
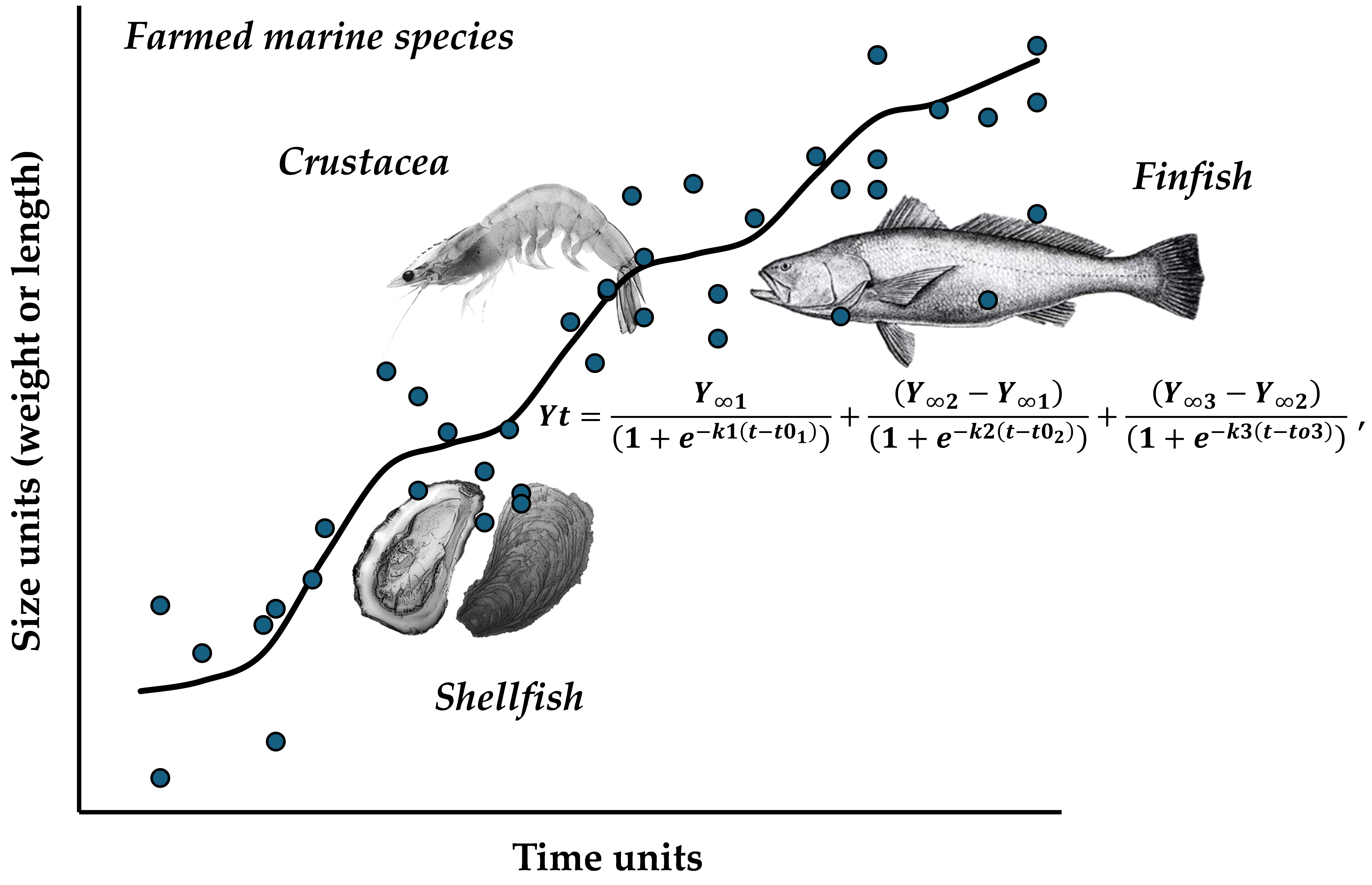
The Fractal Approach to Describe Growth of Farmed Marine Species: Using Double and Triple Logistic Models

 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
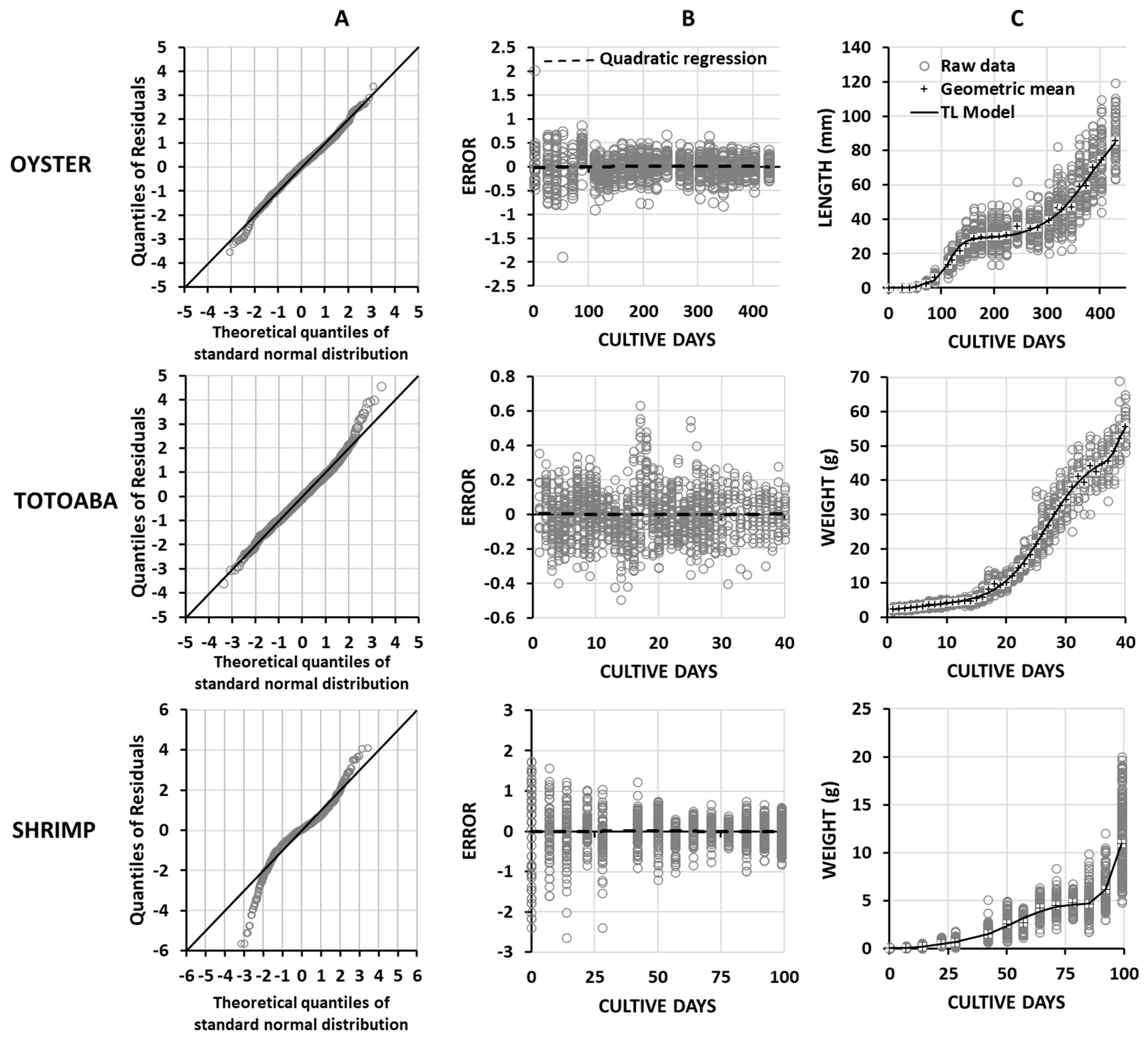
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernandez-Llamas, A.; Ratkowsky, D.A. Growth of fishes, crustaceans and molluscs: Estimation of the von Bertalanffy, Logistic, Gompertz and Richards curves and a new growth model. Mar. Ecol. Progr. Ser. 2004, 282, 237–244. [Google Scholar] [CrossRef]
- Ansah, Y.B.; Frimpong, E.A. Using model-based inference to select a predictive growth curve for farmed Tilapia. N. Am. J. Aquac. 2015, 77, 281–288. [Google Scholar] [CrossRef]
- Baer, A.; Schulz, C.; Traulsen, I.; Krieter, J. Analysing the growth of turbot (Psetta maxima) in a commercial recirculation system with the use of three different growth models. Aquac. Int. 2011, 19, 497–511. [Google Scholar] [CrossRef]
- Pauly, D. Length-converted catch curves and the seasonal growth curves of fishes. Fishbyte 1990, 8, 24–29. [Google Scholar]
- Rodríguez-Domínguez, G.; Castillo-Vargasmachuca, S.G.; Ramírez-Pérez, J.S.; Pérez-González, R.; Aragón-Noriega, E.A. Modelos múltiples para determinar el crecimiento de organismos juveniles de jaiba azul Callinectes arcuatus en cautiverio. Cienc. Pesq. 2014, 22, 29–35. [Google Scholar]
- Day, T.; Taylor, P.D. Von Bertalanffy’s growth equation should not be used to model age and size at maturity. Am. Nat. 1997, 149, 381–393. [Google Scholar] [CrossRef]
- Semmens, J.M.; Pecl, G.T.; Villanueva, R.; Jouffre, D.; Sobrino, I.; Wood, J.B.; Rigby, P.R. Understanding octopus growth: Patterns, variability and physiology. Mar. Freshw. Res. 2004, 55, 367–377. [Google Scholar] [CrossRef]
- Modis, T. Fractal Aspects of Natural Growth. Technol. Forecast. Soc. Change 1994, 47, 63–73. [Google Scholar] [CrossRef]
- Shabani, A.; Sepaskhah, A.R.; Kamgar-Haghighi, A.A.; Honar, T. Using double logistic equation to describe the growth of winter rapeseed. J. Agric. Sci. 2018, 156, 37–45. [Google Scholar] [CrossRef]
- Flinn, S.A.; Midway, S.R. Trends in Growth Modeling in Fisheries Science. Fishes 2021, 6, 1. [Google Scholar] [CrossRef]
- Zhu, L.; Li, L.; Liang, Z. Comparison of six statistical approaches in the selection of appropriate fish growth models. Chin. J. Oceanogr. Limnol. 2009, 27, 457–467. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; pp. 1–488. [Google Scholar]
- Diken, G.; Demir, O.; Naz, M. The inhibitory situational analysis of some feed ingredients for meagre, Argyrosomus regius (Asso 1801) larvae and evaluation for diet formulations. Aquat. Res. 2019, 2, 41–52. [Google Scholar] [CrossRef]
- Curiel-Bernal, M.V.; Cisneros-Mata, M.A.; Rodríguez-Domínguez, G.; Sánchez-Velasco, L.; Jiménez-Rosenberg, S.P.A.; Parés-Sierra, A.; Aragón-Noriega, E.A. Modelling early growth of Totoaba macdonaldi under laboratory conditions. Fishes 2023, 8, 155. [Google Scholar] [CrossRef]
- Valenzuela-Madrigal, I.E.; Valenzuela-Quiñonez, W.; Esparza-Leal, H.M.; Rodríguez-Quiroz, G.; Aragón-Noriega, E.A. Effects of ionic composition on growth and survival of white shrimp Litopenaeus vannamei culture at low-salinity well water. Rev. Biol. Mar. Oceanogr. 2017, 52, 103–112. [Google Scholar] [CrossRef]
- Góngora-Gómez, A.M.; Leal-Sepúlveda, A.L.; García-Ulloa, M.; Aragón-Noriega, E.A.; Valenzuela-Quiñónez, W. Morphometric relationships and growth models for the oyster Crassostrea corteziensis cultivated at the southeastern coast of the Gulf of California, Mexico. Lat. Am. J. Aquat. Res. 2018, 46, 735–743. [Google Scholar] [CrossRef]
- Mundry, R. Issues in information theory-based statistical inference a commentary from a frequentist’s perspective. Behav. Ecol. Sociobiol. 2011, 62, 57–68. [Google Scholar] [CrossRef]
- Hilborn, R.; Walters, C.J. Quantitative Fish Stock Assessment. Choice, Dynamics and Uncertainty, 1st ed.; Chapman and Hall: New York, NY, USA, 1992; pp. 410–428. [Google Scholar]
- Von Bertalanffy, L. A quantitative theory of organic growth. Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Hutchinson, C.E.; TenBrink, T.T. Age determination of the yellow Irish lord: Management implications as a result of new estimates of maximum age. N. Am. J. Fish. Manag. 2011, 31, 1116–1122. [Google Scholar] [CrossRef]
- Lee, L.; Atkinson, D.; Hirst, A.G.; Cornell, S.J. A new framework for growth curve fitting based on the von Bertalanffy growth function. Sci. Rep. 2020, 10, 7953. [Google Scholar] [CrossRef]
- Lorenzen, K. A simple von Bertalanffy model for density-dependent growth in extensive aquaculture, with an application to common carp (Cyprinus carpio). Aquaculture 1996, 142, 191–205. [Google Scholar] [CrossRef]
- De Graaf, G.; Prein, M. Fitting growth with the von Bertalanffy growth function: A comparison of three approaches of multivariate analysis of fish growth in aquaculture experiments. Aquac. Res. 2005, 36, 100–109. [Google Scholar] [CrossRef]
- Brunel, T.; Dickey-Collas, M. Effects of temperature and population density on von Bertalanffy growth parameters in Atlantic herring: A macro-ecological analysis. Mar. Ecol. Prog. Ser. 2010, 405, 15–28. [Google Scholar] [CrossRef]
- Martínez-Jerónimo, F. Description of the individual growth of Daphnia magna (Crustacea: Cladocera) through the von Bertalanffy growth equation. Effect of photoperiod and temperature. J. Limnol. 2012, 13, 65–71. [Google Scholar] [CrossRef]
- Flanagan, C.A. Study of the feasibility of modeling the totoaba fishery of the northern Gulf of California with preliminary estimation of some critical parameters. Univ. Ariz. Biol. Sci. Rep. 1973, 249, 1–58. [Google Scholar]
- Cisneros-Mata, M.Á.; Montemayor-López, G.; Román-Rodríguez, M.J. Life history and conservation of Totoaba macdonaldi. Conserv. Biol. 1995, 9, 806–814. [Google Scholar] [CrossRef]
- Pedrín-Osuna, O.; Córdova-Murueta, J.H.; Delgado-Marchena, M. Crecimiento y mortalidad de la totoaba, Totoaba macdonaldi, del Alto Golfo de California. Cienc. Pesq. 2001, 15, 131–140. [Google Scholar]
- De Anda-Montañez, J.A.; García de León, F.J.; Zenteno-Savín, T.; Balart-Páez, E.; Méndez-Rodríguez, L.C.; Bocanegra-Castillo, N.; Martínez-Aguilar, S.; Campos-Dávila, L.; Román Rodríguez, M.J.; Valenzuela-Quiñonez, F.; et al. Estado de Salud y Estatus de Conservación de la(s) Población(es) de Totoaba (Totoaba macdonaldi) en el Golfo de California: Una Especie en Peligro de Extinción; Informe Final, SNIB-CONABIO. Proyecto Núm. hK050; Centro de Investigaciones Biológicas del Noroeste, S.C.: La Paz, Mexico, 2013; Available online: http://www.conabio.gob.mx/institucion/proyectos/resultados/InfHK050.pdf (accessed on 3 February 2021).
- Valenzuela-Quiñonez, F. Genética y Dinámica Poblacional de la Totoaba (Totoaba macdonaldi Gilbert, 1891) en el Golfo de California. Ph.D. Thesis, Centro de Investigaciones Biológicas del Noroeste, S.C., La Paz, Mexico, 22 May 2014. [Google Scholar]
- Román-Rodríguez, M.; Hammann, M.G. Age and growth of totoaba, Totoaba macdonaldi (Sciaenidae) in the upper Gulf of California. Fish. Bull. 1997, 95, 620–628. [Google Scholar]
- Beverton, R.J.H.; Holt, S.J. On the Dynamics of Exploited Fish Populations, 1st ed.; Chapman and Hall: London, UK, 1957. [Google Scholar]


{kind=link}
{kind=link}
| AIC | BIC | ||||||
|---|---|---|---|---|---|---|---|
| Species | Model | N | Parameters | Normal | Log-Normal | Normal | Log-Normal |
| Crassostrea corteziensis Oyster | Triple Logistic | 1349 | 10 | 9358 | 6983 | 9410 | 7035 |
| Double Logistic | 7 | 9353 | 7608 | 9389 | 7644 | ||
| VB Oscillatory | 6 | 9662 | NC | 9693 | NC | ||
| Totoaba macdonaldi Finfish | Triple Logistic | 1391 | 10 | 6600 | 4658 | 6653 | 4711 |
| Double Logistic | 7 | 6732 | 4676 | 6769 | 4713 | ||
| VB Oscillatory | 6 | 6733 | 5287 | 6764 | 5319 | ||
| Penaeus vannamei Shrimp | Triple Logistic | 1809 | 10 | 6942 | 4677 | 6969 | 4732 |
| Double Logistic | 7 | 6971 | 4820 | 7010 | 4858 | ||
| VB Oscillatory | 6 | 7813 | NC | 7841 | NC | ||
| Species | ||||
|---|---|---|---|---|
| Logistic Harmonic | Parameters | Crassostrea corteziensis | Totoaba macdonaldi | Penaeus vannamei |
| 1 | L∞1 | 2.17 (1.96, 2.35) | 3.99 (3.58, 4.33) | 0.415 (0.269, 0.993) |
| k1 | 1.39 (1.39, 1.39) | 0.21 (0.15, 0.31) | 0.218 (0.155, 0.320) | |
| t01 | 52.9 (52.68, 52.93) | 0.004 (0.002, 0.090) | 19.27 (16.15, 25.31) | |
| 2 | L∞2 | 28.47 (27.6, 29.3) | 48.27 (46.11, 50.21) | 4.85 (4.58, 5.05) |
| k2 | 0.085 (0.082, 0.087) | 0.26 (0.25, 0.27) | 0.108 (0.103, 0.148) | |
| t02 | 114.9 (113.2, 116.6) | 26.69 (26.25, 27.08) | 52.34 (50.51, 54.8) | |
| 3 | L∞3 | 104.53 (94.61, 119.60) | 58.1 (53.51, 89.73) | 10.98 (10.43, 11.55) |
| k3 | 0.0225 (0.0219, 0.023) | 1.603 (0.564, 21.77) | 1.889 (1.888,1.911) | |
| t03 | 383.15 (370.0, 399.0) | 38.8 (37.33, 41.81) | 92.65 (92.47, 92.83) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Domínguez, G.; Aragón-Noriega, E.A.; Payán-Alejo, J.; Mendivil-Mendoza, J.E.; Curiel-Bernal, M.V.; Valenzuela-Quiñonez, W.; Urías-Sotomayor, R. The Fractal Approach to Describe Growth of Farmed Marine Species: Using Double and Triple Logistic Models. Fishes 2024, 9, 106. https://doi.org/10.3390/fishes9030106
Rodríguez-Domínguez G, Aragón-Noriega EA, Payán-Alejo J, Mendivil-Mendoza JE, Curiel-Bernal MV, Valenzuela-Quiñonez W, Urías-Sotomayor R. The Fractal Approach to Describe Growth of Farmed Marine Species: Using Double and Triple Logistic Models. Fishes. 2024; 9(3):106. https://doi.org/10.3390/fishes9030106
Chicago/Turabian StyleRodríguez-Domínguez, Guillermo, Eugenio Alberto Aragón-Noriega, Jorge Payán-Alejo, Jaime Edzael Mendivil-Mendoza, Marcelo Vidal Curiel-Bernal, Wenceslao Valenzuela-Quiñonez, and Ricardo Urías-Sotomayor. 2024. "The Fractal Approach to Describe Growth of Farmed Marine Species: Using Double and Triple Logistic Models" Fishes 9, no. 3: 106. https://doi.org/10.3390/fishes9030106