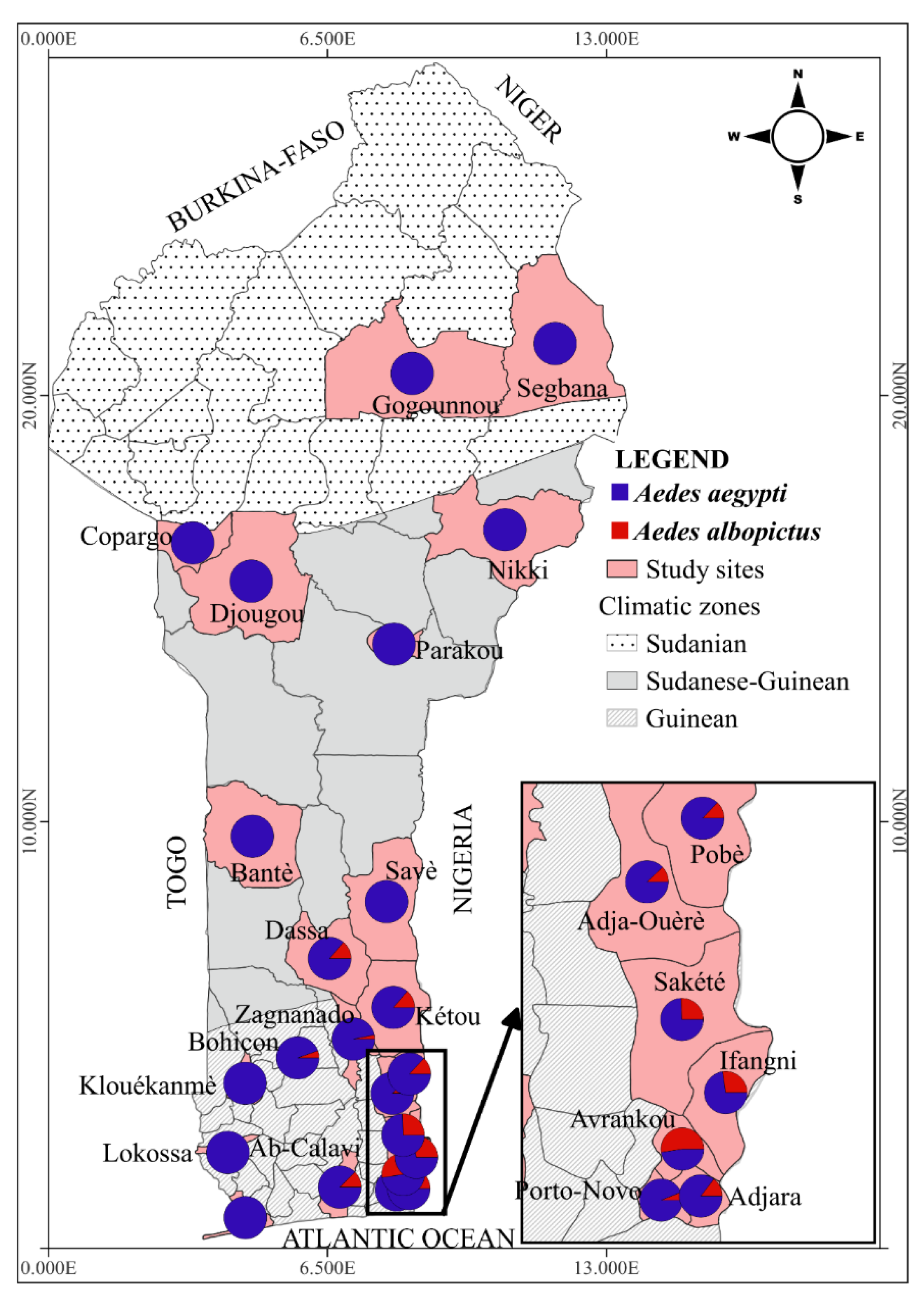
3.1. Diversity of the Aedes Mosquito Species in Benin
3.2. Distribution and Abundance of Aedes Mosquitoes across the Study Sites
3.3. Molecular Identification of Ae. albopictus
3.4. Human Biting Rate (HBR) in Ae. aegypti and Ae. albopictus
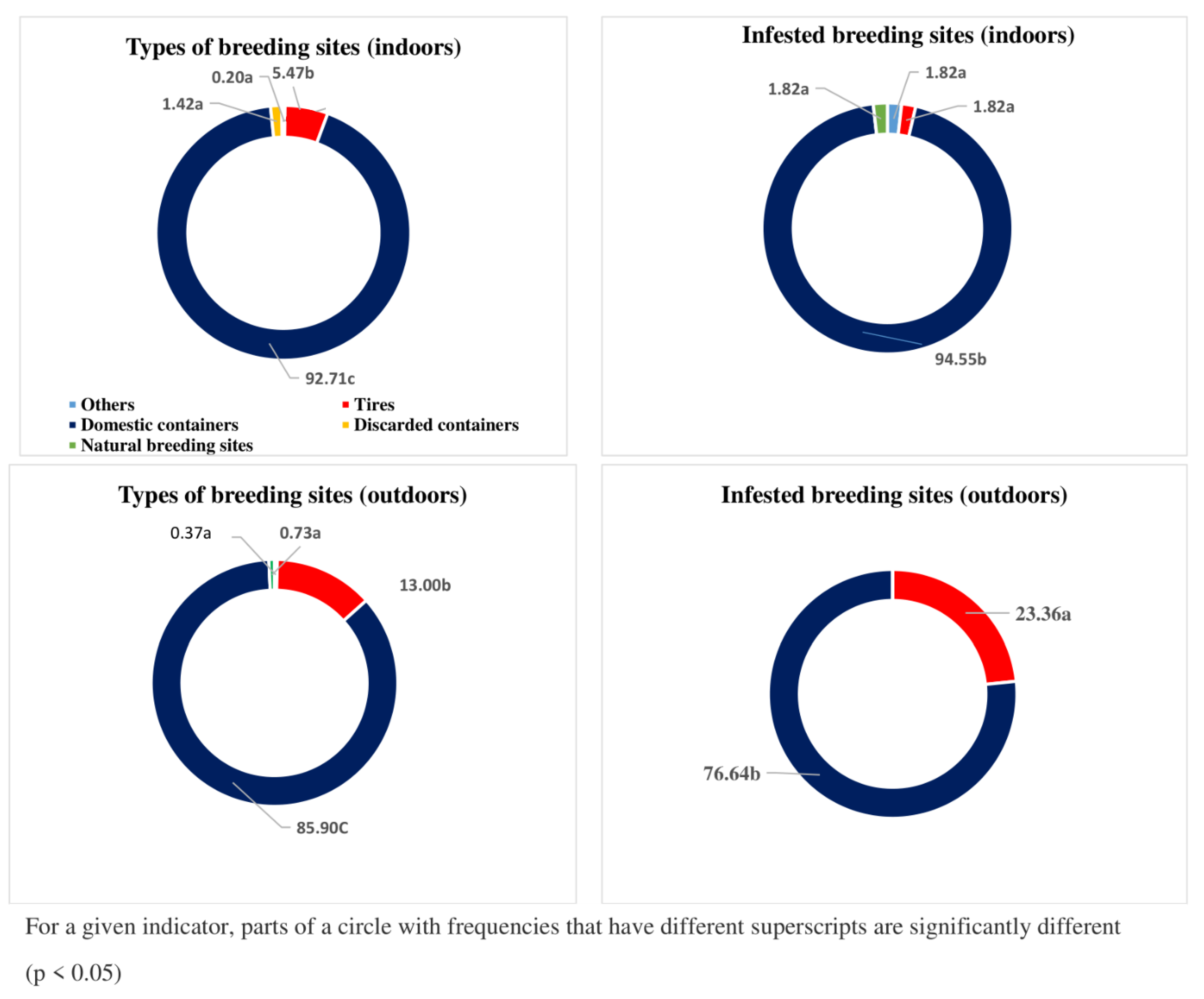
3.5. Type of Breeding Sites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Bortel, W.V.; et al. The global compendium of Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Lwande, O.W.; Obanda, V.; Lindström, A.; Ahlm, C.; Evander, M.; Näslund, J.; Bucht, G. Globe-Troting Aedes aegypti and Aedes albopictus: Risk factors for arbovirus pandemics. Vector Borne Zoonotic Dis. 2020, 20, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Eritja, R.; Palmer, J.R.B.; David, R.; Sanpera-Calbet, I.; Bartumeus, F. Direct evidence of adult Aedes albopictus dispersal by car. Sci. Rep. 2017, 7, 14399. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, B.L.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Justicia, A. Pathways for introduction and dispersal of invasive Aedes mosquito species in Europe: A review. J. Eur. Mosq. Control Assoc. 2020, 38, 1–10. [Google Scholar]
- Kamgang, B.; Ngoagouni, C.; Manirakiza, A.; Nakoune, E.; Paupy, C.; Kazanji, M. Temporal patterns of abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) and mitochondrial DNA analysis of Ae. albopictus in the Central African Republic. PLoS Negl. Trop. Dis. 2013, 7, e2590. [Google Scholar] [CrossRef]
- Marini, G.; Manica, M.; Arnoldi, D.; Inama, E.; Rosà, R.; Rizzoli, A. Influence of temperature on the life-cycle dynamics of Aedes albopictus population established at temperate latitudes: A laboratory experiment. Insects 2020, 11, 808. [Google Scholar] [CrossRef]
- Ferreira-de-Lima, V.H.; Câmara, D.C.P.; Honório, N.A.; Lima-Camara, T.N. The Asian tiger mosquito in Brazil: Observations on biology and ecological interactions since its first detection in 1986. Acta Trop. 2020, 205, 105386. [Google Scholar] [CrossRef]
- Simard, F.; Nchoutpouen, E.; Toto, J.C.; Fontenille, D. Geographic distribution and breeding site preference of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) in Cameroon, Central Africa. J. Med. Entomol. 2005, 42, 726–731. [Google Scholar] [CrossRef]
- Leta, S.; Beyene, T.J.; De Clercq, E.M.; Amenu, K.; Kraemer, M.U.G.; Revie, C.W. Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes albopictus. Int. J. Infect. Dis. 2018, 67, 25–35. [Google Scholar] [CrossRef]
- Girard, M.; Nelson, C.B.; Picot, V.; Gublerd, D.J. Arboviruses: A global public health threat. Vaccine 2020, 38, 3989–3994. [Google Scholar] [CrossRef] [PubMed]
- Cornel, A.J.; Hunt, R.H. Aedes albopictus in Africa? First records of live specimens in imported tires in Cape Town. J. Am. Mosq. Control Assoc. 1991, 7, 107–108. [Google Scholar] [PubMed]
- Savage, H.M.; Ezike, V.I.; Nwankwo, A.C.N.; Spiegel, R.; Miller, B.R. First record of breeding populations of Aedes albopictus in continental Africa: Implications for arboviral transmission. J. Am. Mosq. Control Assoc. 1992, 8, 101–103. [Google Scholar] [PubMed]
- Fontenille, D.; Toto, J.C. Aedes (Stegomyia) albopictus (Skuse), a potential new Dengue vector in southern Cameroon. Emerg. Infect. Dis. 2001, 7, 1066–1067. [Google Scholar] [CrossRef]
- Ngoagouni, C.; Kamgang, B.; Nakoune, E.; Paupy, C.; Kazanji, M. Invasion of Aedes albopictus (Diptera: Culicidae) into central Africa: What consequences for emerging diseases? Parasites Vectors 2015, 8, 191. [Google Scholar] [CrossRef]
- Paupy, C.; Ollomo, B.; Kamgang, B.; Moutailler, S.; Rousset, D.; Demanou, M.; Hervé, J.; Leroy, E.; Simard, F. Comparative role of Aedes albopictus and Aedes aegypti in the emergence of dengue and chikungunya in central Africa. Vector Borne Zoonotic Dis. 2010, 10, 259–266. [Google Scholar] [CrossRef]
- Yadouleton, A.; Hounkanrin, G.; Tchibozo, C.; Bialonski, A.; Schmidt-Chanasit, J.; Jöst, H. First Detection of the Invasive Mosquito Vector Aedes albopictus (Diptera: Culicidae) in Benin, West Africa, 2021. J. Med. Entomol. 2022, 59, 1090–1094. [Google Scholar] [CrossRef]
- Gautret, P.; Botelho-Nevers, E.; Charrel, R.N.; Parola, P. Dengue virus infections in travellers returning from Benin to France, July-August 2010. Eurosurveillance 2010, 15, 36. [Google Scholar] [CrossRef]
- Eckerle, I.; Kapaun, A.; Junghanss, T.; Schnitzler, P.; Drosten, C.; Janisch, T. Dengue virus serotype 3 infection in traveler returning from West Africa to Germany. Emerg. Infect. Dis. 2015, 21, 175–177. [Google Scholar] [CrossRef]
- Allanonto, V.; Yanogo, P.; Sawadogo, B.; Akpo, Y.; Noudeke, D.N.; Saka, B.; Sourakatou, S. Investigation of dengue cases in the Atlantic, Littoral and Ouémé departments, Benin, April-July 2019. J. Interval Epidemiol. Public Health 2021, 3, 5. [Google Scholar]
- Padonou, G.G.; Ossè, R.; Salako, A.S.; Aikpon, R.; Sovi, A.; Kpanou, C.; Sagbohan, H.; Akadiri, Y.; Lamine, B.M.; Akogbeto, M.C. Entomological assessment of the risk of dengue outbreak in Abomey-Calavi Commune, Benin. Trop. Med. Health 2020, 48, 20. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Yellow Fever—African Region (AFRO). 2023. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON431 (accessed on 29 August 2023).
- Tchibozo, C.; Hounkanrin, G.; Yadouleton, A.; Bialonski, A.; Agboli, E.; Lühken, R.; Schmidt-Chanasit, J.; Jöst, H. Surveillance of arthropod-borne viruses in Benin, West Africa 2020–2021: Detection of dengue virus 3 in Aedes aegypti (Diptera: Culicidae). Mil. Med. Res. 2022, 9, 64. [Google Scholar] [CrossRef]
- Dieng, I.; Barry, M.A.; Talla, C.; Sow, B.; Faye, O.; Diagne, M.M.; Sall, A.A.; Falll, G.; Faye, O. Analysis of a Dengue Virus Outbreak in Rosso, Senegal 2021. Trop. Med. Infect. Dis. 2022, 7, 420. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Dengue Worldwide Overview. 2022. Available online: https://www.ecdc.europa.eu/en/dengue-monthly (accessed on 8 August 2023).
- Miller, E.; Huppert, A. The Effects of Host Diversity on Vector-Borne Disease: The Conditions under Which Diversity Will Amplify or Dilute the Disease Risk. PLoS ONE 2013, 8, e80279. [Google Scholar] [CrossRef] [PubMed]
- Jentes, E.S.; Poumerol, G.; Gershman, M.D.; Hill, D.R.; Lemarchand, J.; Lewis, R.F.; Staples, J.E.; Tomoro, O.; Wilder-Smith, A.; Monath, T.P. The revised global yellow fever risk map and recommendations for vaccination, 2010: Consensus of the WHO Informal Working Group on Geographical Risk of Yellow Fever. Lancet Infect. Dis. 2011, 11, 622–632. [Google Scholar] [CrossRef]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Gnanglè, C.P.; Kakaï, R.G.; Assogbadjo, A.E.; Vodounnon, S.; Yabi, J.A.; Sokpon, N. Tendances climatiques passées, modélisation, perceptions et adaptations locales au Bénin. Climatologie 2011, 8, 27–40. [Google Scholar] [CrossRef]
- Ministère de l’Environnement, de l’Habitat et de l’Urbanisme. Rapport National Sur la Diversité Biologique au Bénin; Ministère de l’Environnement, de l’Habitat et de l’Urbanisme: Cotonou, Benin, 1998; 69p. Available online: https://www.cbd.int/doc/world/bj/bj-nr-01-fr.pdf (accessed on 8 August 2023).
- Edwards, F.W. Mosquitoes of the Ethiopian Region. III. Culicine Adults and Pupae; OUP: Oxford, UK, 1941. [Google Scholar]
- Yiau-Min, H. The subgenus Stegomyia of Aedes in the Afrotropical Region with keys to the species (Diptera: Culicidae). Zootaxa 2004, 700, 1–120. [Google Scholar]
- Linton, Y.-M.; Harbach, R.E.; Chang, M.S.; Anthony, T.G.; Matusop, A. Morphological and molecular identity of Anopheles (Cellia) sundaicus (Diptera: Culicidae), the nominotypical member of a malaria vector species complex in Southeast Asia. Syst. Entomol. 2001, 26, 357–366. [Google Scholar] [CrossRef]
- Beckingham, K. Insect rDNA. In The Cell Nucleus, Part A; Busch, H., Rothblum, L., Eds.; Academic Press: New York, NY, USA, 1982; pp. 205–269. [Google Scholar]
- Collins, F.H.; Paskewitz, S.M. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Mol. Biol. 1996, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.H.; Collins, F.H. Species-diagnostic differences in a ribosomal DNA internal transcribed spacer from the sibling species Anopheles freeborni and Anopheles hermsi (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 1991, 45, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Wesson, L.; Eisenberg, D. Atomic solvation parameters applied to molecular dynamics of proteins in solution. Protein Sci. 1992, 1, 227–235. [Google Scholar] [CrossRef]
- Kjer, K.M.; Baldridge, G.D.; Fallon, A.M. Mosquito large subunit ribosomal RNA: Simultaneous alignment of primary and secondary structure. Biochim. Biophys. Acta 1994, 1217, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; CSHL: New York, NY, USA, 1989. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 8 August 2023).
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Rothman, K.J. Epidemiology: An Introduction; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Knudsen, A.B. Global distribution and continuing spread of Aedes albopictus. Parassitol 1995, 37, 91–97. [Google Scholar]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Bortel, W.V. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef]
- Anges, Y.; Gado, D.; Ahadji-Dabla, K.M.; Ramziyath, A.; Carine, T.; Achaz, A.; Roland, T.; Lamine, B.M. Evaluation du comportement trophique de Aedes aegypti dans la ville de cotonou au sud du Bénin. Eur. Sci. J. 2018, 14, 70–79. [Google Scholar] [CrossRef]
- Djènontin, A.; Bio-Bangana, S.; Moiroux, N.; Henry, M.C.; Bousari, O.; Chabi, J.; Ossè, R.; Koudénoukpo, S.; Corbel, V.; Akogbéto, M.; et al. Culicidae diversity, malaria transmission and insecticide resistance alleles in malaria vectors in Ouidah-Kpomasse-Tori district from Benin (West Africa): A pre-intervention study. Parasites Vectors 2010, 3, 83. [Google Scholar] [CrossRef]
- Ndille, E.E.; Doucoure, S.; Damien, G.; Mouchet, F.; Drame, P.M.; Cornelie, S.; Noukpo, H.; Yamadjako, S.; Djenontin, A.; Moiroux, N.; et al. First Attempt to Validate Human IgG Antibody Response to Nterm-34kDa Salivary Peptide as Biomarker for Evaluating Exposure to Aedes aegypti Bites. PLoS Negl. Trop. Dis. 2012, 6, e1905. [Google Scholar]
- Captain-Esoah, M.; Baidoo, P.K.; Frempong, K.K.; Adabie-Gomez, D.; Chabi, J.; Obuobi, D.; Amlalo, G.K.; Veriegh, F.B.; Donkor, M.; Asoala, V.; et al. Biting Behavior and Molecular Identification of Aedes aegypti (Diptera: Culicidae) Subspecies in Some Selected Recent Yellow Fever Outbreak Communities in Northern Ghana. J. Med. Entomol. 2020, 57, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Kampango, A.; Furu, P.; Sarath, D.L.; Haji, K.A.; Konradsen, F.; Schiøler, K.L.; Alifrangis, M.; Saleh, F.; Weldon, C.W. Risk factors for occurrence and abundance of Aedes aegypti and Aedes bromeliae at hotel compounds in Zanzibar. Parasites Vectors 2021, 14, 544. [Google Scholar] [CrossRef] [PubMed]
- Braks, M.A.; Honorio, N.A.; Lourencqo-De-Oliveira, R.; Juliano, S.A.; Lounibos, L.P. Convergent habitat segregation of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in southeastern Brazil and Florida. J. Med. Entomol. 2003, 40, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.D.; Nazni, W.A.; Lee, H.L.; Seleena, B.; Masri, S.M.; Chiang, Y.F.; Sofian-Azirun, M. Mixed breeding of Aedes aegypti (L.) and Aedes albopictus Skuse in four dengue endemic areas in Kuala Lumpur and Selangor, Malaysia. Trop. Biomed. 2006, 23, 224–227. [Google Scholar] [PubMed]
- Chukwuekezie, O.C.; Nwankwo, A.C.; Nwosu, E.O. Diversity and distribution of Aedes mosquitoes in Nigeria. N. Y. Sci. J. 2018, 11, 50–57. [Google Scholar]
- Braks, M.A.H.; Honorio, N.A.; Lounibos, L.P.; Lourenco-De-Oliveira, R.; Juliano, S.A. Interspecific competition between two invasive species of container mosquitoes, Aedes aegypti and Aedes albopictus (Diptera: Culicidae), in Brazil. Ann. Entomol. Soc. Am. 2004, 97, 130–139. [Google Scholar] [CrossRef]
- Camara, D.C.; Codeço, C.T.; Juliano, S.A.; Lounibos, L.P.; Riback, T.I.; Pereira, G.R.; Honorio, N.A. Seasonal Differences in Density but Similar Competitive Impact of Aedes albopictus (Skuse) on Aedes aegypti (L.) in Rio de Janeiro, Brazil. PLoS ONE 2016, 11, e0157120. [Google Scholar] [CrossRef]
- Egid, B.R.; Coulibaly, M.; Dadzie, S.K.; Kamgang, B.; McCall, P.J.; Sedda, L.; Toe, K.H.; Wilson, A.L. Review of the ecology and behaviour of Aedes aegypti and Aedes albopictus in Western Africa and implications for vector control. Curr. Res. Parasitol. Vector Borne Dis. 2022, 2, 100074. [Google Scholar] [CrossRef]
- Juliano, S.A.; Lounibos, L.P. Ecology of invasive mosquitoes: Effects on resident species and on human health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [CrossRef]
- Forattini, O.P. Culicideologia Médica—Identificação, Biologiae Epidemiologia; Edusp: São Paulo, Brazil, 2002; Volume 2. [Google Scholar]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; Evans, B.R.; Zheng, W.; Obas, V.; Barrera-Martinez, L.; Egizi, A.; Zhao, H.; Caccone, A.; Powell, J.R. Human impacts have shaped historical and recent evolution in Aedes aegypti, the dengue and yellow fever mosquito. Evolution 2013, 68, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Kamgang, B.; Happi, J.Y.; Boisier, P.; Njiokou, F.; Herve, J.P.; Simard, F.; Paupy, C. Geographic and ecological distribution of the dengue and chikungunya virus vectors Aedes aegypti and Aedes albopictus in three major Cameroonian towns. Med. Vet. Entomol. 2010, 24, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Adeleke, M.A.; Sam-Wobo, S.O.; Garza-Hernandez, J.A.; Oluwole, A.S.; Mafiana, C.F.; Reyes-Villanueva, F.; Rodriguez-Perez, M.A.; Mafiana, C.F. Twenty-three years after the first record of Aedes albopictus in Nigeria: Its current distribution and potential epidemiological implications. Afr. Entomol. 2015, 23, 348–355. [Google Scholar] [CrossRef]
- Bennett, K.L.; Gómez, C.; Almanza, A.; Rovira, J.R.; McMillan, W.O.; Enriquez, V.; Barraza, E.; Diaz, M.; Sanchez-Galan, J.E.; Whiteman, A.; et al. High infestation of invasive Aedes mosquitoes in used tires along the local transport network of Panama. Parasites Vectors 2019, 12, 264. [Google Scholar] [CrossRef]
- Farjana, T.; Tuno, N.; Higa, Y. Effects of temperature and diet on development and interspecies competition in Aedes aegypti and Aedes albopictus. Med. Vet. Entomol. 2012, 26, 210–217. [Google Scholar] [CrossRef]
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Ecologia 2002, 130, 458–469. [Google Scholar] [CrossRef]
- Tedjou, A.N.; Kamgang, B.; Yougang, A.P.; Njiokou, F.; Wondji, C.S. Update on the geographical distribution and prevalence of Aedes aegypti and Aedes albopictus (Diptera: Culicidae), two major arbovirus vectors in Cameroon. PLoS Negl. Trop. Dis. 2019, 13, e0007137. [Google Scholar] [CrossRef]
- Humphrey, J.M.; Cleton, N.B.; Reusken, C.B.E.M.; Glesby, M.J.; Koopmans, M.P.G.; Abu-Raddad, L.J. Dengue in the Middle East and North Africa: A systematic review. PLoS Negl. Trop. Dis. 2016, 10, e0005194. [Google Scholar] [CrossRef]
- Khan, J.; Khan, I.; Amin, I. A comprehensive entomological, serological and molecular study of 2013 dengue outbreak of swat, Khyber Pakhtunkhwa, Pakistan. PLoS ONE 2016, 11, e0147416. [Google Scholar] [CrossRef]
- Nascimento, K.L.C.; da Silva, J.F.M.; Zequi, J.A.C.; Lopes, J. Comparison Between Larval Survey Index and Positive Ovitrap Index in the Evaluation of Populations of Aedes (Stegomyia) aegypti (Linnaeus, 1762) North of Paraná, Brazil. Environ. Health Insights 2020, 14, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Okoye, C.F.; Onyido, A.E.; Chikwendu, J.I. Abundance of mosquito vectors of human diseases at the Awka campus of Nnamdi Azikiwe University, Awka Anambra state Nigeria. Microbes Infect. 2023, 4, 259–267. [Google Scholar] [CrossRef]
- Reinhold, J.M.; Lazzari, C.R.; Lahondère, C. Effects of the environmental temperature on Aedes aegypti and Aedes albopictus mosquitoes: A review. Insects 2018, 9, 158. [Google Scholar] [CrossRef]
- Diarrassouba, S.; Dossou-Yovo, J. Atypical activity rhythm in Aedes aegypti in a sub-sudanian savannah zone of Côte dʼIvoire. Bull. Soc. Pathol. Exot. 1997, 90, 361–363. (In French) [Google Scholar] [PubMed]
- Labbo, R.; Doumma, A.; Mahamadou, I.; Arzika, I.; Soumana, A.; Kadri, S.; Idi, I.; Testa, J. Distribution and relative densities of Aedes aegypti in Niger. Med. Sante Trop. 2019, 29, 47–54. [Google Scholar]
- Futami, K.; Valderrama, A.; Baldi, M.; Minakawa, N.; Rodríguez, R.M.; Chaves, L.F. New and Common Haplotypes Shape Genetic Diversity in Asian Tiger Mosquito Populations from Costa Rica and Panamá. J. Econ. Entomol. 2015, 108, 761–768. [Google Scholar] [CrossRef]
- Bowman, L.R.; Runge-Ranzinger, S.; McCall, P.J. Assessing the relationship between vector indices and dengue transmission: A systematic review of the evidence. PLoS Negl. Trop. Dis. 2014, 8, e2848. [Google Scholar] [CrossRef]
- Sanchez, L.; Vanlerberghe, V.; Alfonso, L.; Marquetti, M.C.; Guzman, M.G.; Bisset, J.; van der Stuyft, P. Aedes aegypti larval indices and risk for dengue epidemics. Emerg. Infect. Dis. 2006, 12, 800–806. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Localities (Communes) | Ae. aegypti | Ae. albopictus | Ae. africanus | Ae. vitatus | Ae. (Neomelaniconion) palpalis | Ae. luteocephalus | Total | Taxa (S) |
|---|---|---|---|---|---|---|---|---|
| ni (Pi) | ni (Pi) | ni (Pi) | ni (Pi) | ni (Pi) | ni (Pi) | N | ||
| Ifangni | 1964 (47.83) | 1731 (42.16) | 0 | 0 | 1 (0.02) | 410 (9.99) | 4106 | 4 |
| Kétou | 139 (82.74) | 22 (13.10) | 1 (0.60) | 0 | 3 (1.79) | 3 (1.79) | 168 | 5 |
| Sakété | 128 (55.41) | 44 (19.05) | 8 (3.46) | 2 (0.87) | 49 (21.21) | 0 | 231 | 5 |
| Pobè | 771 (87.71) | 92 (10.47) | 0 | 0 | 0 | 16 (1.82) | 879 | 3 |
| Adja-Ouèrè | 482 (87.96) | 66 (12.04) | 0 | 0 | 0 | 0 | 548 | 2 |
| Porto-Novo | 1653 (92.14) | 133 (7.41) | 0 | 0 | 0 | 4 (0.22) | 1790 | 3 |
| Adjara | 128 (85.33) | 22 (14.67) | 0 | 0 | 0 | 0 | 150 | 2 |
| Avrankou | 178 (46.97) | 196 (51.72) | 0 | 2 (0.53) | 3 (0.79) | 0 | 379 | 4 |
| Abomey–Calavi | 181 (87.44) | 26 (12.56) | 0 | 0 | 0 | 0 | 207 | 2 |
| Bohicon | 252 (89.68) | 29 (10.32) | 0 | 0 | 0 | 0 | 281 | 2 |
| Zangnanado | 189 (96.92) | 6 (3.08) | 0 | 0 | 0 | 0 | 195 | 2 |
| Lokossa | 200 (100) | 0 | 0 | 0 | 0 | 0 | 200 | 1 |
| Klouekanmè | 251 (100) | 0 | 0 | 0 | 0 | 0 | 251 | 1 |
| Grand-Popo | 600 (100) | 0 | 0 | 0 | 0 | 0 | 600 | 1 |
| Djougou | 241 (100) | 0 | 0 | 0 | 0 | 0 | 241 | 1 |
| Copargo | 301 (100) | 0 | 0 | 0 | 0 | 0 | 301 | 1 |
| Savè | 114 (100) | 0 | 0 | 0 | 0 | 0 | 114 | 1 |
| Parakou | 314 (98.74) | 0 | 0 | 4 (1.25) | 0 | 0 | 318 | 2 |
| Nikki | 152 (98.70) | 0 | 0 | 1 (0.65) | 0 | 1 (0.65) | 154 | 3 |
| Bantè | 551 (100) | 0 | 0 | 0 | 0 | 0 | 551 | 1 |
| Dassa | 206 (86.19) | 33 (13.81) | 0 | 0 | 0 | 0 | 239 | 2 |
| Ségbana | 210 (99.52) | 0 | 0 | 1 (0.47) | 0 | 0 | 211 | 2 |
| Gogounou | 303 (97.74) | 0 | 0 | 6 (1.93) | 0 | 1 (0.32) | 310 | 3 |
| Total | 9508 | 2400 | 9 | 16 | 56 | 435 | 12,424 | - |
| Proportion (Pi) (%) [95% CI] | 76.53 [75.77–77.27] | 19.32 [18.63–20.02] | 0.07 [0.03–0.14] | 0.13 [0.07–0.21] | 0.45 [0.34–0.58] | 3.50 [3.18–3.84] | - | - |
| Eco-Geographical Zones | Total | Taxas (S) | Equitability (J) [IC-95%] | Shannon (H) [IC-95%] |
|---|---|---|---|---|
| Subequatorial climate (Oueme, Plateau, Atlantic, Mono, Couffo and Zou regions) | 9985 | 6 | 0.42 [0.41–0.43] | 0.76 [0.74–0.77] |
| Sudanese-Guinean climate (Collines, Donga and Borgou regions) | 1918 | 4 | 0.08 [0.05–0.10] | 0.11 [0.08–0.14] |
| Sudanese climate (Alibori region) | 521 | 3 | 0.07 [0.03–0.012] | 0.08 [0.03–0.13] |
| Eco-Climatic Zones | Ae. aegypti | Ae. albopictus | Ae. luteocephalus | Others Aedes | Total |
|---|---|---|---|---|---|
| N (%) | N (%) | N (%) | N (%) | ||
| Subequatorial climate (Oueme, Plateau, Atlantic, Mono, Couffo and Zou regions) | 7116 (71.27) | 2367 (23.71) | 433 (4.34) | 69 (4.34) | 9985 |
| Sudanese-Guinean climate (Collines, Donga and Borgou regions) | 1879 (97.97) | 33 (1.72) | 1 (0.05) | 5 (0.26) | 1918 |
| Sudanese climate (Alibori region) | 513 (98.46) | 0 | 1 (0.19) | 7 (1.34) | 521 |
| Climates and Aedes Species | Human Landing Catch (HLC) | Larvae and Pupae Collection | Ovitrap Method | Total |
|---|---|---|---|---|
| n (%) | n (%) | n (%) | N | |
| Subequatorial climate | ||||
| Ae. aegypti | 1398 (19.65) | 151 (2.12) | 5567 (78.23) | 7116 |
| Ae. albopictus | 131 (5.53) | 54 (2.28) | 2182 (92.18) | 2367 |
| Ae. luteocephalus | 4 (0.92) | 0 (0.00) | 429 (99.08) | 433 |
| Ae. palpalis | 56 (100.00) | 0 (0.00) | 0 (0.00) | 56 |
| Ae. africanus | 3 (33.33) | 6 (66.67) | 0 (0.00) | 9 |
| Ae. vitatus | 4 (100.00) | 0 (0.00) | 0 (0.00) | 4 |
| Sudano-Guinean climate | ||||
| Ae. aegypti | 1480 (78.77) | 348 (18.52) | 51 (2.71) | 1879 |
| Ae. albopictus | 33 (100.00) | 0 (0.00) | 0 (0.00) | 33 |
| Ae. vitatus | 4 (80.00) | 0 (0.00) | 1 (20.00) | 5 |
| Ae. luteocephalus | 0 (0.00) | 1 (100.00) | 0 (0.00) | 1 |
| Sudanian climate | ||||
| Ae. aegypti | 253 (49.32) | 197 (38.40) | 63 (12.28) | 513 |
| Ae. vitatus | 2 (28.57) | 5 (71.43) | 0 (0.00) | 7 |
| Ae. luteocephalus | 0 (0.00) | 0 (0.00) | 1 (100.00) | 1 |
| All areas | ||||
| Ae. aegypti | 3131 (32.93) | 688 (7.32) | 5681 (59.75) | 9508 |
| Ae. albopictus | 164 (6.83) | 54 (2.25) | 2182 (90.92) | 2400 |
| Ae. luteocephalus | 4 (0.92) | 1 (0.23) | 430 (98.85) | 435 |
| Ae. palpalis | 56 (100.00) | 0 (0.00) | 0 (0.00) | 56 |
| Ae. africanus | 3 (33.33) | 6 (66.67) | 0 (0.00) | 9 |
| Ae. vitatus | 10 (62.50) | 5 (31.25) | 1 (6.25) | 16 |
| Total | 3368 (27.11) | 762 (6.13) | 8294 (66.75) | 12,424 |
| Localities (Communes) | N-Tested | N-Confirmed |
|---|---|---|
| Abomey-Calavi | 26 | 26 |
| Adjarra | 22 | 22 |
| Avrankou | 30 | 30 |
| Ifangni | 30 | 30 |
| Kétou | 22 | 22 |
| Porto-Novo | 30 | 30 |
| Bohicon | 29 | 29 |
| Adja-ouèrè | 30 | 30 |
| Dassa | 30 | 30 |
| Pobè | 30 | 30 |
| Zangnanado | 6 | 6 |
| Sakété | 30 | 30 |
| Total | 315 | 315 |
| Localities (Communes) | Periods | Nb Ae. aegypti Collected | HBR/Day | CI-95% | |||||
|---|---|---|---|---|---|---|---|---|---|
| Indoors | Outdoors | Total | Indoors | Outdoors | RR and CI-95% | Total HBR /Day | |||
| Porto-Novo | May 2021 | 41 | 70 | 111 | 5.13 a | 8.75 b | 0.58 [0.38–0.87] | 6.94 | [5.71–8.35] |
| July 2021 | 5 | 145 | 150 | 0.63 a | 18.13 b | 0.03 [0.01–0.08] | 9.38 | [7.93–11.00] | |
| October 2022 | 9 | 22 | 31 | 1.13 a | 2.75 b | 0.4 [0.16–0.92] | 1.94 | [1.32–2.75] | |
| Total | Rainy season (May 2021–October 2022) | 55 | 237 | 292 | 2.29 a | 9.88 b | 0.23 [0.17–0.31] | 6.08 | [5.41–6.82] |
| Ifangni | May 2021 | 7 | 49 | 56 | 0.88 a | 6.13 b | 0.14 [0.05–0.32] | 3.5 | [2.64–4.54] |
| July 2021 | 13 | 40 | 53 | 1.63 a | 5 b | 0.32 [0.16–0.62] | 3.31 | [2.48–4.33] | |
| Total | Rainy season (May–July 2021) | 20 | 89 | 109 | 1.25 a | 5.56 b | 0.22 [0.13–0.37] | 3.41 | [2.80–4.11] |
| Kétou | Dry season (August 2022) | 2 | 20 | 22 | 0.25 a | 2.5 b | 0.1 [0.01–0.41] | 1.38 | [0.86–2.08] |
| Avrankou | Dry season (August 2022) | 1 | 2 | 3 | 0.13 a | 0.25 a | 0.5 [0.00–9.6] | 0.19 | [0.038–0.55] |
| Overall total | Dry and rainy seasons (May 2021–August 2022) | 153 | 674 | 827 | 1.59 a | 7.02 b | 0.23 [0.19–0.27] | 4.31 | [4.02–4.61] |
| Localities (Communes) | Periods | Nb Ae. albopictus Collected | HBR/Day | CI-95% | |||||
|---|---|---|---|---|---|---|---|---|---|
| Indoors | Outdoors | Total | Indoors | Outdoors | RR and CI-95% | Total HBR /Day | |||
| Porto-Novo | May 2021 | 2 | 5 | 7 | 0.25 a | 0.63 a | 0.4 [0.04–2.44] | 0.44 | [0.17–0.90] |
| July 2021 | 1 | 5 | 6 | 0.13 a | 0.63 b | 0.2 [0.004–1.79] | 0.38 | [0.14–0.82] | |
| October 2022 | 2 | 13 | 15 | 0.25 a | 1.63 a | 0.15 [0.02–0.68] | 0.94 | [0.52–1.54] | |
| Total | Rainy season (May 2021–October 2022) | 5 | 23 | 28 | 0.21 a | 0.96 b | 0.22 [0.06–0.58] | 0.58 | [0.39–0.84] |
| Ifangni | May 2021 | 6 | 15 | 21 | 0.75 a | 1.87 a | 0.4 [0.13–1.09] | 1.31 | [0.81–2.00] |
| July 2021 | 5 | 9 | 14 | 0.63 a | 1.13 a | 0.55 [0.15–1.84] | 0.88 | [0.48–1.47] | |
| Total | Rainy season (May–July 2021) | 11 | 24 | 35 | 0.69 a | 1.50 b | 0.46 [0.20–0.97] | 1.09 | [0.76–1.52] |
| Ketou | Dry season (August 2022) | 0 | 12 | 12 | 0.00 a | 1.50 b | 0 [0.00–0.36] | 0.75 | [0.39–1.31] |
| Avrankou | Dry season (August 2022) | 5 | 39 | 44 | 0.63 a | 4.88 b | 0.13 [0.04–0.32] | 2.75 | [2.00–3.69] |
| Overall total | Dry and rainy seasons (May 2021–August 2022) | 37 | 145 | 182 | 0.39 a | 1.51 b | 0.25 [0.17–0.37] | 0.95 | [0.81–1.09] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padonou, G.G.; Konkon, A.K.; Salako, A.S.; Zoungbédji, D.M.; Ossè, R.; Sovi, A.; Azondekon, R.; Sidick, A.; Ahouandjinou, J.M.; Adoha, C.J.; et al. Distribution and Abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Benin, West Africa. Trop. Med. Infect. Dis. 2023, 8, 439. https://doi.org/10.3390/tropicalmed8090439
Padonou GG, Konkon AK, Salako AS, Zoungbédji DM, Ossè R, Sovi A, Azondekon R, Sidick A, Ahouandjinou JM, Adoha CJ, et al. Distribution and Abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Benin, West Africa. Tropical Medicine and Infectious Disease. 2023; 8(9):439. https://doi.org/10.3390/tropicalmed8090439
Chicago/Turabian StylePadonou, Germain Gil, Alphonse Keller Konkon, Albert Sourou Salako, David Mahouton Zoungbédji, Razaki Ossè, Arthur Sovi, Roseric Azondekon, Aboubakar Sidick, Juvénal Minassou Ahouandjinou, Constantin Jesukèdè Adoha, and et al. 2023. "Distribution and Abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Benin, West Africa" Tropical Medicine and Infectious Disease 8, no. 9: 439. https://doi.org/10.3390/tropicalmed8090439