
Chikungunya Virus Diagnosis: A Review of Current Antigen Detection Methods


Abstract
:1. Introduction
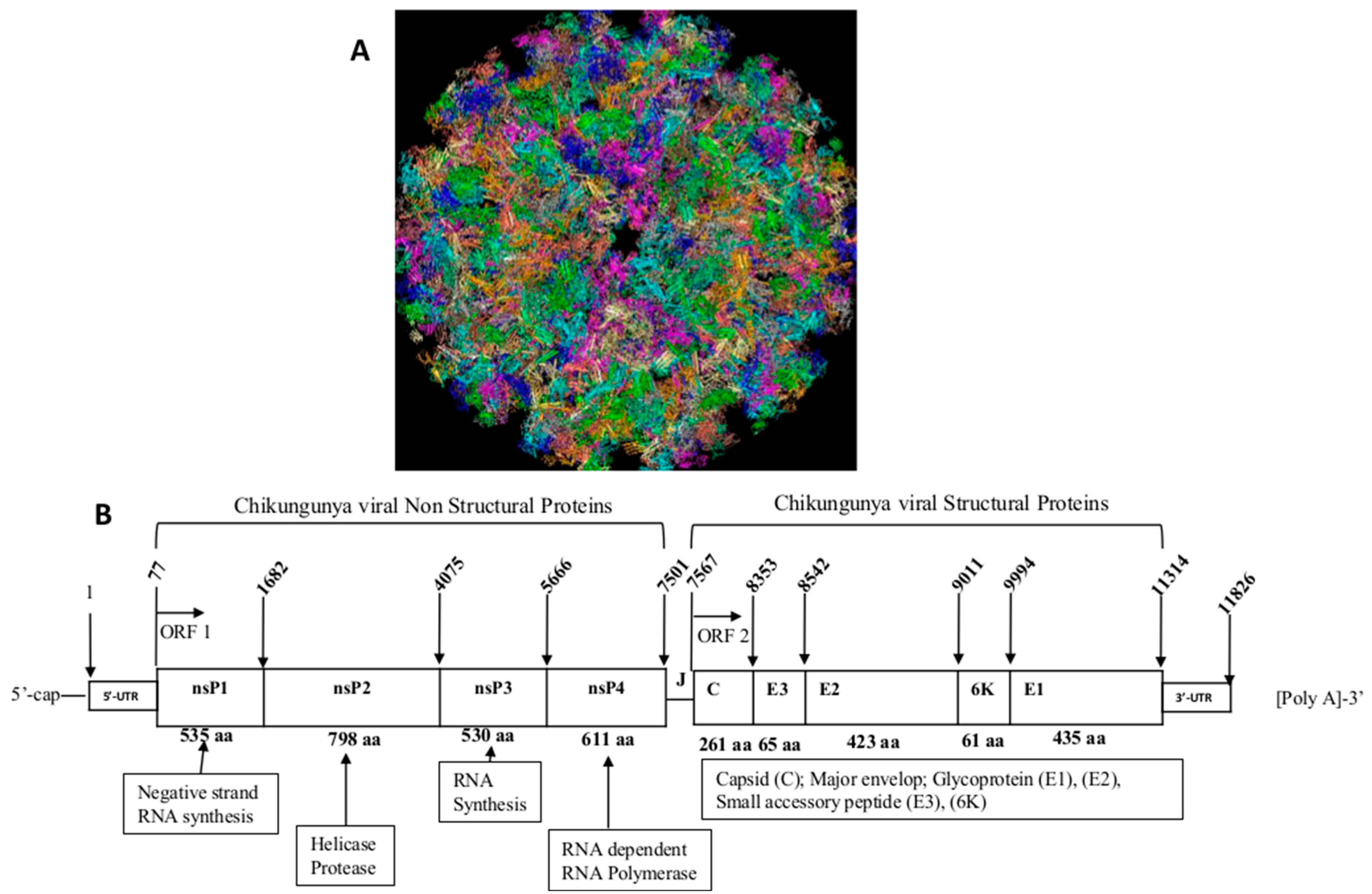
2. Physiopathology of Chikungunya Virus Infection
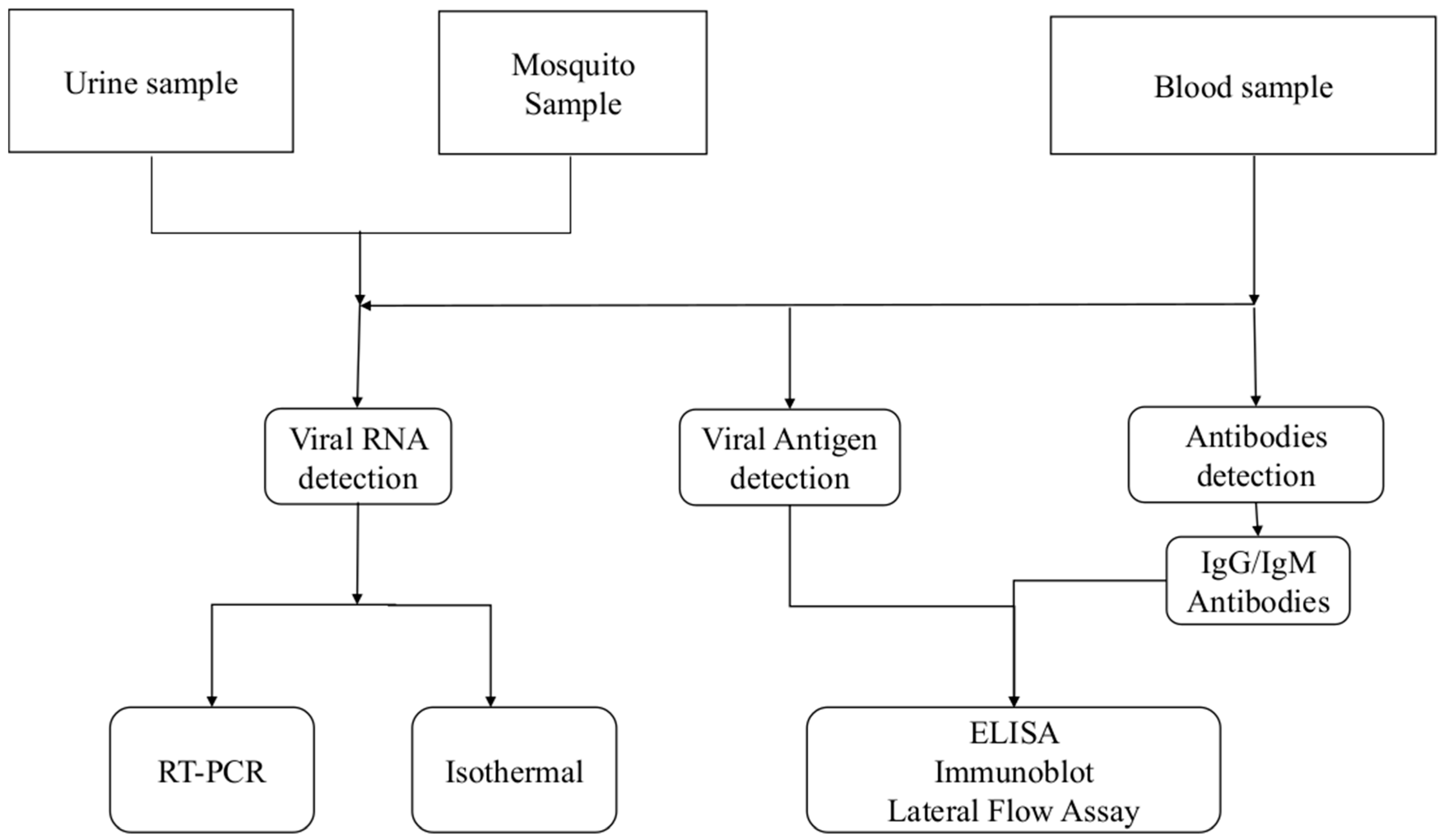
3. Diagnosis of Chikungunya Virus Infection
4. Chikungunya Detection Using Viral Antigen E1 and E2 as a Target
5. Future Perspectives on Chikungunya Diagnostics
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Burt, F.J.; Chen, W.; Miner, J.J.; Lenschow, D.J.; Merits, A.; Schnettler, E.; Kohl, A.; Rudd, P.A.; Taylor, A.; Herrero, L.J.; et al. Chikungunya Virus: An Update on the Biology and Pathogenesis of This Emerging Pathogen. Lancet Infect. Dis. 2017, 17, e107–e117. [Google Scholar] [CrossRef] [PubMed]
- Khongwichit, S.; Chansaenroj, J.; Chirathaworn, C.; Poovorawan, Y. Chikungunya Virus Infection: Molecular Biology, Clinical Characteristics, and Epidemiology in Asian Countries. J. Biomed. Sci. 2021, 28, 84. [Google Scholar] [CrossRef] [PubMed]
- Caglioti, C.; Lalle, E.; Castilletti, C.; Carletti, F.; Capobianchi, M.R.; Bordi, L. Chikungunya Virus Infection: An Overview. N. Microbiol. 2013, 36, 211–227. [Google Scholar]
- Moizéis, R.N.C.; de Medeiros Fernandes, T.A.A.; Guedes, P.M.d.M.; Pereira, H.W.B.; Lanza, D.C.F.; de Azevedo, J.W.V.; Galvão, J.M.d.A.; Fernandes, J.V. Chikungunya Fever: A Threat to Global Public Health. Pathog. Glob. Health 2018, 112, 182–194. [Google Scholar] [CrossRef]
- Soumahoro, M.-K.; Boelle, P.-Y.; Gaüzere, B.-A.; Atsou, K.; Pelat, C.; Lambert, B.; Ruche, G.L.; Gastellu-Etchegorry, M.; Renault, P.; Sarazin, M.; et al. The Chikungunya Epidemic on La Réunion Island in 2005–2006: A Cost-of-Illness Study. PLoS Negl. Trop. Dis. 2011, 5, e1197. [Google Scholar] [CrossRef] [Green Version]
- Simo, F.B.N.; Bigna, J.J.; Well, E.A.; Kenmoe, S.; Sado, F.B.Y.; Weaver, S.C.; Moundipa, P.F.; Demanou, M. Chikungunya Virus Infection Prevalence in Africa: A Contemporaneous Systematic Review and Meta-Analysis. Public Health 2019, 166, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Reyes, E.; Núñez-Avellaneda, D.; Alonso Palomares, L.A.; Salazar, M.I. Chikungunya: Molecular Aspects, Clinical Outcomes and Pathogenesis. Rev. Investig. Clin. Organo Hosp. Enferm. Nutr. 2017, 69, 299–307. [Google Scholar] [CrossRef]
- Kendall, C.; Khalid, H.; Müller, M.; Banda, D.H.; Kohl, A.; Merits, A.; Stonehouse, N.J.; Tuplin, A. Structural and Phenotypic Analysis of Chikungunya Virus RNA Replication Elements. Nucleic Acids Res. 2019, 47, 9296–9312. [Google Scholar] [CrossRef] [Green Version]
- Metz, S.W.; Geertsema, C.; Martina, B.E.; Andrade, P.; Heldens, J.G.; van Oers, M.M.; Goldbach, R.W.; Vlak, J.M.; Pijlman, G.P. Functional Processing and Secretion of Chikungunya Virus E1 and E2 Glycoproteins in Insect Cells. Virol. J. 2011, 8, 353. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.M.; Long, F.; Edeling, M.A.; Lin, H.; van Duijl-Richter, M.K.S.; Fong, R.H.; Kahle, K.M.; Smit, J.M.; Jin, J.; Simmons, G.; et al. Broadly Neutralizing Alphavirus Antibodies Bind an Epitope on E2 and Inhibit Entry and Egress. Cell 2015, 163, 1095–1107. [Google Scholar] [CrossRef] [Green Version]
- Voss, J.E.; Vaney, M.-C.; Duquerroy, S.; Vonrhein, C.; Girard-Blanc, C.; Crublet, E.; Thompson, A.; Bricogne, G.; Rey, F.A. Glycoprotein Organization of Chikungunya Virus Particles Revealed by X-Ray Crystallography. Nature 2010, 468, 709–712. [Google Scholar] [CrossRef]
- Yap, M.L.; Klose, T.; Urakami, A.; Hasan, S.S.; Akahata, W.; Rossmann, M.G. Structural Studies of Chikungunya Virus Maturation. Proc. Natl. Acad. Sci. USA 2017, 114, 13703–13707. [Google Scholar] [CrossRef] [Green Version]
- Metz, S.W.H. Chikungunya Virus-Like Particle Vaccine. Wageningen University: Wageningen, The Netherlands, 2013; ISBN 978-94-6173-565-2. [Google Scholar]
- Reiter, P.; Fontenille, D.; Paupy, C. Aedes Albopictus as an Epidemic Vector of Chikungunya Virus: Another Emerging Problem? Lancet Infect. Dis. 2006, 6, 463–464. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, O.; Albert, M.L. Biology and Pathogenesis of Chikungunya Virus. Nat. Rev. Microbiol. 2010, 8, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Dubrulle, M.; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.-B. Chikungunya Virus and Aedes Mosquitoes: Saliva Is Infectious as Soon as Two Days after Oral Infection. PLoS ONE 2009, 4, e5895. [Google Scholar] [CrossRef] [PubMed]
- Solignat, M.; Gay, B.; Higgs, S.; Briant, L.; Devaux, C. Replication Cycle of Chikungunya: A Re-Emerging Arbovirus. Virology 2009, 393, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Rudd, P.A.; Wilson, J.; Gardner, J.; Larcher, T.; Babarit, C.; Le, T.T.; Anraku, I.; Kumagai, Y.; Loo, Y.-M.; Gale, M.; et al. Interferon Response Factors 3 and 7 Protect against Chikungunya Virus Hemorrhagic Fever and Shock. J. Virol. 2012, 86, 9888–9898. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ozden, S.; Huerre, M.; Riviere, J.-P.; Coffey, L.L.; Afonso, P.V.; Mouly, V.; de Monredon, J.; Roger, J.-C.; El Amrani, M.; et al. Human Muscle Satellite Cells as Targets of Chikungunya Virus Infection. PLoS ONE 2007, 2, e527. [Google Scholar] [CrossRef]
- Suganthini, K.N.; Chandrasekar, P.H. Chikungunya Virus: Background, Pathophysiology, Etiology. Medscape 2022, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Natrajan, M.S.; Rojas, A.; Waggoner, J.J. Beyond Fever and Pain: Diagnostic Methods for Chikungunya Virus. J. Clin. Microbiol. 2019, 57, e00350-19. [Google Scholar] [CrossRef] [Green Version]
- Islam, K.U.; Iqbal, J. An Update on Molecular Diagnostics for COVID-19. Front. Cell. Infect. Microbiol. 2020, 10, 560616. [Google Scholar] [CrossRef] [PubMed]
- Zamarina, T.V.; Pimenova, E.V.; Khrapova, N.P.; Baturin, A.A. Current State of Chikungunya Fever Laboratory Diagnosis (Review of Literature). Klin. Lab. Diagn. 2021, 66, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Burdino, E.; Calleri, G.; Caramello, P.; Ghisetti, V. Unmet Needs for a Rapid Diagnosis of Chikungunya Virus Infection. Emerg. Infect. Dis. 2016, 22, 1837–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, S.J.R.; Pardee, K.; Balasuriya, U.B.R.; Pena, L. Development and Validation of a One-Step Reverse Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) for Rapid Detection of ZIKV in Patient Samples from Brazil. Sci. Rep. 2021, 11, 4111. [Google Scholar] [CrossRef]
- Panning, M.; Charrel, R.N.; Mantke, O.D.; Landt, O.; Niedrig, M.; Drosten, C. Coordinated Implementation of Chikungunya Virus Reverse Transcription–PCR. Emerg. Infect. Dis. 2009, 15, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Azami, N.A.M.; Moi, M.L.; Takasaki, T. Neutralization Assay for Chikungunya Virus Infection: Plaque Reduction Neutralization Test. Methods Mol. Biol. 2016, 1426, 273–282. [Google Scholar] [CrossRef]
- Pellenz, S. An Introduction to ELISA (Part 1). Available online: https://www.antibodies-online.com/resources/17/1461/an-introduction-to-elisa-part-1/ (accessed on 28 June 2023).
- Lequin, R.M. Enzyme Immunoassay (EIA)/Enzyme-Linked Immunosorbent Assay (ELISA). Clin. Chem. 2005, 51, 2415–2418. [Google Scholar] [CrossRef] [Green Version]
- Im, K.; Mareninov, S.; Diaz, M.F.P.; Yong, W.H. An Introduction to Performing Immunofluorescence Staining. Methods Mol. Biol. 2019, 1897, 299–311. [Google Scholar] [CrossRef]
- Javelle, E.; Gautret, P.; Leparc-Goffart, I. Letter to the Editor: False-Positive Results with Rapid Diagnostic Tests (RDT) for Dengue. Euro Surveill. 2019, 24, 1900304. [Google Scholar] [CrossRef]
- Kashyap, R.S.; Morey, S.H.; Ramteke, S.S.; Chandak, N.H.; Parida, M.; Deshpande, P.S.; Purohit, H.J.; Taori, G.M.; Daginawala, H.F. Diagnosis of Chikungunya Fever in an Indian Population by an Indirect Enzyme-Linked Immunosorbent Assay Protocol Based on an Antigen Detection Assay: A Prospective Cohort Study. Clin. Vaccine Immunol. CVI 2010, 17, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Okabayashi, T.; Sasaki, T.; Masrinoul, P.; Chantawat, N.; Yoksan, S.; Nitatpattana, N.; Chusri, S.; Morales Vargas, R.E.; Grandadam, M.; Brey, P.T.; et al. Detection of Chikungunya Virus Antigen by a Novel Rapid Immunochromatographic Test. J. Clin. Microbiol. 2015, 53, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Jain, J.; Okabayashi, T.; Kaur, N.; Nakayama, E.; Shioda, T.; Gaind, R.; Kurosu, T.; Sunil, S. Evaluation of an Immunochromatography Rapid Diagnosis Kit for Detection of Chikungunya Virus Antigen in India, a Dengue-Endemic Country. Virol. J. 2018, 15, 84. [Google Scholar] [CrossRef] [PubMed]
- Huits, R.; Okabayashi, T.; Cnops, L.; Barbé, B.; Berg, R.V.D.; Bartholomeeusen, K.; Ariën, K.K.; Jacobs, J.; Bottieau, E.; Nakayama, E.E.; et al. Diagnostic Accuracy of a Rapid E1-Antigen Test for Chikungunya Virus Infection in a Reference Setting. Clin. Microbiol. Infect. 2018, 24, 78–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.C.; Park, H.; Shin, H.-J.; Nguyen, N.M.; Nguyen, A.T.V.; Trinh, T.-T.T.; Duong, T.H.Y.; Tuong, H.T.; Hoang, V.T.; Seo, G.-E.; et al. Fluorescent Immunosorbent Assay for Chikungunya Virus Detection. Intervirology 2019, 62, 145–155. [Google Scholar] [CrossRef]
- Suzuki, K.; Huits, R.; Phadungsombat, J.; Tuekprakhon, A.; Nakayama, E.E.; van den Berg, R.; Barbé, B.; Cnops, L.; Rahim, R.; Hasan, A.; et al. Promising Application of Monoclonal Antibody against Chikungunya Virus E1-Antigen across Genotypes in Immunochromatographic Rapid Diagnostic Tests. Virol. J. 2020, 17, 90. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.; Bosch, I.; Salcedo, N.; Herrera, B.B.; de Puig, H.; Narváez, C.F.; Caicedo-Borrero, D.M.; Lorenzana, I.; Parham, L.; García, K.; et al. Development and Validation of a Rapid Lateral Flow E1/E2-Antigen Test and ELISA in Patients Infected with Emerging Asian Strain of Chikungunya Virus in the Americas. Viruses 2020, 12, 971. [Google Scholar] [CrossRef] [PubMed]
- Chua, C.L.; Sam, I.-C.; Chan, Y.F. Expression and Purification of E2 Glycoprotein from Insect Cells (Sf9) for Use in Serology. Methods Mol. Biol. 2016, 1426, 51–61. [Google Scholar] [CrossRef]
- Khan, M.; Dhanwani, R.; Kumar, J.S.; Rao, P.V.L.; Parida, M. Comparative Evaluation of the Diagnostic Potential of Recombinant Envelope Proteins and Native Cell Culture Purified Viral Antigens of Chikungunya Virus. J. Med. Virol. 2014, 86, 1169–1175. [Google Scholar] [CrossRef]
- Weber, C.; König, R.; Niedrig, M.; Emmerich, P.; Schnierle, B.S. A Neutralization Assay for Chikungunya Virus Infections in a Multiplex Format. J. Virol. Methods 2014, 201, 7–12. [Google Scholar] [CrossRef]
- Holmes, A.C.; Basore, K.; Fremont, D.H.; Diamond, M.S. A Molecular Understanding of Alphavirus Entry. PLoS Pathog. 2020, 16, e1008876. [Google Scholar] [CrossRef]
- Kumar, P.; Pok, K.-Y.; Tan, L.-K.; Angela, C.; Leo, Y.-S.; Ng, L.-C. Development and Evaluation of Baculovirus-Expressed Chikungunya Virus E1 Envelope Proteins for Serodiagnosis of Chikungunya Infection. J. Virol. Methods 2014, 206, 67–75. [Google Scholar] [CrossRef]
- Bagno, F.F.; Godói, L.C.; Figueiredo, M.M.; Sérgio, S.A.R.; Moraes, T.d.F.S.; Salazar, N.d.C.; Kim, Y.C.; Reyes-Sandoval, A.; da Fonseca, F.G. Chikungunya E2 Protein Produced in E. Coli and HEK293-T Cells-Comparison of Their Performances in ELISA. Viruses 2020, 12, 939. [Google Scholar] [CrossRef] [PubMed]
- Shukla, M.; Chandley, P.; Tapryal, S.; Kumar, N.; Mukherjee, S.P.; Rohatgi, S. Expression, Purification, and Refolding of Chikungunya Virus Full-Length Envelope E2 Protein along with B-Cell and T-Cell Epitope Analyses Using Immuno-Informatics Approaches. ACS Omega 2022, 7, 3491–3513. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.K.; Priya, R.; Shrivastava, A. Production of Recombinant Chikungunya Virus Envelope 2 Protein in Escherichia Coli. Appl. Microbiol. Biotechnol. 2014, 98, 2461–2471. [Google Scholar] [CrossRef]
- Verma, A.; Chandele, A.; Nayak, K.; Kaja, M.K.; Arulandu, A.; Lodha, R.; Ray, P. High Yield Expression and Purification of Chikungunya Virus E2 Recombinant Protein and Its Evaluation for Serodiagnosis. J. Virol. Methods 2016, 235, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.; Chen, H.; Chen, C.K.; Min, N.; Chu, J.J.H. Antiviral Phosphorodiamidate Morpholino Oligomers Are Protective against Chikungunya Virus Infection on Cell-Based and Murine Models. Sci. Rep. 2015, 5, 12727. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yang, J.; Kim, Y.B.; Lee, H.-J.; Kim, S.; Poo, H. Development of a Specific CHIKV-E2 Monoclonal Antibody for Chikungunya Diagnosis. Virol. Sin. 2019, 34, 563–571. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Kumar, P.; Mohan, T.; Verma, P.; Parida, M.M.; Hoti, S.L.; Rao, D.N. Evaluation of Multiple Antigenic Peptides Based on the Chikungunya E2 Protein for Improved Serological Diagnosis of Infection. Viral Immunol. 2015, 28, 107–112. [Google Scholar] [CrossRef]
- Verma, P.; Bhatnagar, S.; Kumar, P.; Chattree, V.; Parida, M.M.; Hoti, S.L.; Ali, S.; Rao, D.N. Analysis of Antibody Response (IgM, IgG, IgG3) to Chikungunya Virus Using Panel of Peptides Derived from Envelope Protein for Serodiagnosis. Clin. Chem. Lab. Med. 2014, 52, 297–307. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Method | Target of the Assay | Sensitivity | Specificity | Reference |
|---|---|---|---|---|
| ELISA | Chikungunya viral antigen | 85% | / | [32] |
| Immuno-chromatography | Chikungunya viral antigen | 89.4% | 94.4% | [33] |
| Immuno-chromatography | E | 93.7% | 95.5% | [34] |
| Immuno-chromatography | E1 | 8/9 (88.9%, 95% CI 56.5–98.0) ECSA genotype, 10/30 (33.3%, 95% CI 19.2–51.2) | 49/59 (83.1%, 95% CI 71.5–90.5) | [35] |
| FLISA | E1 | / | / | [36] |
| Immuno-chromatography | E1 | 92% (92/100) (95% CI 85.0–95.9) | 100% (100/100) | [37] |
| Lateral flow assay/ELISA | E1/E2 | 51% (ELISA) and 100% (LFA) | 96.67% (ELISA) and 100% (LFA) | [38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simo, F.B.N.; Burt, F.J.; Makoah, N.A. Chikungunya Virus Diagnosis: A Review of Current Antigen Detection Methods. Trop. Med. Infect. Dis. 2023, 8, 365. https://doi.org/10.3390/tropicalmed8070365
Simo FBN, Burt FJ, Makoah NA. Chikungunya Virus Diagnosis: A Review of Current Antigen Detection Methods. Tropical Medicine and Infectious Disease. 2023; 8(7):365. https://doi.org/10.3390/tropicalmed8070365
Chicago/Turabian StyleSimo, Fredy Brice Nemg, Felicity Jane Burt, and Nigel Aminake Makoah. 2023. "Chikungunya Virus Diagnosis: A Review of Current Antigen Detection Methods" Tropical Medicine and Infectious Disease 8, no. 7: 365. https://doi.org/10.3390/tropicalmed8070365