
Effect of Simulated Organic–Inorganic N Deposition on Leaf Stoichiometry, Chlorophyll Content, and Chlorophyll Fluorescence in Torreya grandis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. ON and IN Addition Experiment
2.3. Leaf Stoichiometry Measurement
2.4. Chlorophyll Measurement
2.5. Chlorophyll Fluorescence Measurement
2.6. Statistical Analyses
3. Results
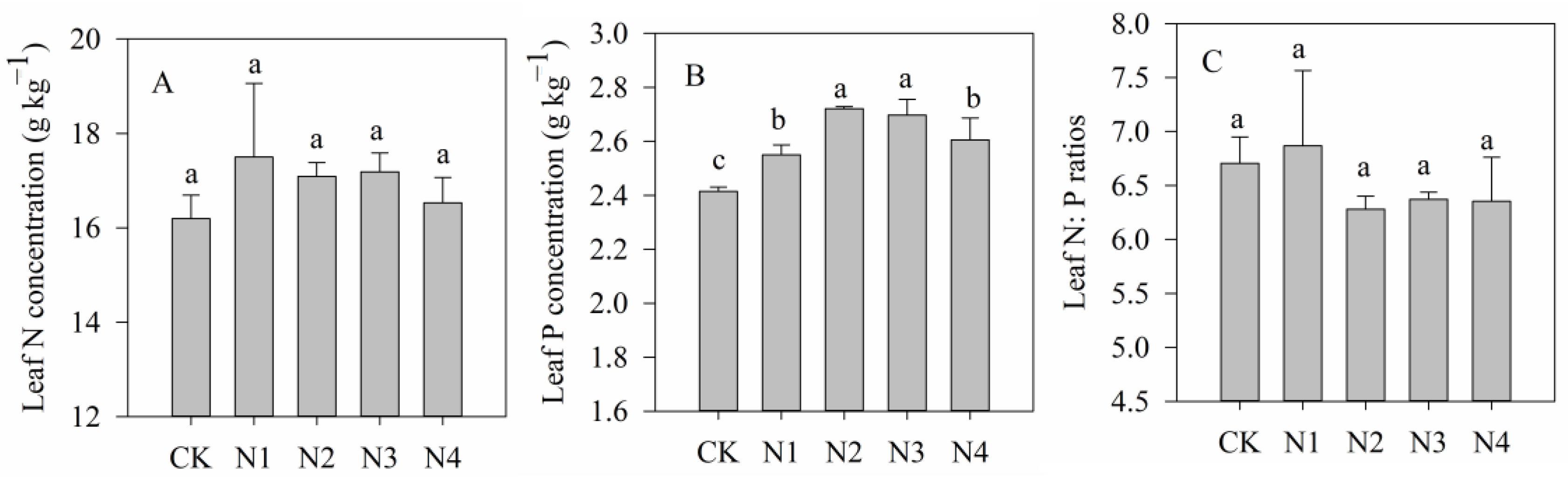
3.1. Foliar Stoichiometry Characteristics
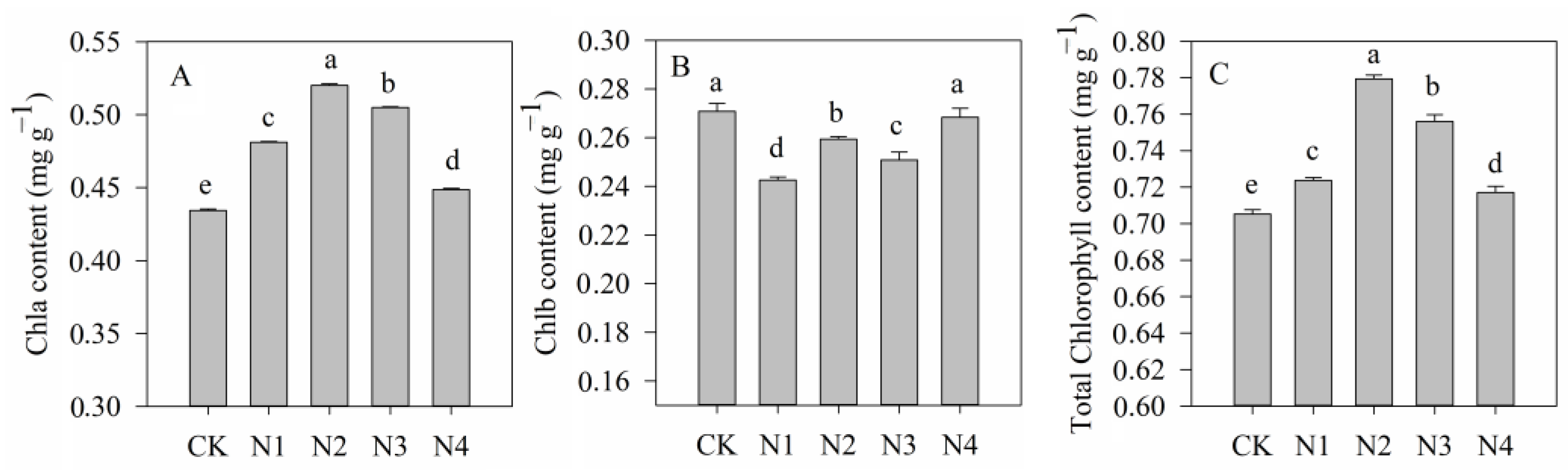
3.2. Chlorophyll Characteristics
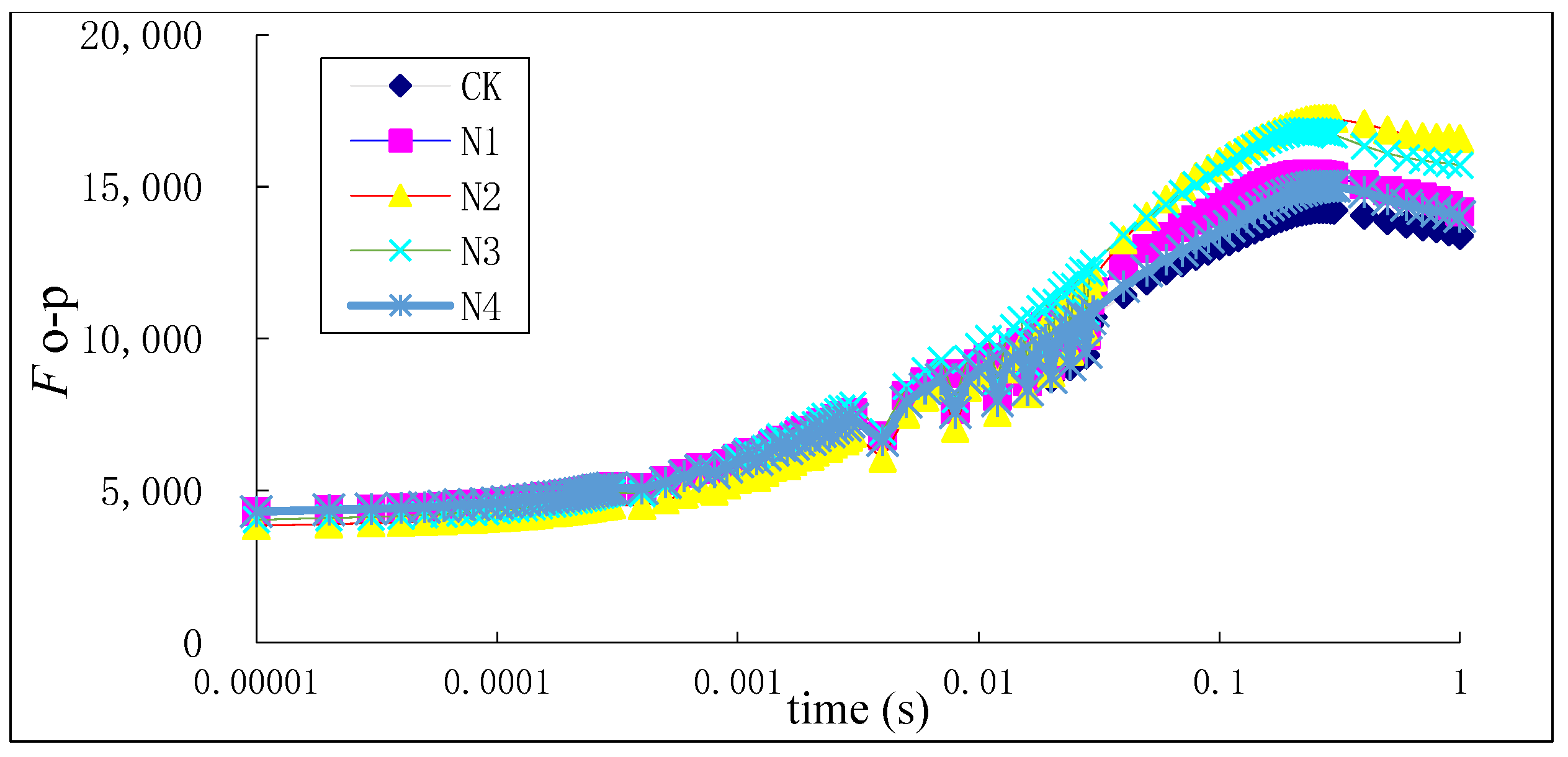
3.3. Chlorophyll Fluorescence Characteristics
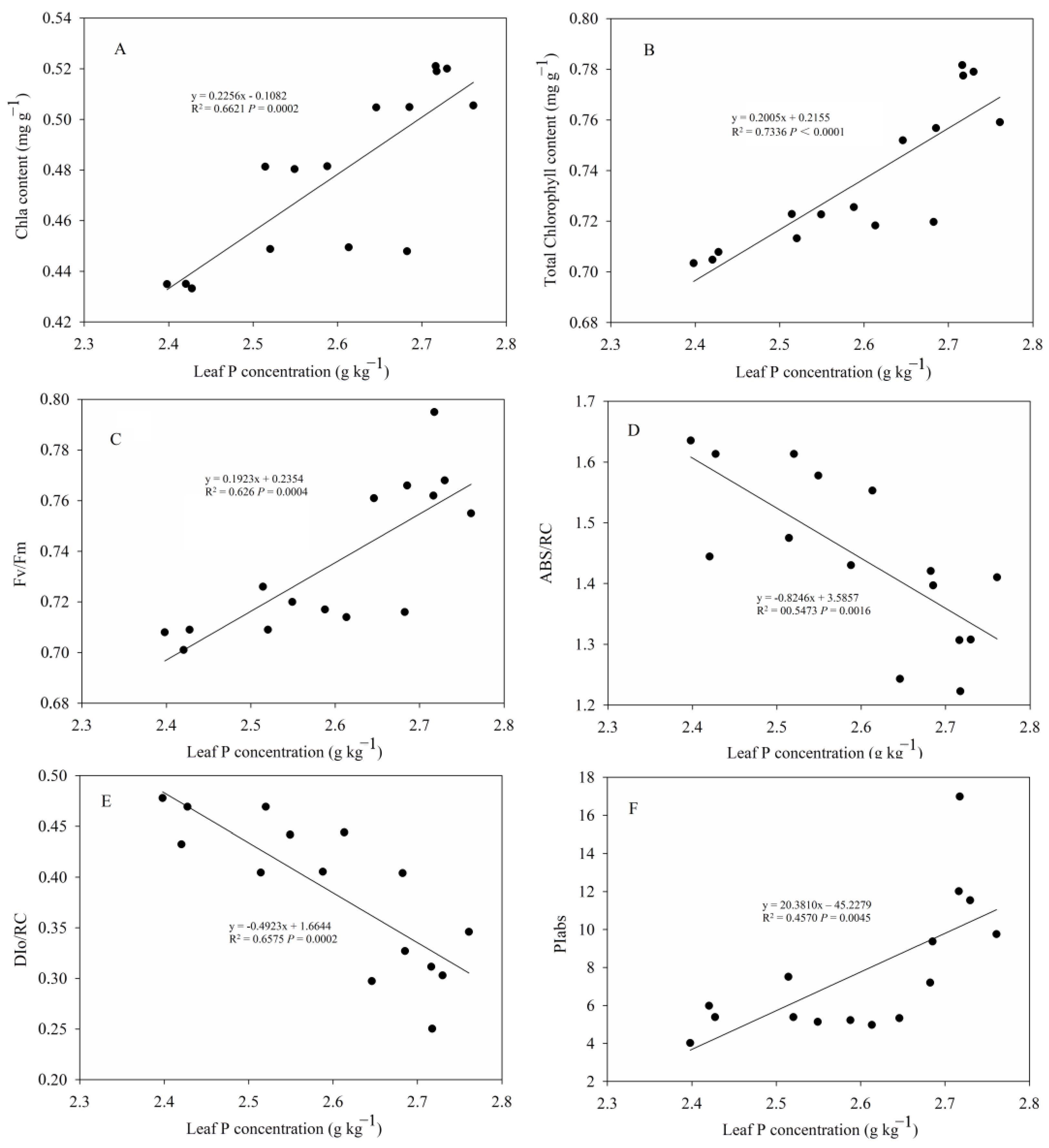
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, X.; Zhang, Y.; Han, W. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Francini, A.; Sebastiani, L. Abiotic stress effects on performance of horticultural crops. Horticulturae 2019, 5, 67. [Google Scholar] [CrossRef]
- Chang, Y.; Liu, X.J.; Li, K.H. Research progress in atmospheric nitrogen deposition. Arid Zone Res. 2012, 29, 972–979. [Google Scholar]
- Chakraborty, A. Impact of 30 years precipitation regime differences on forest soil physiology and microbial assemblages. Front. For. Glob. Chang. 2023, 6, 1142979. [Google Scholar] [CrossRef]
- Song, X.; Gu, H.; Wang, M.; Zhou, G.; Li, Q. Management practices regulate the response of Moso bamboo foliar stoichiometry to nitrogen deposition. Sci. Rep. 2016, 6, 24107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wu, J.; Li, Q.; Hänninen, H.; Peng, C.; Yao, H.; Ying, Y. Nitrogen deposition enhances photosynthesis in Moso bamboo but increases susceptibility to other stress factors. Front. Plant Sci. 2017, 8, 1975. [Google Scholar] [CrossRef]
- Du, Y.; Guo, P.; Liu, J.; Wang, C.; Yang, N.; Jiao, Z. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Glob. Chang. Biol. 2014, 20, 3222–3228. [Google Scholar] [CrossRef]
- Lim, H.; Jämtgård, S.; Oren, R.; Gruffman, L.; Kunz, S.; Näsholm, T. Organic nitrogen enhances nitrogen nutrition and early growth of Pinus sylvestris seedlings. Tree Physiol. 2022, 42, 513–522. [Google Scholar] [CrossRef]
- Schreeg, L.A.; Santiago, L.S.; Wright, S.J. Stem, root, and older leaf N: P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology 2014, 95, 2062–2068. [Google Scholar] [CrossRef]
- Maaroufi, N.I.; De Long, J.R. Global change impacts on forest soils: Linkage between soil biota and carbon-nitrogen-phosphorus stoichiometry. Front. For. Glob. Chang. 2020, 3, 16. [Google Scholar] [CrossRef]
- Sardans, J.; Alonso, R.; Janssens, I.A. Foliar and soil concentrations and stoichiometry of nitrogen and phosphorous across european Pinus Sylvestris forests: Relationships with climate, N deposition and tree growth. Functional. Ecol. 2016, 30, 676–689. [Google Scholar] [CrossRef]
- Guo, P.; Wang, C.; Feng, X. Mixed inorganic and organic nitrogen addition enhanced extracellular enzymatic activities in a subtropical forest soil in east China. Water Air Soil Pollut. 2011, 216, 229–237. Available online: https://link.springer.com/article/10.1007/s11270-010-0530-x (accessed on 9 July 2010). [CrossRef]
- Guo, P.; Han, T.; Zhang, L. Changes of soil bacterial activities and functions after different N additions in a temperate forest. Environ. Sci. Pollut. Res. 2017, 24, 3853–3860. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Mao, Q.; Gilliam, F.S. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Niu, S. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10, 024019. Available online: https://iopscience.iop.org/article/10.1088/1748-9326/10/2/024019/meta (accessed on 20 February 2015). [CrossRef]
- Holzmann, S.; Missong, A.; Puhlmann, H. Impact of anthropogenic induced nitrogen input and liming on phosphorus leaching in forest soils. J. Plant Nutr. Soil Sci. 2016, 179, 443–453. [Google Scholar] [CrossRef]
- Zhang, H.; Ge, Y.; Xie, X. High throughput analysis of leaf chlorophyll content in sorghum using RGB, hyperspectral, and fluorescence imaging and sensor fusion. Plant Methods 2022, 18, 60. [Google Scholar] [CrossRef]
- Chen, M. Chlorophyll modifications and their spectral extension in oxygenic photosynthesis. Annu. Rev. Biochem. 2014, 83, 317–340. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M. Leaf pigment content. Compr. Remote Sens. 2018, 3, 117–142. [Google Scholar]
- Liu, Y.; Wang, T.; Fang, S.; Zhou, M.; Qin, J. Responses of morphology, gas exchange, photochemical activity of photosystem II, and antioxidant balance in Cyclocarya paliurus to light spectra. Front. Plant Sci. 2018, 9, 1704. [Google Scholar] [CrossRef]
- Sage, R.F.; Pearcy, R.W.; Seemann, J.R. The nitrogen use efficiency of C3 and C4 plants: III. Leaf nitrogen effects on the activity of carboxylating enzymes in Chenopodium album (L.) and Amaranthus retroflexus (L.). Plant Physiol. 1987, 85, 355–359. [Google Scholar] [CrossRef]
- Hua, Q.; Yu, Y.; Dong, S. Leaf spectral responses of Poa crymophila to nitrogen deposition and climate change on Qinghai-Tibetan plateau. Agric. Ecosyst. Environ. 2019, 284, 106598. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Murata, N. Environmental stress inhibits the synthesis de novo of proteins involved in the photodamage–repair cycle of photosystem II in Synechocystis sp. PCC 6803. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2004, 1657, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Lee, J.G. Effect of drought stress on chlorophyll fluorescence parameters, phytochemical contents, and antioxidant activities in lettuce seedlings. Horticulturae 2021, 7, 238. [Google Scholar] [CrossRef]
- Zhang, J.; Lv, J.; Li, Q. Effects of nitrogen deposition and management practices on leaf litterfall and N and P return in a Moso bamboo forest. Biogeochemistry 2017, 134, 115–124. Available online: https://link.springer.com/article/10.1007/s10533-017-0349-2 (accessed on 2 June 2017). [CrossRef]
- Li, Y.; Wu, X.Q. Factors Influencing Leaf Chlorophyll Content in Natural Forests at the Biome Scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, Z.; Ye, Y.; Wang, D.; Jin, S. Enhanced salt tolerance of Torreya grandis genders is related to nitric oxide level and antioxidant capacity. Front. Plant Sci. 2022, 13, 906071. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Xu, Y.; Chen, W.; Han, Y.; Wang, G.G.; Jin, S. Sexual differences in gas exchange and chlorophyll fluorescence of Torreya grandis under drought stress. Trees 2022, 36, 283–294. [Google Scholar] [CrossRef]
- Zhang, R.; Zhao, Y.X.; Lin, J.H. Biochar application alleviates unbalanced nutrient uptake caused by N deposition in Torreya grandis trees and seedlings. For. Ecol. Manag. 2019, 432, 319–326. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, P.; Chen, S.; Hang, Z.; Jin, S. Simulated organic–inorganic nitrogen deposition changes the growth rate, leaf stoichiometry, and phenolic content of Cyclocarya Paliurus. For. Ecol. Manag. 2023, 546, 121313. [Google Scholar] [CrossRef]
- Wu, T.; Dong, Y.; Yu, M.; Wang, G.G. Leaf nitrogen and phosphorus stoichiometry of Quercus species across China. For. Ecol. Manag. 2012, 284, 116–123. [Google Scholar] [CrossRef]
- Bukhat, S.; Manzoor, H.; Athar, H.R. Salicylic acid induced photosynthetic adaptability of Raphanus sativus to salt stress is associated with antioxidant capacity. J. Plant Growth Regul. 2020, 39, 809–822. Available online: https://link.springer.com/article/10.1007/s00344-019-10024-z (accessed on 24 September 2019). [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Cessna, S.; Demmig-Adams, B.; Adams, W.W., III. Exploring photosynthesis and plant stress using inexpensive chlorophyll fluorometers. J. Nat. Resour. Life Sci. Educ. 2010, 39, 22–30. [Google Scholar] [CrossRef]
- Smil, V. Nitrogen in crop production: An account of global flows. Glob. Biogeochem. Cycles 1999, 13, 647–662. [Google Scholar] [CrossRef]
- Güsewell, S. N: P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Greenwood, D.J.; Karpinets, T.V.; Zhang, K. A unifying concept for the dependence of whole-crop N: P ratio on biomass: Theory and experiment. Ann. Bot. 2008, 102, 967–977. [Google Scholar] [CrossRef]
- Silver, W.L.; Thompson, A.W.; Reich, A. Nitrogen cycling in tropical plantation forests: Potential controls on nitrogen retention. Ecol. Appl. 2005, 15, 1604–1614. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, Y.; Wang, J. Effects of long-term organic–inorganic nitrogen application on maize yield and nitrogen-containing gas emission. Agronomy 2023, 13, 848. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Bowles, T.M.; Atallah, S.S.; Campbell, E.E. Addressing agricultural nitrogen losses in a changing climate. Nat. Sustain. 2018, 1, 399–408. Available online: https://www.nature.com/articles/s41893-018-0106-0 (accessed on 14 August 2018). [CrossRef]
- Huang, J.; Duan, Y.; Xu, M. Nitrogen mobility, ammonia volatilization, and estimated leaching loss from long-term manure incorporation in red soil. J. Integr. Agric. 2017, 16, 2082–2092. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Y.; Song, L.; Song, X.; Hänninen, H.; Wu, J. Biochar enhances nut quality of Torreya grandis and soil fertility under simulated nitrogen deposition. For. Ecol. Manag. 2017, 391, 321–329. [Google Scholar] [CrossRef]
- Lv, F.; Song, J.; Giltrap, D. Crop yield and N2O emission affected by long-term organic manure substitution fertilizer under winter wheat-summer maize cropping system. Sci. Total Environ. 2020, 732, 139321. [Google Scholar] [CrossRef]
- Southon, G.E.; Field, C.; Caporn, S.; Britton, A.; Power, S. Nitrogen deposition reduces plant diversity and alters ecosystem functioning: Field-scale evidence from a nationwide survey of UK heathlands. PLoS ONE 2013, 8, e59031. [Google Scholar] [CrossRef]
- Yu, Q.; Elser, J.J.; He, N. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia 2011, 166, 1–10. [Google Scholar] [CrossRef]
- Long, M.; Wu, H.H.; Smith, M.D. Nitrogen deposition promotes phosphorus uptake of plants in a semi-arid temperate grassland. Plant Soil 2016, 408, 475–484. Available online: https://link.springer.com/article/10.1007/s11104-016-3022-y (accessed on 20 August 2016). [CrossRef]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. Available online: https://link.springer.com/article/10.1007/s11104-011-0950-4 (accessed on 3 September 2011). [CrossRef]
- Naqvi, K.R.; Jávorfi, T.; Melø, T.B. More on the catalysis of internal conversion in chlorophyll a by an adjacent carotenoid in light-harvesting complex (Chla/b LHCII) of higher plants: Time-resolved triplet-minus-singlet spectra of detergent-perturbed complexes. Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 1998, 55, 193–204. [Google Scholar] [CrossRef]
- Cai, J.G.; Wei, M.Q.; Zhang, Y. Effects of shading on photosynthetic characteristics and chlorophyll fluorescence parameters in leaves of Hydrangea macrophylla. Chin. J. Plant Ecol. 2017, 41, 570–576. [Google Scholar]
- He, J.; Chee, C.W.; Goh, C.J. ‘Photoinhibition’of Heliconia under natural tropical conditions: The importance of leaf orientation for light interception and leaf temperature. Plant Cell Environ. 1996, 19, 1238–1248. [Google Scholar] [CrossRef]
- Ma, J.; Sun, M.; Qiu, L. The 5-Aminolevulinic Acid (5-ALA) Supplement enhances PSII photochemical activity and antioxidant activity in the late growth promotion of Pseudostellaria heterophylla. Plants 2022, 11, 3035. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fv/Fm | PIabs | ABS/RC | DIo/RC | TRo/RC | DIo/CSm |
|---|---|---|---|---|---|---|
| CK | 0.71 ± 0.00 b | 5.19 ± 1.00 b | 1.56 ± 0.10 a | 0.46 ± 0.02 a | 1.10 ± 0.08 a | 4388.67 ± 549.19 a |
| N1 | 0.72 ± 0.00 b | 5.95 ± 1.34 b | 1.49 ± 0.08 ab | 0.42 ± 0.02 a | 1.08 ± 0.06 ab | 4306.33 ± 611.09 a |
| N2 | 0.76 ± 0.02 a | 13.51 ± 3.02 a | 1.28 ± 0.05 c | 0.29 ± 0.03 b | 0.99 ± 0.02 b | 3851.33 ± 572.06 a |
| N3 | 0.76 ± 0.01 a | 8.15 ± 2.45 b | 1.35 ± 0.09 bc | 0.32 ± 0.02 b | 1.03 ± 0.07 ab | 4035.00 ± 354.27 a |
| N4 | 0.71 ± 0.00 b | 5.85 ± 1.18 b | 1.53 ± 0.10 a | 0.44 ± 0.03 a | 1.09 ± 0.07 ab | 4317.00 ± 362.30 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Zhang, H.; Shi, X.; Han, Y.; Liu, Y.; Jin, S. Effect of Simulated Organic–Inorganic N Deposition on Leaf Stoichiometry, Chlorophyll Content, and Chlorophyll Fluorescence in Torreya grandis. Horticulturae 2023, 9, 1042. https://doi.org/10.3390/horticulturae9091042
Yuan Y, Zhang H, Shi X, Han Y, Liu Y, Jin S. Effect of Simulated Organic–Inorganic N Deposition on Leaf Stoichiometry, Chlorophyll Content, and Chlorophyll Fluorescence in Torreya grandis. Horticulturae. 2023; 9(9):1042. https://doi.org/10.3390/horticulturae9091042
Chicago/Turabian StyleYuan, Yichao, Haochen Zhang, Xianmeng Shi, Yini Han, Yang Liu, and Songheng Jin. 2023. "Effect of Simulated Organic–Inorganic N Deposition on Leaf Stoichiometry, Chlorophyll Content, and Chlorophyll Fluorescence in Torreya grandis" Horticulturae 9, no. 9: 1042. https://doi.org/10.3390/horticulturae9091042