
Molecular Characteristics, Expression Patterns, and Response of Insulin-like Growth Factors Gene Induced by Sex Steroid Hormones in Blotched Snakehead (Channa maculata)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Sampling
2.2. Hormone Treatment
2.3. RNA Extraction and cDNA Synthesis
2.4. Full-Length cDNA Cloning and Sequence Analysis for IGFs in Blotched Snakehead (C. maculata)
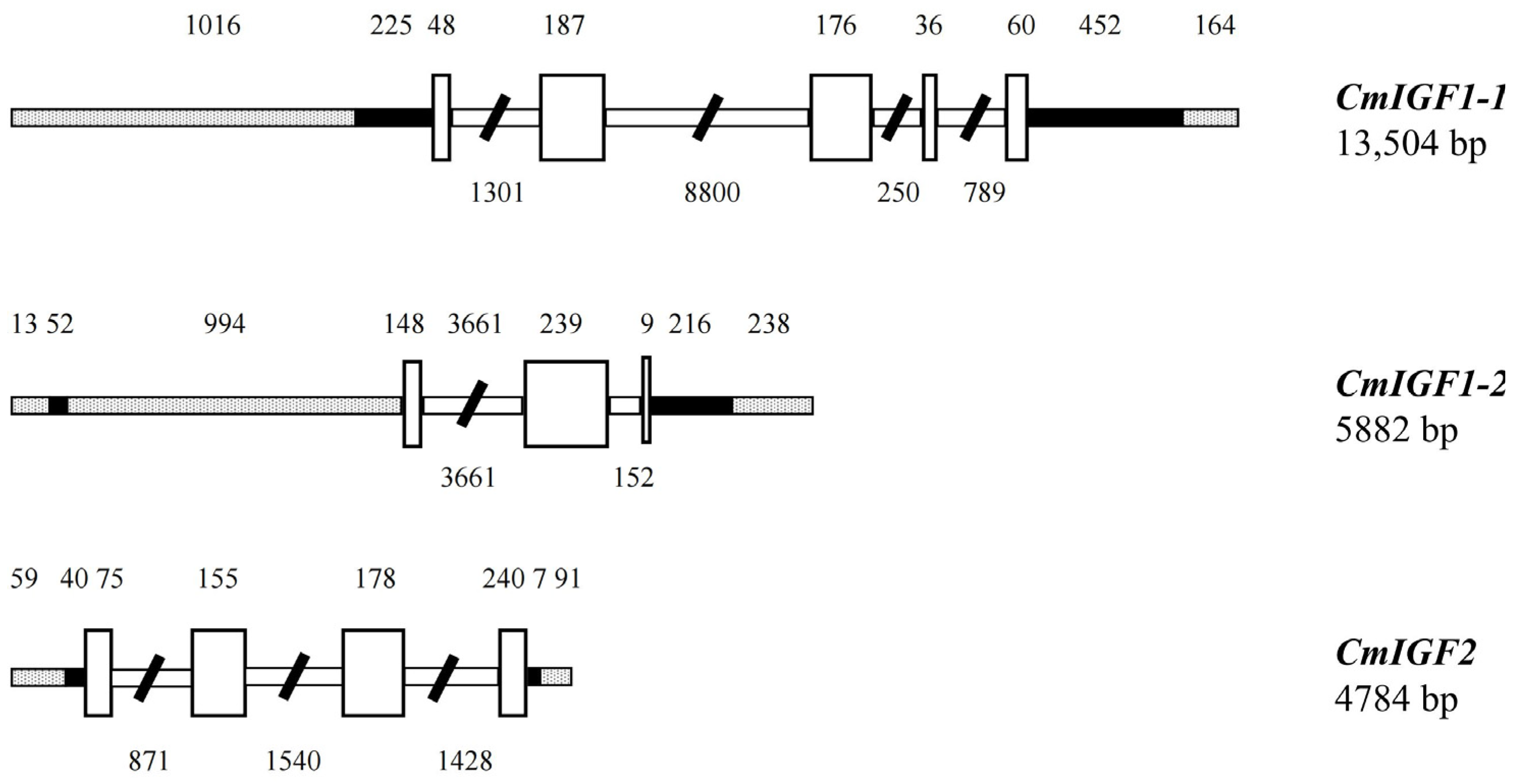
2.5. Cloning the Genomic Sequence of IGFs Genes in Blotched Snakehead (C. maculata)
2.6. Quantitative Real-Time PCR (qPCR)
2.7. Statistical Analysis
3. Results
3.1. Characterization of CmIGFs
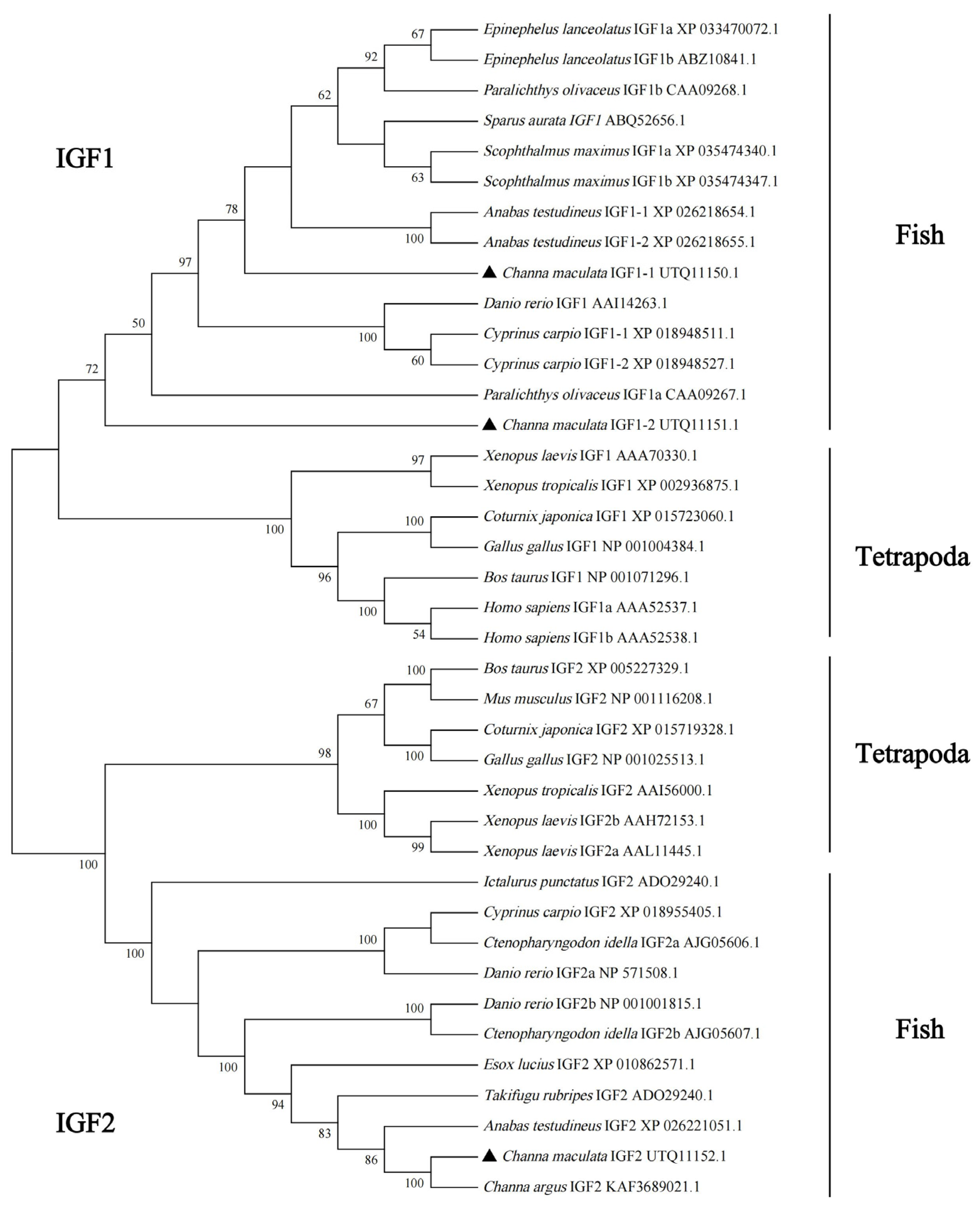
3.2. Multiple Alignments and Phylogenetic Analysis
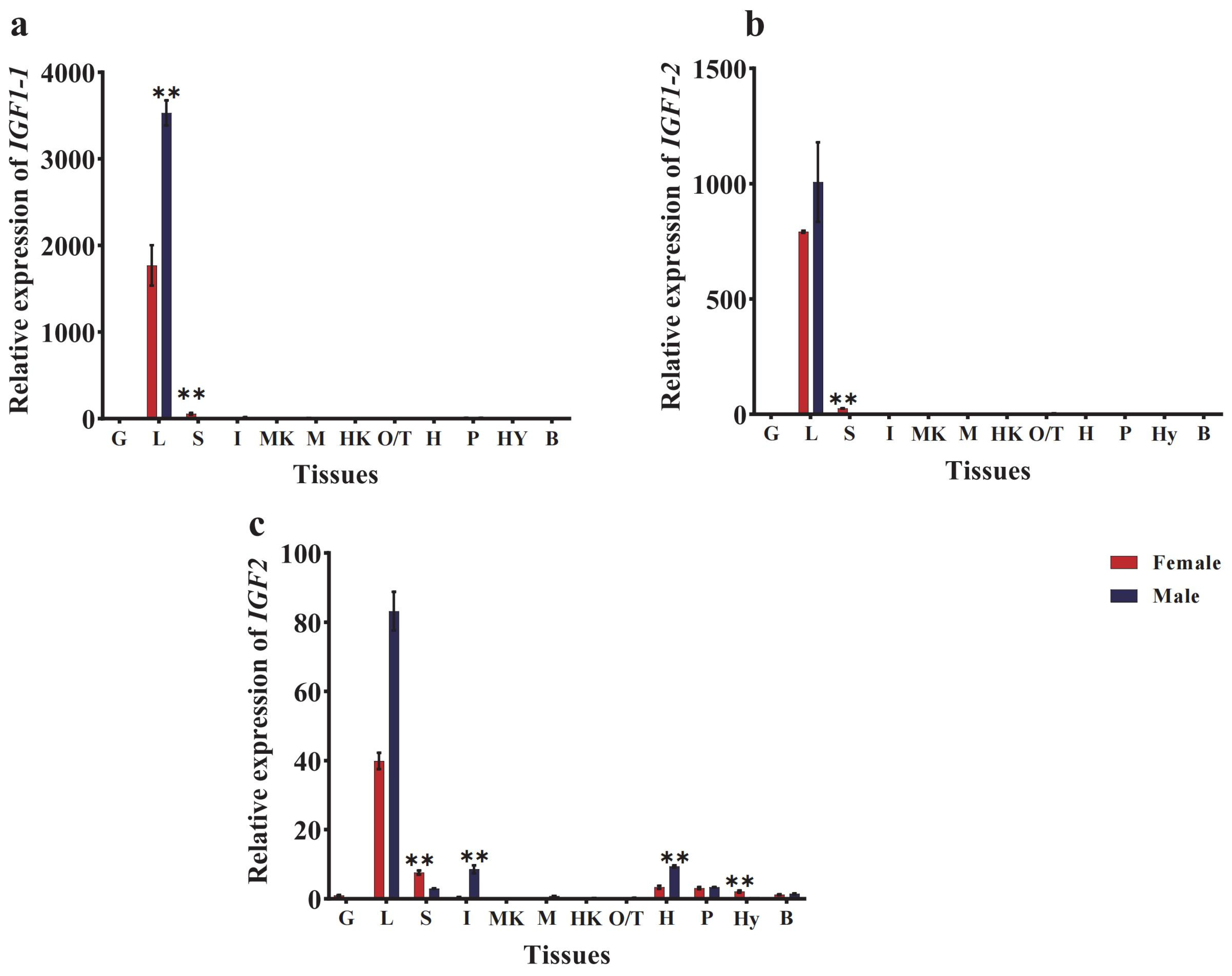
3.3. Tissue Distribution of CmIGFs
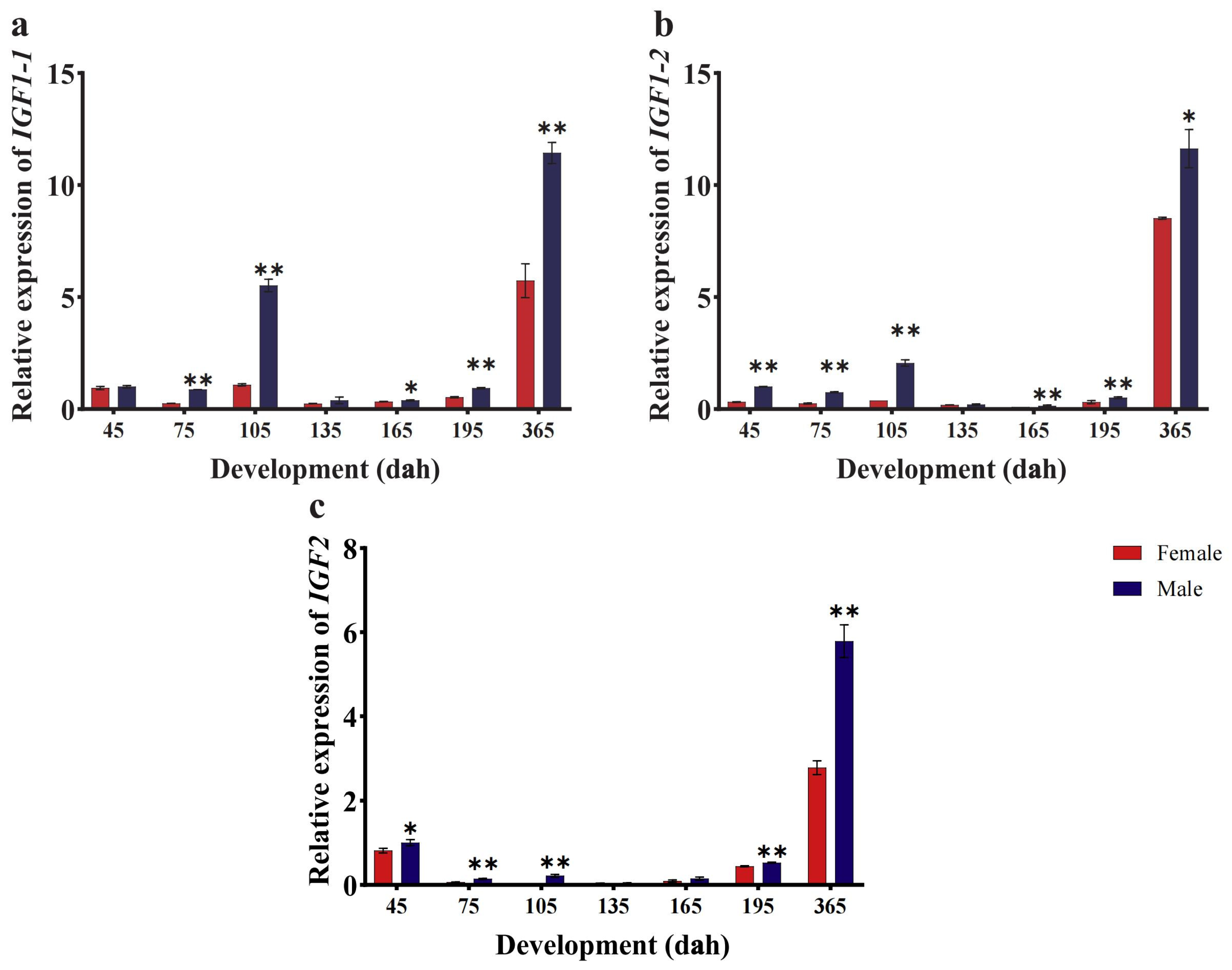
3.4. CmIGFs Expression Patterns at Different Developmental Stages
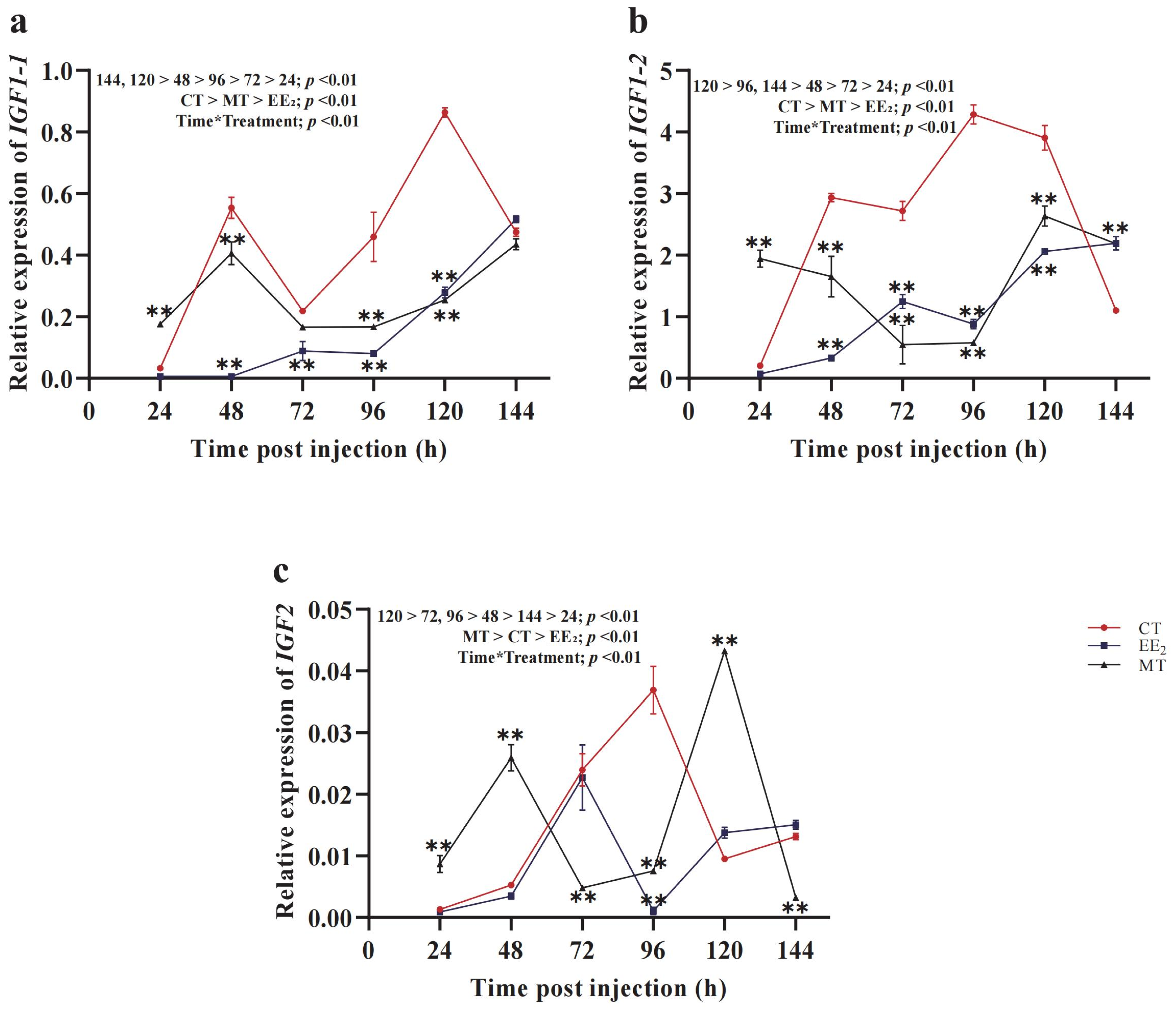
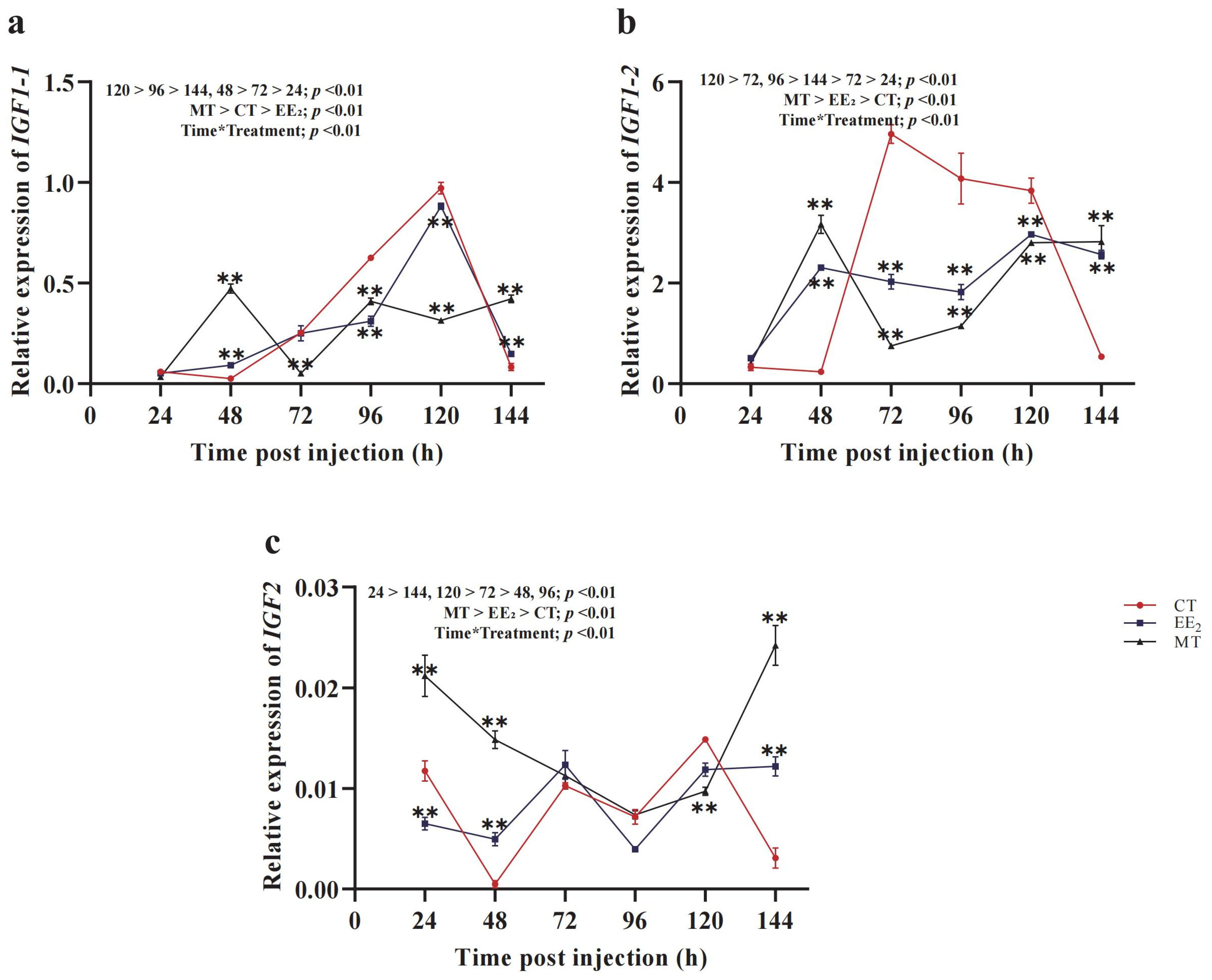
3.5. Effects of Short-Term Hormone Treatment on CmIGFs in Females
3.6. Effects of Short-Term Hormone Treatment on CmIGFs in Males
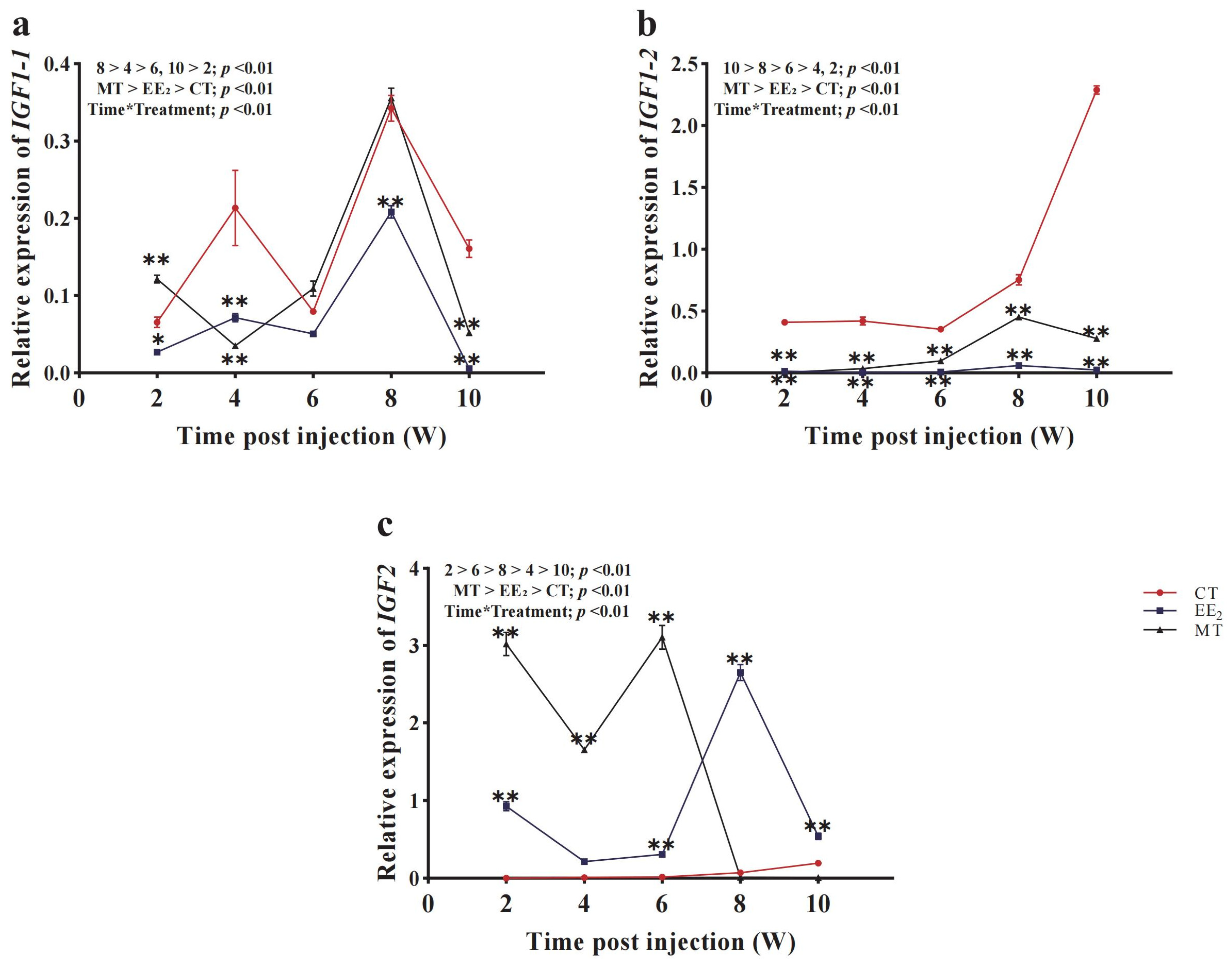
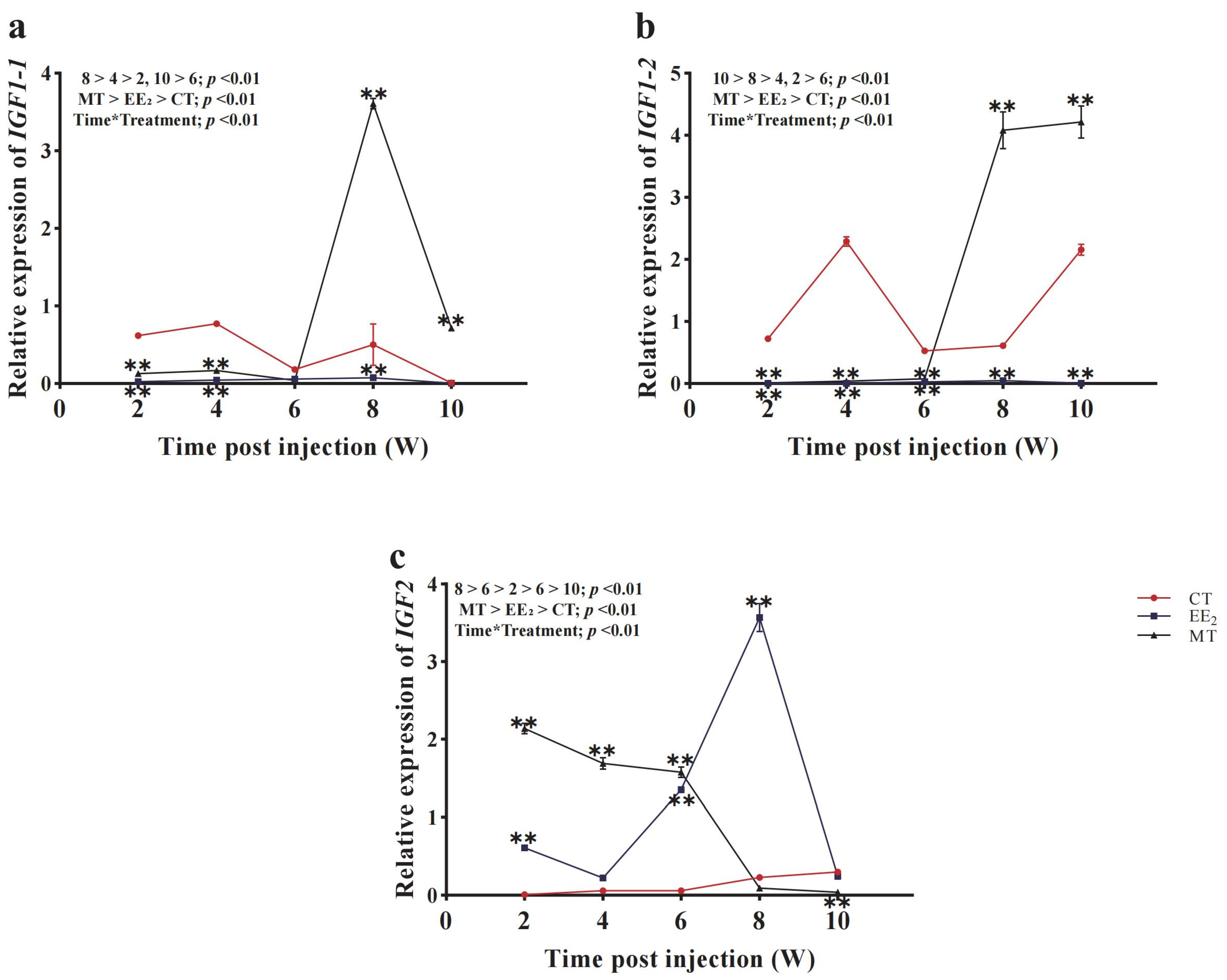
3.7. Effects of Long-Term Hormone Treatment on CmIGFs in Females
3.8. Effects of Long-Term Hormone Treatment on CmIGFs in Males
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dixit, M.; Poudel, S.B.; Yakar, S. Effects of GH/IGF axis on bone and cartilage. Mol. Cell. Endocrinol. 2021, 519, 111052. [Google Scholar] [CrossRef] [PubMed]
- Ranke, M.B.; Wit, J.M. Growth hormone—Past, present and future. Nat. Rev. Endocrinol. 2018, 14, 285–300. [Google Scholar] [CrossRef]
- Sun, C.; Sun, H.; Dong, J.; Tian, Y.; Hu, J.; Ye, X. Correlation analysis of mandarin fish (Siniperca chuatsi) growth hormone gene polymorphisms and growth traits. J. Genet. 2019, 98, 58. [Google Scholar] [CrossRef]
- Zhao, C.; Zheng, S.; Dang, Y.; Wang, M.; Ren, Y. Identification of a new insulin-like growth factor 3 (igf3) in turbot (Scophthalmus maximus): Comparison and expression analysis of IGF system genes during gonadal development. Fishes 2023, 8, 240. [Google Scholar] [CrossRef]
- Baral, K.; Rotwein, P. The Insulin-like The insulin-like growth factor 2 gene in mammals: Organizational complexity within a conserved locus. PLoS ONE 2019, 14, e0219155. [Google Scholar] [CrossRef]
- Cao, Q.; Duguay, S.J.; Plisetskaya, E.; Steiner, D.F.; Shu, J. Nucleotide sequence and growth hormone-regulated expression of salmon insulin-like growth factor I mRNA. Mol. Endocrinol. 1989, 3, 2005–2010. [Google Scholar] [CrossRef] [PubMed]
- Bersin, T.V.; Cordova, K.L.; Journey, M.L.; Beckman, B.R.; Lema, S.C. Food deprivation reduces sensitivity of liver Igf1 synthesis pathways to growth hormone in juvenile gopher rockfish (Sebastes carnatus). Gen. Comp. Endocrinol. 2024, 346, 114404. [Google Scholar] [CrossRef]
- Caelers, A.; Berishvili, G.; Meli, M.L.; Eppler, E.; Reinecke, M. Establishment of a real-time RT-PCR for the determination of absolute amounts of IGF-I and IGF-II gene expression in liver and extrahepatic sites of the tilapia. Gen. Comp. Endocrinol. 2004, 137, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Kamei, H.; Modi, Z.; Duan, C. Zebrafish IGF genes: Gene duplication, conservation and divergence, and novel roles in midline and notochord development. PLoS ONE 2009, 4, e7026. [Google Scholar] [CrossRef]
- Ma, Q.; Liu, S.; Zhuang, Z.; Sun, Z.; Liu, C.; Su, Y.; Tang, Q. Molecular cloning, expression analysis of insulin-like growth factor I (IGF-I) gene and IGF-I serum concentration in female and male tongue sole (Cynoglossus semilaevis). Comp. Biochem. Physiol. Part B Biocchem. Mol. Biol. 2011, 160, 208–214. [Google Scholar] [CrossRef]
- Ma, W.; Wu, J.; Zhang, J.; He, Y.; Gui, J.; Mei, J. Sex differences in the expression of GH/IGF axis genes underlie sexual size dimorphism in the yellow catfish (Pelteobagrus fulvidraco). Sci. China Life Sci. 2016, 59, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Chen, H.; Jiang, D.; Deng, S.; Zhu, C.; Wu, T.; Li, G. Insulin-like growth factors 1 and 2 in spotted scat (Scatophagus argus): Molecular cloning and differential expression during embryonic development. J. Guangdong Ocean. Uni. 2018, 38, 7–14. (In Chinese) [Google Scholar] [CrossRef]
- Shamblott, M.J.; Chen, T.T. Identification of a second insulin-like growth factor in a fish species. Proc. Natl. Acad. Sci. USA 1992, 89, 8913–8917. [Google Scholar] [CrossRef]
- Yuan, Y.; Hong, Y. Medaka insulin-like growth factor-2 supports self-renewal of the embryonic stem cell line and blastomeres in vitro. Sci. Rep. 2017, 7, 78. [Google Scholar] [CrossRef]
- White, Y.A.R.; Kyle, J.T.; Wood, A.W. Targeted gene knockdown in zebrafish reveals distinct intraembryonic functions for insulin-like growth factor II signaling. Endocrinology 2009, 150, 4366–4375. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zeng, L.; Duan, D.; Zhang, H.; Wang, Y.; Li, W.; Lin, H. Growth hormone and two forms of insulin-like growth factors I in the giant grouper (Epinephelus lanceolatus): Molecular cloning and characterization of tissue distribution. Fish Physiol. Biochem. 2010, 36, 201–212. [Google Scholar] [CrossRef]
- Yuan, X.; Jiang, X.; Pu, J.; Li, Z.; Zou, S. Functional conservation and divergence of duplicated insulin-like growth factor 2 genes in grass carp (Ctenopharyngodon idellus). Gene 2011, 470, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Zhao, J.; Tang, S.; Zhao, Y. Effects of estradiol and testosterone on the expression of growth-related genes in female and male Nile tilapia, Oreochromis niloticus. J. World Aquac. Soc. 2018, 49, 216–228. [Google Scholar] [CrossRef]
- Nipkow, M.; Wirthgen, E.; Luft, P.; Rebl, A.; Hoeflich, A.; Goldammer, T. Characterization of igf1 and igf2 genes during maraena whitefish (Coregonus maraena) ontogeny and the effect of temperature on embryogenesis and igf expression. Growth Horm. Igf. Res. 2018, 40, 32–43. [Google Scholar] [CrossRef]
- Venken, K.; Boonen, S.; Kopchick, J.; Coschigano, K.; Moverare, S.; Bouillon, R.; Ohlsson, C.; Vanderschueren, D. Growth without growth hormone receptor: Estradiol is a major growth-hormone independent regulator of hepatic insulin-like growth factor-I synthesis. J. Bone Miner. Res. 2004, 19, S43. [Google Scholar] [CrossRef]
- Norbeck, L.A.; Sheridan, M.A. An in vitro model for evaluating peripheral regulation of growth in fish: Effects of 17β-estradiol and testosterone on the expression of growth hormone receptors, insulin-like growth factors, and insulin-like growth factor type 1 receptors in rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol. 2011, 173, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, Y.; Chen, Y.; Zhou, L. Steroid hormones (E2 and MT) displayed difference in sex for Nile tilapia Oreochromis niloticus. Oceanol. Limnol. Sin. 2015, 46, 1487–1493. (In Chinese) [Google Scholar]
- Shen, M.; Zhu, J.; Wu, T.; Ren, T.; Zhang, Z.; Li, X.; Liu, F.; Zheng, S. Cloning and expression analysis of insunlin-like growth factor-I gene before and after reproduction in Zacco platypus. Acta Hydrobiol. Sin. 2020, 44, 1182–1190. [Google Scholar] [CrossRef]
- Ponce, M.; Infante, C.; Funes, V.; Manchado, M. Molecular characterization and gene expression analysis of insulin-like growth factors I and II in the redbanded seabream, Pagrus auriga: Transcriptional regulation by growth hormone. Comp. Biochem. Physiol. Part B Biocchem. Mol. Biol. 2008, 150, 418–426. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, T.; Chen, H.; Jiang, D.; Zhu, C.; Deng, S.; Zhang, Y.; Li, G. Estradiol-17 beta regulates the expression of insulin-like growth factors 1 and 2 via estradiol receptors in spotted scat (Scatophagus argus). Comp. Biochem. Physiol. Part B Biocchem. Mol. Biol. 2019, 237, 110328. [Google Scholar] [CrossRef]
- Ou, M.; Chen, K.; Gao, D.; Wu, Y.; Luo, Q.; Liu, H.; Zhao, J. Characterization, expression and cpg methylation analysis of dmrt1 and its response to steroid hormone in blotched snakehead (Channa maculata). Comp. Biochem. Physiol. Part B Biocchem. Mol. Biol. 2022, 257, 110672. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.; Chen, K.; Gao, D.; Wu, Y.; Chen, Z.; Luo, Q.; Liu, H.; Zhao, J. Comparative transcriptome analysis on four types of gonadal tissues of blotched snakehead (Channa maculata). Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 35, 100708. [Google Scholar] [CrossRef]
- Gao, D.; Ou, M.; Wu, Y.; Chen, K.; Liu, H.; Luo, Q.; Zhao, J. Gene cloning and expression analysis of growth hormonr gene from blotched snakehead (Channa maculata). J. Agric. Biotech. 2021, 29, 2328–2341. Available online: http://journal05.magtech.org.cn/Jwk_ny/EN/10.3969/j.issn.1674-7968.2021.12.007 (accessed on 18 January 2024). (In Chinese).
- Zhao, J.; Ou, M.; Wang, Y.; Liu, H.; Luo, Q.; Zhu, X.; Chen, B.; Chen, K. Breeding of YY super-male of blotched snakehead (Channa maculata) and production of all-male hybrid (Channa argus ♀ × C. maculata ♂). Aquaculture 2021, 538, 736450. [Google Scholar] [CrossRef]
- Ou, M.; Huang, R.; Yang, C.; Gui, B.; Luo, Q.; Zhao, J.; Li, Y.; Liao, L.; Zhu, Z.; Wang, Y.; et al. Chromosome-level genome assemblies of Channa argus and Channa maculata and comparative analysis of their temperature adaptability. Gigascience 2021, 10, giab070. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Chen, K.; Zhu, X.; Luo, Q.; Zhao, J.; Li, W.; Wu, X.; Xu, H. Identification of suitable reference genes for quantitative real-time PCR normalization in blotched snakehead Channa maculata. J. Fish Biol. 2017, 90, 2312–2322. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Wang, M.; Ning, J.; Fan, F.; Zhu, C. Cloning and association analysis with growth traits of igf gene in ussuri catfish Pseudobagrus ussuriensis. Fish Sci. 2022, 41, 738–748. (In Chinese) [Google Scholar] [CrossRef]
- Shamblott, M.J.; Chen, T. Age-related and tissue-specific levels of five forms of insulin-like growth factor mRNA in a teleost. Mol. Mar. Biol. Biotechnol. 1993, 2, 351–361. [Google Scholar]
- Ndandala, C.B.; Dai, M.S.; Mustapha, U.F.; Li, X.; Liu, J.; Huang, H.; Li, G.; Chen, H. Current research and future perspectives of GH and IGFs family genes in somatic growth and reproduction of teleost fish. Aquac. Rep. 2022, 26, 101289. [Google Scholar] [CrossRef]
- Sun, P.; Wu, Z.; You, F.; Li, J. Annual cycle change of sex steroid hormones in cultured Paralichthys olivaceus. Mar. Fish 2013, 35, 34–37. [Google Scholar] [CrossRef]
- Parey, E.; Louis, A.; Montfort, J. An atlas of fish genome evolution reveals delayed rediploidization following the teleost whole-genome duplication. Genome Res. 2022, 32, 1685–1697. [Google Scholar] [CrossRef]
- Qian, K.; Wen, H.; Chi, M.; Ni, M.; Zhang, D.; Ding, Y. Solation of full-length cDNA of Insulin-like growth factor-1 (IGF-1) gene of Japanese sea bass (Lateolabrax japonicas) and its expression analysis. J. Ocean Univ. China 2014, 44, 27–34. (In Chinese) [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Lai, J.; Song, M.; Gong, Q. Molecular cloning of insulin-like growth factor 1, 2 in Acipenser dabryanus and their expression level during starvation stress. Southwest China J. Agric. Sci. 2020, 33, 447–455. (In Chinese) [Google Scholar] [CrossRef]
- Degani, G.; Tzchori, I.; Yom-Din, S.; Goldberg, D.; Jackson, K. Growth differences and growth hormone expression in male and female European eels [Anguilla anguilla (L.)]. Gen. Comp. Endocrinol. 2003, 134, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Liu, S.; Zhuang, Z.; Lin, L.; Sun, Z.; Liu, C.; Ma, H.; Su, Y.; Tang, Q. Genomic structure, polymorphism and expression analysis of the growth hormone (GH) gene in female and male half-smooth tongue sole (Cynoglossus semilaevis). Gene 2012, 493, 92–104. [Google Scholar] [CrossRef]
- Yang, H.; Chen, H.; Zhao, H.; Liu, L.; Xie, Z.; Xiao, L.; Li, S.; Zhang, Y.; Lin, H. Molecular cloning of the insulin-like growth factor 3 and difference in the expression of igf genes in orange-spotted grouper (Epinephelus coioides). Comp. Biochem. Phys. B 2015, 186, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Wang, L.; Zhu, W.; Fu, J.; Dong, J.; Dong, Z. A novel igf3 gene in common carp (Cyprinus carpio): Evidence for its role in regulating gonadal development. PLoS ONE 2016, 7, e1002447. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ou, M.; Gao, D.; Chen, K.; Luo, Q.; Liu, H.; Zhao, J. Molecular cloning, expression and respone of foxl2 gene induced by sex steroid hormones in blotched snakehead Channa maculata. J. Dalian Ocean Uni. 2022, 37, 49–60. (In Chinese) [Google Scholar] [CrossRef]
- Li, X.; Mei, J.; Ge, C.; Liu, X.; Gui, J. Sex determination mechanisms and sex control approaches in aquaculture animals. Sci. China Life Sci. 2022, 65, 1091–1122. [Google Scholar] [CrossRef]
- Hu, Y.; Zou, Z.; Zhu, J.; Li, D.; Xiao, W.; Han, Y.; Le, Y.; Wang, T.; Yang, H. The expression analysis of IGF1 gene during different development stages in Nile tilapia. Chin. Agric. Sci. Bull. 2014, 30, 107–111. (In Chinese) [Google Scholar]
- Liu, J.; Zhao, J.; Zhang, M.; Dai, W. Cloning and expression of full-length cDNA of insulin-like growth factor-Ⅱ in mandarin fish Siniperca chuatsi. J. Dalian Ocean Uni. 2012, 27, 495–501. (In Chinese) [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, H.; Wang, T.; Wang, D.; Zhang, H.; Yin, S.; Chen, S.; Zhou, G. Cloning and temporal expression analysis of GHR and IGF-2 in Odontobutis potamophila. Mar. Fish 2019, 41, 421–433. (In Chinese) [Google Scholar] [CrossRef]
- James, M.O. Steroid catabolism in marine and freshwater fish. J. Steroid. Biochem. Mol. Biol. 2011, 127, 167–175. [Google Scholar] [CrossRef]
- Shved, N.; Berishvili, G.; D’Cotta, H.; Baroiller, J.F.; Segner, H.; Eppler, E.; Reinecke, M. Ethinylestradiol differentially interferes with IGF-I in liver and extrahepatic sites during development of male and female bony fish. J. Endocrinol. 2007, 195, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Wu, T.; Zhang, Y.; Gao, J.; Yang, Y.; Qin, F.; Liu, S.; Zheng, Y.; Wang, Z. Responsiveness of four gender-specific genes, figla, foxl2, scp3 and sox9a to 17α-ethinylestradiol in adult rare minnow Gobiocypris rarus. Gen. Comp. Endocrinol. 2014, 200, 44–53. [Google Scholar] [CrossRef]
- Liu, S.; Lv, W.; Lv, X. Effect of 17α-methyltestosterone (MT) on mRNA expression of vtg gene in liver of Brachydanio rerio. Anim. Husb. Feed Sci. 2016, 3, 9–11. (In Chinese) [Google Scholar] [CrossRef]
- Fruchtman, S.; McVey, D.C.; Borski, R.J. Characterization of pituitary IGF-I receptors: Modulation of prolactin and growth hormone. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R468–R476. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′~3′) | Application |
|---|---|---|
| IGF1-1-F1 | CTCCTGTAGCCACACCCTC | Partial sequence obtaining |
| IGF1-1-R1 | GAATGACTGTGTCCAGGTAAAG | |
| IGF1-2-F1 | ATGGGCTGTATCTCCTGTAGTC | |
| IGF1-2-R1 | AGTATTCTCGGCAAGTCGGT | |
| IGF2-F1 | AGCAAAGATACGGACAGCAC | |
| IGF2-R1 | GTTGACATAGTTATCCGTGGC | |
| IGF1-1-5′R-out | AAGCCTCTTTCTCCACACACAAACTGC | 5′-RACE PCR amplification |
| IGF1-1-5′R-in | GCAGTGAGAGGGTGTGGCTACAGGAG | |
| IGF1-2-5′R-out | AGCGTGTGGGTTTACT | |
| IGF1-2-5′R-in | TGTCGACCAGCTCCACCC | 3′-RACE PCR amplification |
| IGF1-1-3′F-out | CCTGCCAAGACTAACAAGCCAACTCG | |
| IGF1-1-3′F-in | GAGAACAACTAAGAGACCTTTACCTGGACACA | |
| IGF1-1-F2 | CCTGTTCGCTAAATCTCACTTCTC | ORF identification |
| IGF1-1-R2 | CATTTGTCCATTCGCTCCATC | |
| IGF1-2-F2 | GGACTACAAGAGAGACGG | |
| IGF1-2-R2 | TTTGTCCCTTCGCTCCAT | |
| IGF2-F2 | AGCCAAATAACCCCCAACA | |
| IGF2-R2 | AGCGGGCTCATTTGTGG | |
| IGF1-1-DL-F | CGCTCTTTCCTCTCAGTGGC | qPCR amplification |
| IGF1-1-DL-R | CCATAGCCTGTGGGTTTACTGA | |
| IGF1-2-DL-F | GTTTGTGTGTGGAGACAGAGGC | |
| IGF1-2-DL-R | GCACGCACAGAGTGAGTTGG | |
| IGF2-DL-F | GTCTTCGTCCAGTCGTTCGC | |
| IGF2-DL-R | TGTTGCCCCTGCTGGTTG | |
| β-actin-F | GCAAGCAGGAGTATGATGAG | |
| β-actin-R | TTGGGATTGTTTCAGTCAGT | |
| EF1α-F | GGGAGACCCACAATAACATCG | |
| EF1α-R | CCAGGCATACTTGAAGGAGC |
| Primer Name | Sequence (5′~3′) | Length (bp) |
|---|---|---|
| IGF1-1-gDNA-F1 | TTTATGATTGGGTCACAGCA | 1674 |
| IGF1-1-gDNA-R1 | GGACTCAGCAGGAATTACTCT | |
| IGF1-1-gDNA-F2 | GTTACTTACTGGCAGGTTTT | 1827 |
| IGF1-1-gDNA-R2 | TGTTTGGGTTCTACTCAATT | |
| IGF1-1-gDNA-F3 | GGCGGCAAATTAGAGTTGTG | 1848 |
| IGF1-1-gDNA-R3 | ATGGACGAACTGAGGTTACAAG | |
| IGF1-1-gDNA-F4 | ACAAACGCTGTGAAGTGGTC | 1764 |
| IGF1-1-gDNA-R4 | CAGGGAGCTACTTAATGCTTA | |
| IGF1-1-gDNA-F5 | GGGTGATTTCACTGGGATGT | 1962 |
| IGF1-1-gDNA-R5 | AACCTGTGGATTCTTGGAGC | |
| IGF1-1-gDNA-F6 | TAGTCCCTGCCCAGCCGTAA | 1997 |
| IGF1-1-gDNA-R6 | AGAAACAAAGCATAGGTGAA | |
| IGF1-1-gDNA-F7 | TCGATTCCCCTGTCCCCTAA | 2008 |
| IGF1-1-gDNA-R7 | GCCTGCGTTTCGACTTCACG | |
| IGF1-1-gDNA-F8 | GATGTTTTAGGCAGCGTCTG | 1588 |
| IGF1-1-gDNA-R8 | AACCGTGTTTTACTCTTTTAG | |
| IGF1-2-gDNA-F1 | TGCCTTTGTAGTTTACCTTT | 1829 |
| IGF1-2-gDNA-R1 | TAATTTGTCCCCTTTATTCG | |
| IGF1-2-gDNA-F2 | ATGCTAGGACTGAAATGCTA | 1727 |
| IGF1-2-gDNA-R2 | TAGATGATAAATAACGGGTA | |
| IGF1-2-gDNA-F3 | GTCAGTGCTGTTCTTTCCAA | 1155 |
| IGF1-2-gDNA-R3 | AAAAGGGGCTGTGCCTTGTT | |
| IGF1-2-gDNA-F4 | AAGTGAAGCATTTCAAACTT | 1546 |
| IGF1-2-gDNA-R4 | TAACTGGCAGAAGATGACTA | |
| IGF2-gDNA-F1 | GGAGGAGCGATGGGTGGTGG | 1677 |
| IGF2-gDNA-R1 | AGCGGCCCATTGTCAGTCCG | |
| IGF2-gDNA-F2 | GTCTCAAGACTTCGTCCAGG | 1623 |
| IGF2-gDNA-R2 | GTCTCAAGACTTCGTCCAGG | |
| IGF2-gDNA-F3 | CCACTATGGGAAACAATGCC | 1742 |
| IGF2-gDNA-R3 | AGGACTGCCACAGAAATCAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wu, Y.; Zhang, Y.; Zhang, J.; Chen, K.; Liu, H.; Luo, Q.; Fei, S.; Zhao, J.; Ou, M. Molecular Characteristics, Expression Patterns, and Response of Insulin-like Growth Factors Gene Induced by Sex Steroid Hormones in Blotched Snakehead (Channa maculata). Fishes 2024, 9, 120. https://doi.org/10.3390/fishes9040120
Zhang X, Wu Y, Zhang Y, Zhang J, Chen K, Liu H, Luo Q, Fei S, Zhao J, Ou M. Molecular Characteristics, Expression Patterns, and Response of Insulin-like Growth Factors Gene Induced by Sex Steroid Hormones in Blotched Snakehead (Channa maculata). Fishes. 2024; 9(4):120. https://doi.org/10.3390/fishes9040120
Chicago/Turabian StyleZhang, Xiaotian, Yuxia Wu, Yang Zhang, Jin Zhang, Kunci Chen, Haiyang Liu, Qing Luo, Shuzhan Fei, Jian Zhao, and Mi Ou. 2024. "Molecular Characteristics, Expression Patterns, and Response of Insulin-like Growth Factors Gene Induced by Sex Steroid Hormones in Blotched Snakehead (Channa maculata)" Fishes 9, no. 4: 120. https://doi.org/10.3390/fishes9040120