
Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf

Abstract
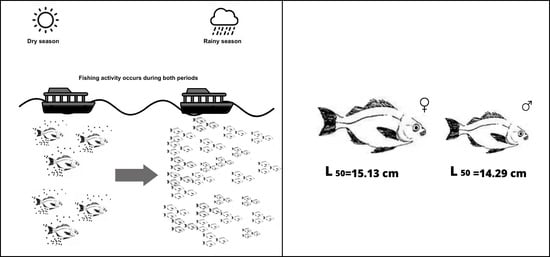
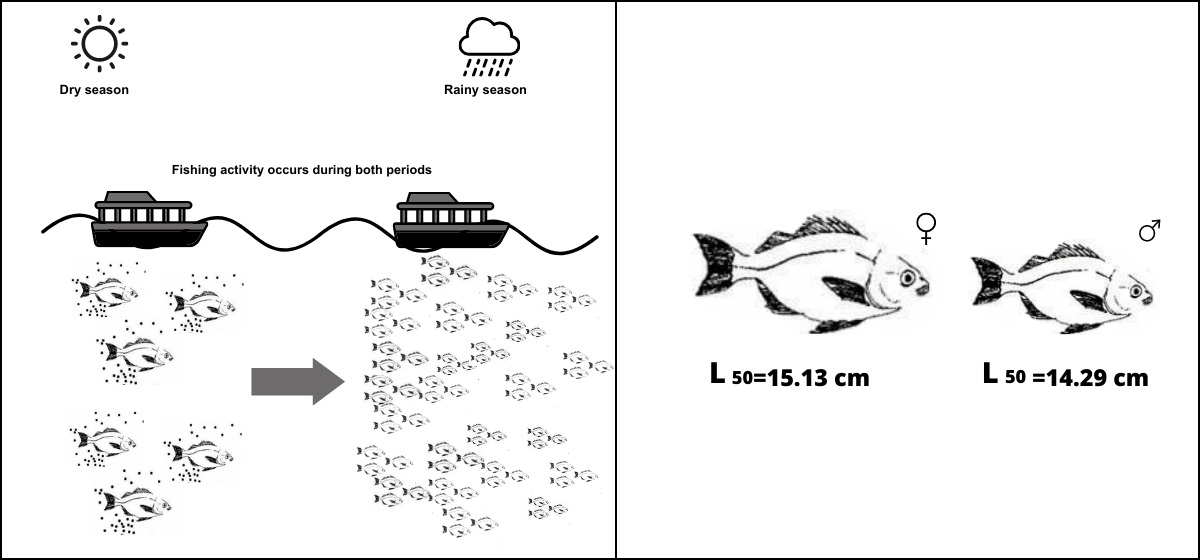
:
1. Introduction
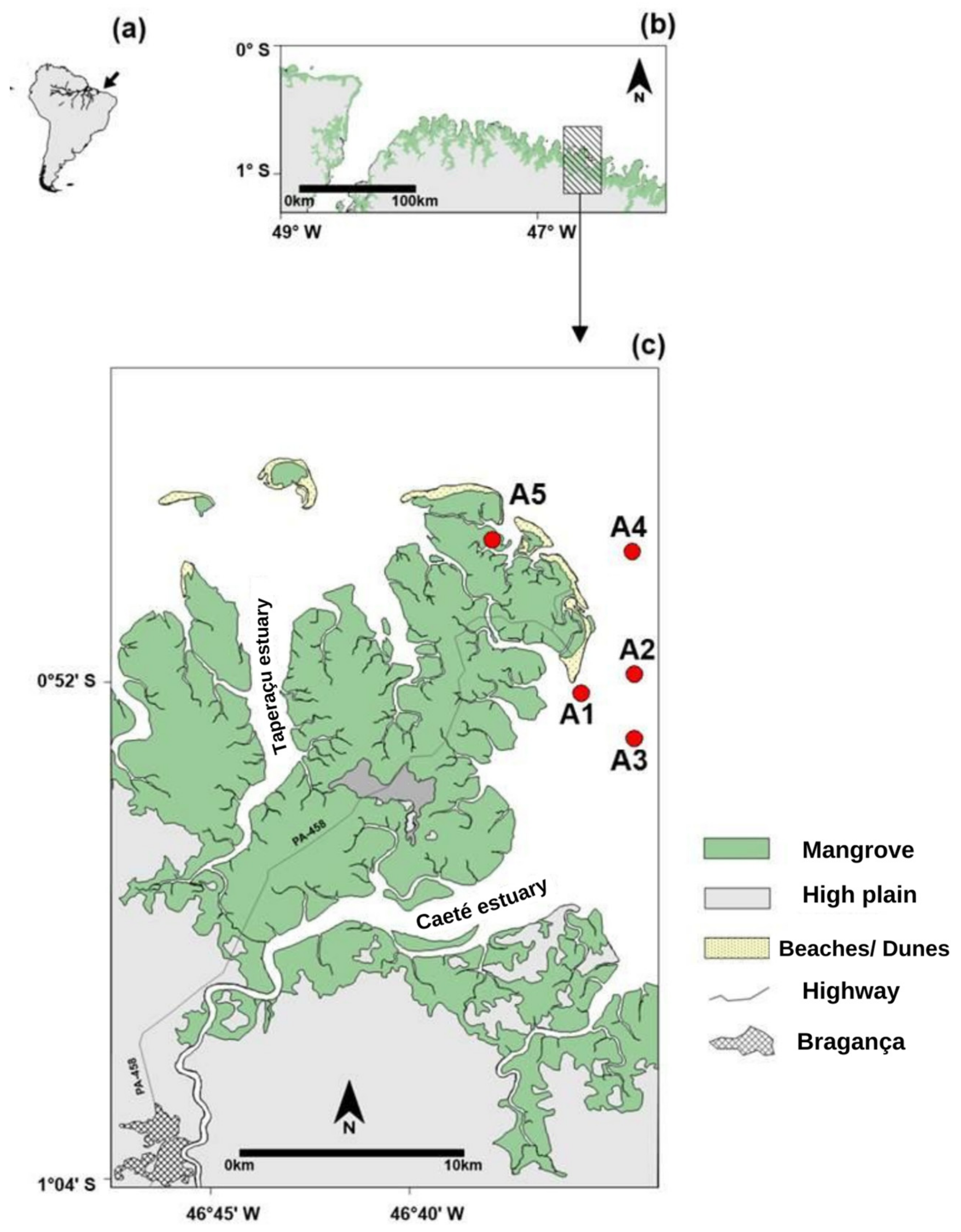
2. Materials and Methods
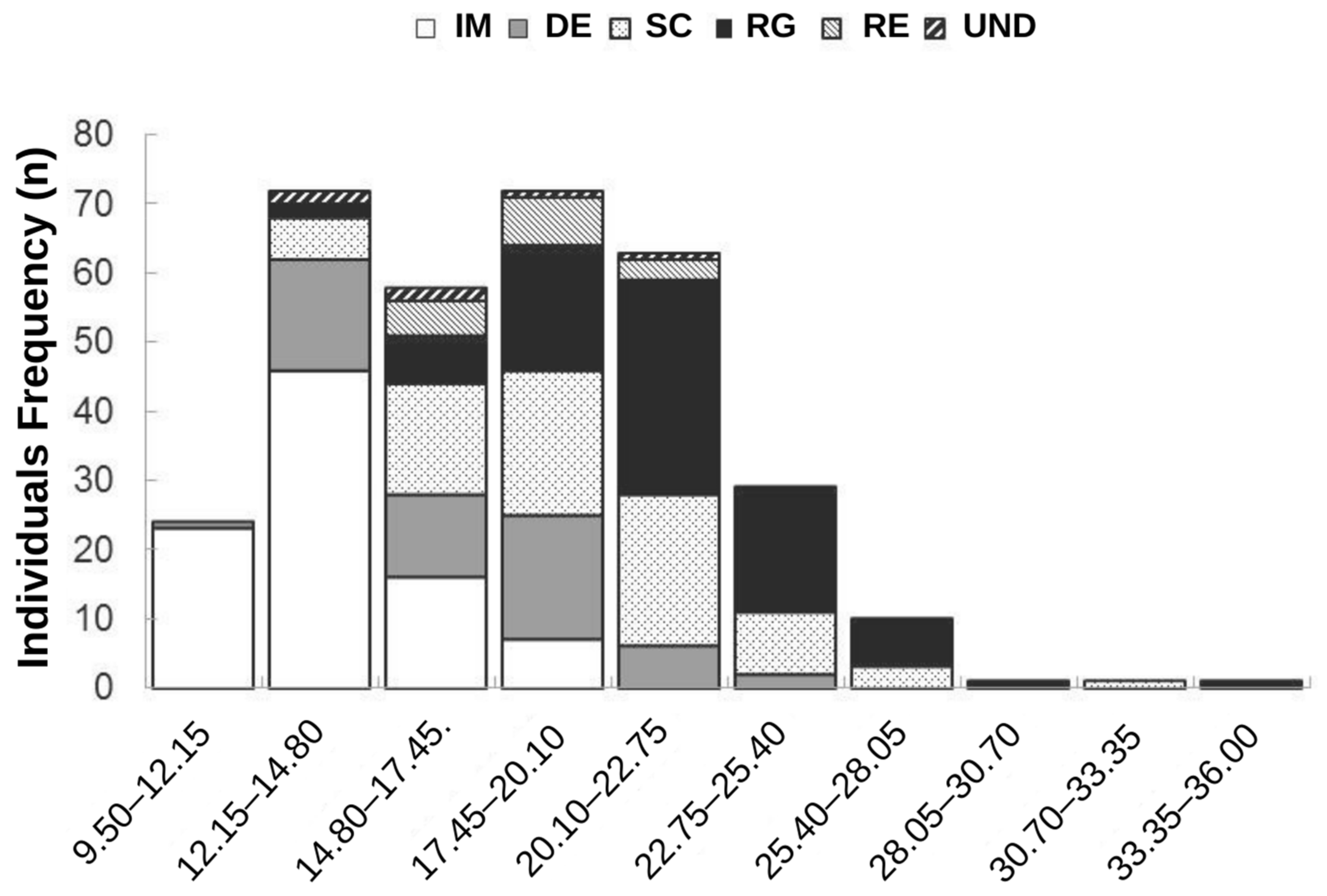
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winemiller, K.O. Life history strategies, population regulation, and implications for fisheries management. Can. J. Fish. Aquat. Sci. 2005, 62, 872–885. [Google Scholar] [CrossRef]
- Schreck, C.B. Stress and fish reproduction: The roles of allostasis and hormesis. Gen. Comp. Endocrinol. 2010, 165, 549–556. [Google Scholar] [CrossRef]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- Fonteles Filho, A.A. Oceanografia, Biologia e Dinâmica Populacional de Recursos Pesqueiros; Expressão Gráfica e Editora: Fortaleza, Brazil, 2011. [Google Scholar]
- Barletta-Bergan, A.; Barletta, M.; Saint-Paul, U. Structure and Seasonal Dynamics of Larval Fish in the Caeté River Estuary in North Brazil; Estuarine, Coastal and Shelf Science: Bragança, PA, Brazil, 2002; Volume 54, pp. 193–206. [Google Scholar]
- Barletta-Bergan, A. An Evaluation of the Larval Fish Assemblage in a North Brazilian Mangrove Area. In Mangrove Dynamics and Management in North Brazil; Saint-Paul, U., Schneider, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 209–220. [Google Scholar]
- Giarrizzo, T.; Krumme, U. Spatial differences and seasonal cyclicity in the intertidal fish fauna from four mangrove creeks in a salinity zone of the Curuçá estuary, north Brazil. Bull. Mar. Sci. 2007, 80, 739–754. [Google Scholar]
- Barletta, M.; Saint-Paul, U. Distribution pattern of fish in a mangrove estuary. In Mangrove Dynamics and Management in North Brazil; Saint-Paul, U., Schneider, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 171–208. [Google Scholar]
- Fernandes, S.C.P.; Bentes, A.B.; Pereira, L.D.J.G.; Nascimento, M.S.; Bentes, B. Variação temporal da captura comercial do peixe pedra, Genyatremus luteus, desembarcado em um pólo pesqueiro da costa norte do Brasil–Península de Ajuruteua–Bragança-PA. Bol. Inst. Pesca 2015, 41, 173–182. [Google Scholar]
- Fernandes, S.C.P.; Pereira, L.D.J.G.; Nascimento, M.S.; Andrade, C.E.R.; Bentes, B. Biologia Populacional do peixe pedra, Genyatremus luteus (BLOCH, 1790), costa amazônica brasileira. Bol. Inst. Pesca 2017, 43, 527–541. [Google Scholar] [CrossRef]
- Lindeman, K.C.; Richards, W.J. Haemulidae: Grunts. In Early Stages of Atlantic Fishes: An Identification Guide for Western Central North Atlantic; Richards, W.J., Ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 1597–1646. [Google Scholar]
- Maneschy, M.C. Pescadores curralistas no litoral do estado do Pará: Evolução e continuidade de uma pesca tradicional. Rev. Soc. Bras. Hist. Ciênc. 1993, 10, 53–73. [Google Scholar]
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; Macewicz, B.J.; Lowerre-Barbieri, S.K. A Standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. Dyn. Manag. Ecosyst. Sci. 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Moraes, B.C.; Costa, J.M.N.; Costa, A.C.L.; Costa, M.H. Variação espacial e temporal da precipitação no estado do Pará. Acta Amaz. 2005, 35, 207–217. [Google Scholar] [CrossRef]
- King, M. Fisheries Biology, Assessment and Management; Fishing News Books: Oxford, UK, 1995. [Google Scholar]
- Le Cren, E.D. The length-weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–219. [Google Scholar] [CrossRef]
- Vazzoler, A.E.A.M. Biologia da Reprodução de Peixes Teleósteos: Teoria e Prática; Eduem: Maringá, Brazil, 1996. [Google Scholar]
- Froese, R.; Tsikliras, A.C.; Stergiou, K.I. 2011: Editorial note on weight–length relations of fishes. Acta Ichthyol. Piscat. 2011, 41, 261–263. [Google Scholar] [CrossRef]
- Haimovici, M.; Canziani, G.V. Length-weight relationship of marine fishes from Southern Brazil. Environ. Sci. 2000, 23, 19–23. [Google Scholar]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Elliott, M.; Whitfield, A.K.; Potter, I.C.; Blaber, S.J.; Cyrus, D.P.; Nordlie, F.G.; Harrison, T.D. The guild approach to categorizing estuarine fish assemblages: A global review. Fish Fish. 2007, 8, 241–268. [Google Scholar] [CrossRef]
- Andrade-Tubino, M.F.; Ribeiro, A.L.R.; Vianna, M. Organização espaço-temporal das ictiocenoses demersais nos ecossistemas estuarinos brasileiros: Uma síntese. Oecol. Aust. 2009, 12, 640–661. [Google Scholar] [CrossRef]
- Auth, T.D.; Daly, E.A.; Brodeur, R.D.; Fisher, J.L. Phenological and distributional shifts in ichthyoplankton associated with recent warming in the northeast Pacific Ocean. Glob. Chang. Biol. 2018, 24, 259–272. [Google Scholar] [CrossRef]
- Mavruk, S.; Bengil, F.; Yüksek, A.; Özyurt, C.E.; Kiyağa, V.B.; Avşar, D. Intra-annual patterns of coastal larval fish assemblages along environmental gradients in the northeastern Mediterranean. Fish. Oceanogr. 2017, 27, 232–245. [Google Scholar] [CrossRef]
- Jalabert, B. Particularities of reproduction and oogenesis in teleost fish compared to mammals. Reprod. Nutr. Dev. 2005, 45, 261–279. [Google Scholar] [CrossRef]
- Noleto-Filho, E.M.; Yauri, W.L.M.; Luvizotto-Santos, R. Captura de reprodutores de peixe-pedra Genyatremus luteus (Bloch, 1797) (Pisces: Haemulidae) e manutenção em sistema fechado. Bol. Lab. Hidrobiol. 2012, 1, 55–60. [Google Scholar]
- Nikolskii, G.V. The Ecology of Fishes; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Johannes, R.E. Reproductive strategies of coastal marine fishes in the tropics. Environ. Biol. Fishes 1978, 3, 65–84. [Google Scholar] [CrossRef]
- Gómez, G.; Guzman, R.; Chacon, R. Algunos aspectos de La biologia reproductiva y poblacional del torroto, Genyatremus luteus, (Block, 1797) (Pisces: Haemulidae) em el golfo de Parla, Venezuela. Zootec. Trop. 2002, 20, 223–234. [Google Scholar]
- Marques, N.E.F.; Sampaio, J.A.A.; Bezerra, S.F.B.; Yauri, W.M.; Santos, R.L. Indução hormonal à desova do peixe pedra Genyatremus luteus (Bloch, 1790)(Teleostei: Haemulidae). Bol. Lab. Hidrobiol. 2016, 26, 32–35. [Google Scholar]
- Moura, H.T.G.S.; Nunes, Z.M.P. Caracterização sazonal das águas do sistema estuarino do Caeté (Bragança-PA). Bol. Inst. Pesca 2016, 42, 844–854. [Google Scholar] [CrossRef]
- Oliveira, S.M.O.; Pereira, L.C.C.; Vila-Concejo, A. Processos morfodinâmicos em uma praia de macromaré no litoral amazônico. Quat. Environ. Geosci. 2014, 5, 125–136. [Google Scholar] [CrossRef]
- Silva, A.M.; Oliveira, M.R.; Chellappa, S. Biologia reprodutiva do coró, Pomadasys corvinaeformis Steindachner (Osteichthyes: Haemulidae) das águas costeiras do Rio Grande do Norte, Brasil. Biota Amaz. 2012, 2, 15–24. [Google Scholar] [CrossRef]
- Shatunovskiy, M.I. Some indices of reproductive ability of fishes. J. Ichthyol. 1988, 28, 119–123. [Google Scholar]
- Araújo, B.R. Desova e fecundidade em peixes de água doce e marinhos. Rev. Biol. Ciênc. Terra 2009, 9, 24–31. [Google Scholar]
- Wooton, R.J. Ecology of Teleost Fishes; Chapman and Hall: London, UK, 1990. [Google Scholar]
- Domínguez-Petit, R.; Korta, M.; Saborido-Rey, F.; Murua, H.; Sainza, M.; Piñeiro, C. Changes in size at maturity of European hake Atlantic populations in relation with stock structure and environmental regimes. J. Mar. Syst. 2008, 71, 260–278. [Google Scholar] [CrossRef]
- Lojo, D.; Cousido-Rocha, M.; Cerviño, S.; Dominguez-Petit, R.; Sainza, M.; Pennino, M.G. Assessing changes in size at maturity for the European hake (Merluccius merluccius) in Atlantic Iberian waters. Sci. Mar. 2022, 86, e046. [Google Scholar] [CrossRef]
- Lima, W.M.G.D.; Pinaya, W.H.D.; Lutz ÍAD, F.; Nascimento, M.S.D.; Silva, L.E.O.D.; Cintra, I.H.A.; Bentes, B. Effects of the trawling industrial fishery system on fish community structure on the inner Amazon shelf. Ocean. Coast. Res. 2021, 69, 9–10. [Google Scholar] [CrossRef]
- De Souza-Araujo, J.; Hussey, N.E.; Hauser-Davis, R.A.; Rosa, A.H.; de Oliveira Lima, M.; Giarrizzo, T. Human risk assessment of toxic elements (As, Cd, Hg, Pb) in marine fish from the Amazon. Chemosphere 2022, 301, 134575. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month/Year | Sex | N | Total Length (cm) | Weight (g) | ||
|---|---|---|---|---|---|---|
| Min–Max | Average ± SD | Min–Max | Average ± SD | |||
| Apr/16 | F | 6 | 18.20–35.40 | 23.38 ± 5.66 | 114.00–771.00 | 277.67 ± 224.40 |
| M | 4 | 13.90–22.90 | 17.75 ± 3.70 | 47.00–198.00 | 112.75 ± 64.01 | |
| May/16 | F | 22 | 17.60–31.90 | 23.23 ± 3.21 | 88.00–549.00 | 235.73 ± 100.53 |
| M | 14 | 14.80–23.20 | 19.16 ± 1.90 | 74.00–233.00 | 138.43 ± 43.84 | |
| Jun/16 | F | 13 | 10.40–24.60 | 20.78 ± 4.20 | 22.00–302.00 | 194.46 ± 79.08 |
| M | 5 | 9.50–21.90 | 17.18 ± 4.39 | 16.00–188.00 | 109.80 ± 65.36 | |
| UND | 1 | 11.2 | - | 23 | - | |
| Jul/16 | F | 15 | 16.50–25.70 | 20.33 ± 2.31 | 78.00–315.00 | 159.60 ± 62.14 |
| M | 11 | 14.50–20.10 | 17.28 ± 1.86 | 51.00–127.00 | 91.73 ± 24.51 | |
| UND | 1 | 13.6 | - | 46 | - | |
| Aug/16 | F | 17 | 11.80–27.40 | 18.62 ± 4.29 | 28.00–371.00 | 135.71 ± 93.58 |
| M | 22 | 10.80–20.10 | 15.49 ± 2.90 | 22.00–147.00 | 72.95 ± 39.77 | |
| UND | 1 | 10.1 | - | 19 | - | |
| Sep/16 | F | 30 | 11.60–22.50 | 16.55 ± 2.73 | 26.00–206.00 | 84.40 ± 41.81 |
| M | 30 | 11.10–21.20 | 16.00 ± 3.21 | 25.00–180.00 | 81.73 ± 47.59 | |
| UND | 6 | 10.40–18.60 | 13.27 ± 3.11 | 18.00–114.00 | 43.50 ± 33.95 | |
| Oct/16 | F | 10 | 13.20–25.80 | 20.15 ± 3.97 | 41.00–289.00 | 152.20 ± 79.11 |
| M | 17 | 11.30–22.60 | 16.26 ± 4.44 | 25.00–190.00 | 91.59 ± 65.32 | |
| UND | 6 | 10.10–12.90 | 11.44 ± 1.00 | 20.00–45.00 | 31.40 ± 9.26 | |
| Nov/16 | F | - | - | - | - | - |
| M | - | - | - | - | - | |
| Dec/16 | F | 14 | 12.60–23.70 | 16.55 ± 3.50 | 36.00–211.00 | 90.50 ± 59.78 |
| M | 17 | 12.00–24.60 | 17.69 ± 3.86 | 32.00–252.00 | 109.53 ± 69.12 | |
| UND | 1 | 16.2 | - | 65 | - | |
| Jan/17 | F | 17 | 13.60–28.60 | 21.55 ± 3.64 | 42.00–385.00 | 190.29 ± 86.59 |
| M | 8 | 11.50–21.40 | 16.28 ± 3.37 | 28.00–151.00 | 85.38 ± 40.27 | |
| Feb/17 | F | - | - | - | - | - |
| M | - | - | - | - | - | |
| Mar/17 | F | 13 | 12.40–19.10 | 15.37 ± 2.16 | 32.00–120.00 | 68.92 ± 28.11 |
| M | 5 | 12.50–17.90 | 15.54 ± 1.92 | 35.00–98.00 | 68.00 ± 25.18 | |
| UND | 2 | 13.40–14.00 | 13.70 ± 0.30 | 45.00–49.00 | 47.00 ± 2.00 | |
| Apr/17 | F | 17 | 14.10–25.40 | 18.56 ± 3.27 | 47.00–305.00 | 122.41 ± 69.03 |
| M | 7 | 13.00–17.80 | 15.37 ± 1.67 | 37.00–106.00 | 65.29 ± 22.79 | |
| Total | F | 174 | 10.40–35.40 | 19.28 ± 4.28 | 22.00–771.00 | 147.21 ± 101.50 |
| M | 140 | 9.50–24.60 | 16.63 ± 3.41 | 16.00–252.00 | 92.14 ± 53.82 | |
| UND | 17 | 10.10–18.60 | 12.66 ± 2.39 | 18.00–114.00 | 39.12 ± 23.35 | |
| Total individuals | 331 | 9.50–35.40 | 17.82 ± 4.23 | 16.00–771.00 | 118.37 ± 87.86 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, S.; Pinheiro, L.; Bentes, B. Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf. Fishes 2023, 8, 181. https://doi.org/10.3390/fishes8040181
Fernandes S, Pinheiro L, Bentes B. Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf. Fishes. 2023; 8(4):181. https://doi.org/10.3390/fishes8040181
Chicago/Turabian StyleFernandes, Suelly, Larissa Pinheiro, and Bianca Bentes. 2023. "Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf" Fishes 8, no. 4: 181. https://doi.org/10.3390/fishes8040181