
Species-Specific Level Variation in Polyamines in Coniferous and Deciduous Woody Plant Species in Urban Areas
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Characteristics and Sampling Strategy
Microclimate Characteristics
2.2. Polyamine Analysis
2.3. Determination of Antioxidant Capacity and Total Phenolic and Flavonoid Contents
- The ABTS assay was employed to assess total antioxidant capacity, which relies on monitoring the conversion of the cationic radical 2,2′-azinobis(3-ethylbenzothiozoline-6-sulfonic acid), ABTS•+ from a blue-green color to its neutral and colorless form at 734 nm. This procedure followed the methodology outlined by Miller and Rice-Evans [32].
- The DPPH-scavenging activity was assessed using the method described by Arnao [33]. This method relies on the transformation of purple DPPH• (2,2-diphenyl-1-picrylhydrazyl radical) into its reduced yellow form, DPPH-H, following a 30 min incubation at 30 °C in the absence of light. The absorbance was subsequently measured at 515 nm.
- The inhibition of the nitric oxide radical (NO•) was assessed via the Griess diazotization process, following the methodology devised by Hensley et al. [34]. The extent of inhibition was determined by measuring the absorbance of the resulting chromophore at 546 nm. This approach allowed for the quantification of the NO• inhibitory effects.
- The ferric reducing antioxidant power (FRAP) assay involves the reduction in FeIII –TPTZ (Iron (II)-2,4,5-tripyridyl-S-triazine) under low pH conditions, resulting in the formation of a blue-colored FeII–TPTZ complex. The measurement involves reading the absorbance of an intense blue complex at 593 nm, as detailed in a prior description published by Benzie and Strain [35].
- The total phenolic content (TPC) was quantified according to the Folin–Ciocalteu (FC) method described by Singleton et al. [36]. The method is based on the spectrophotometric detection of phenols forming a colored complex with a FC reagent. Absorbance was measured spectrophotometrically at 760 nm, and TPC was expressed as milligrams of gallic acid equivalents (GAE) per gram of DW (mg GAE g−1 DW), calculated according to the standard calibration curve.
- The total flavonoid content (TFC) was quantified spectrophotometrically, employing aluminum chloride (AlCl3) as the complexing reagent for flavonoids. Absorbance readings were taken at 415 nm, and quercetin was used as a standard for the calibration curve. The TFC value of the extract is presented as milligrams of quercetin equivalents (QE) per gram of DW (mg QE g−1 DW) [37].
2.4. Statistical Analysis
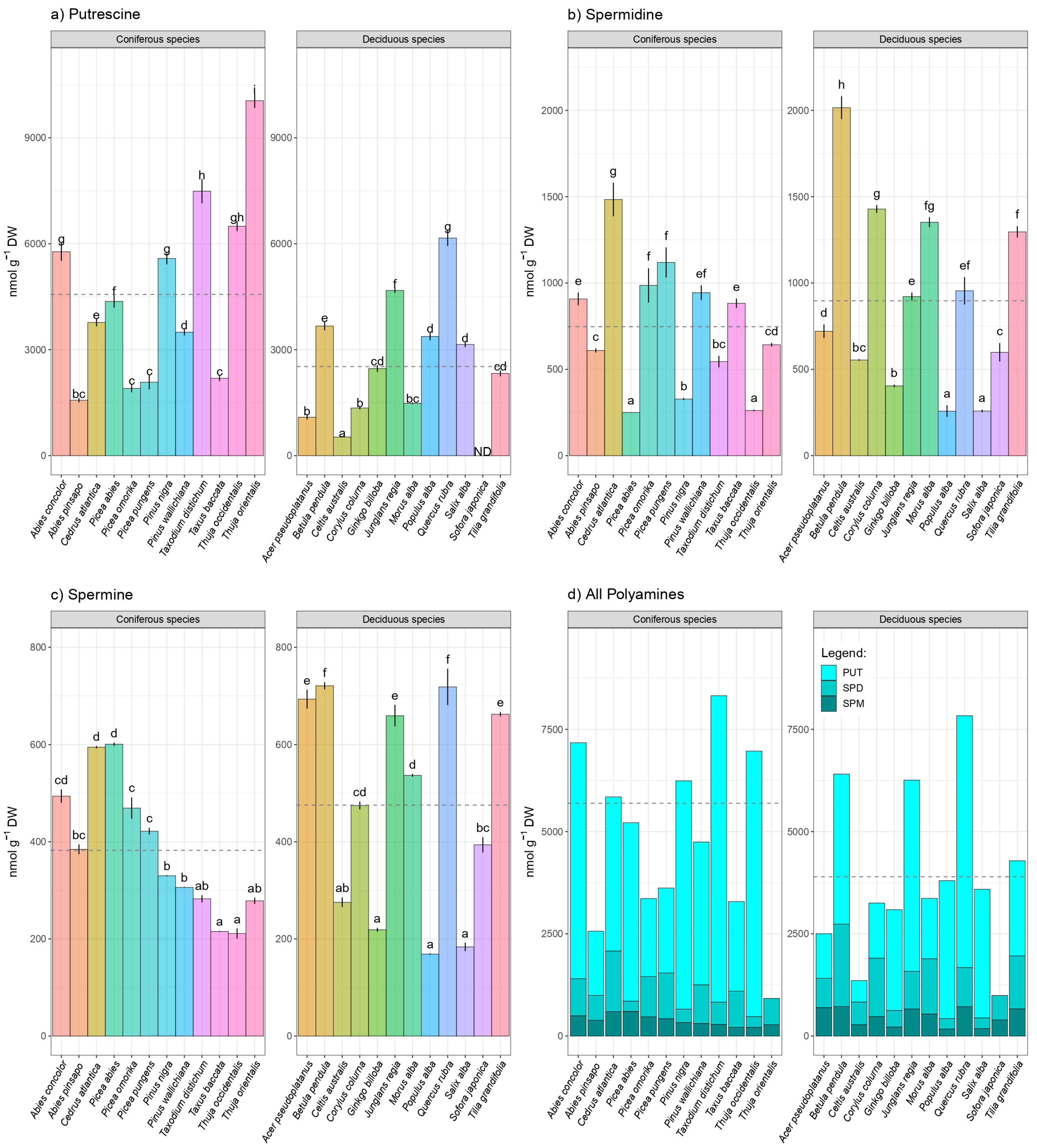
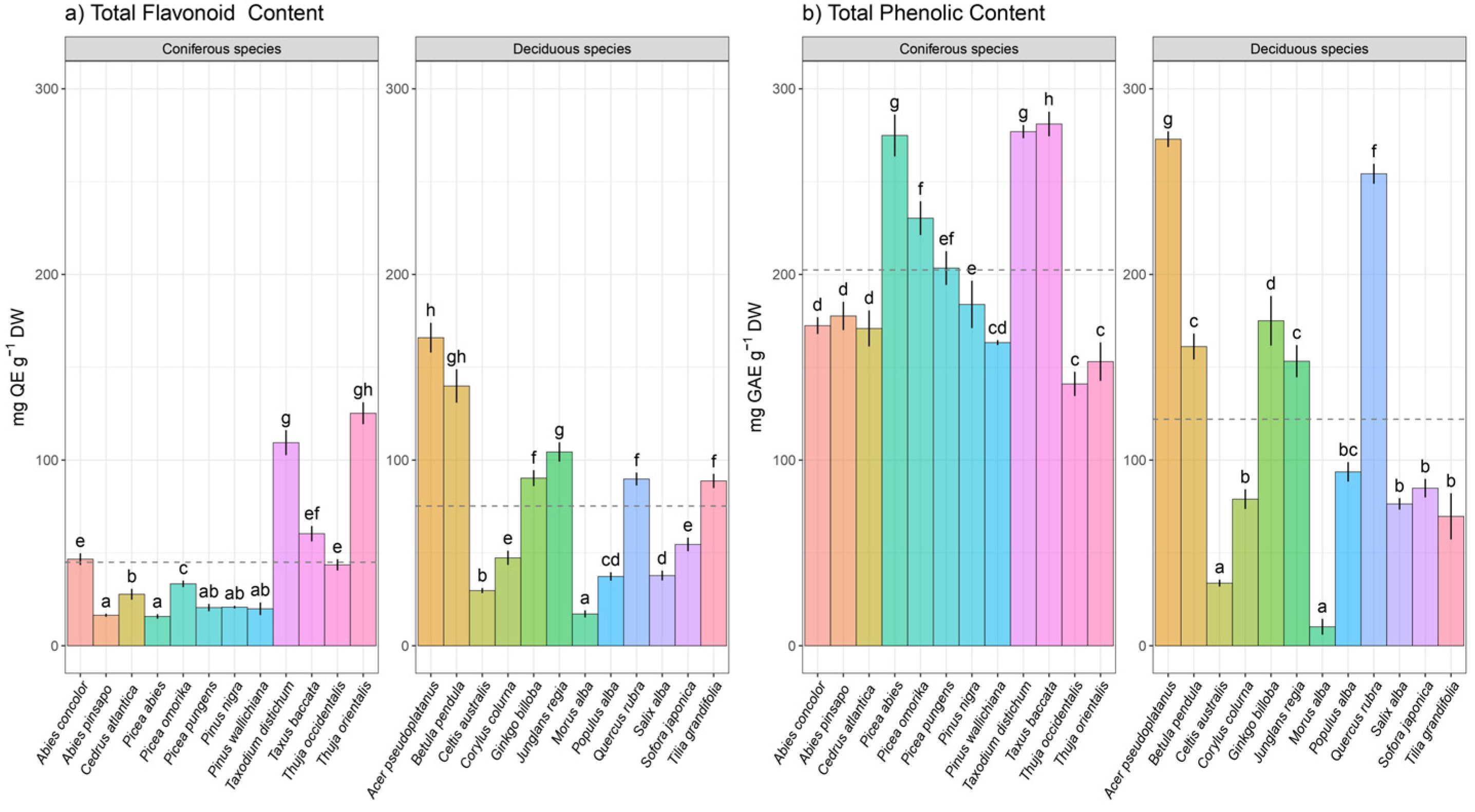
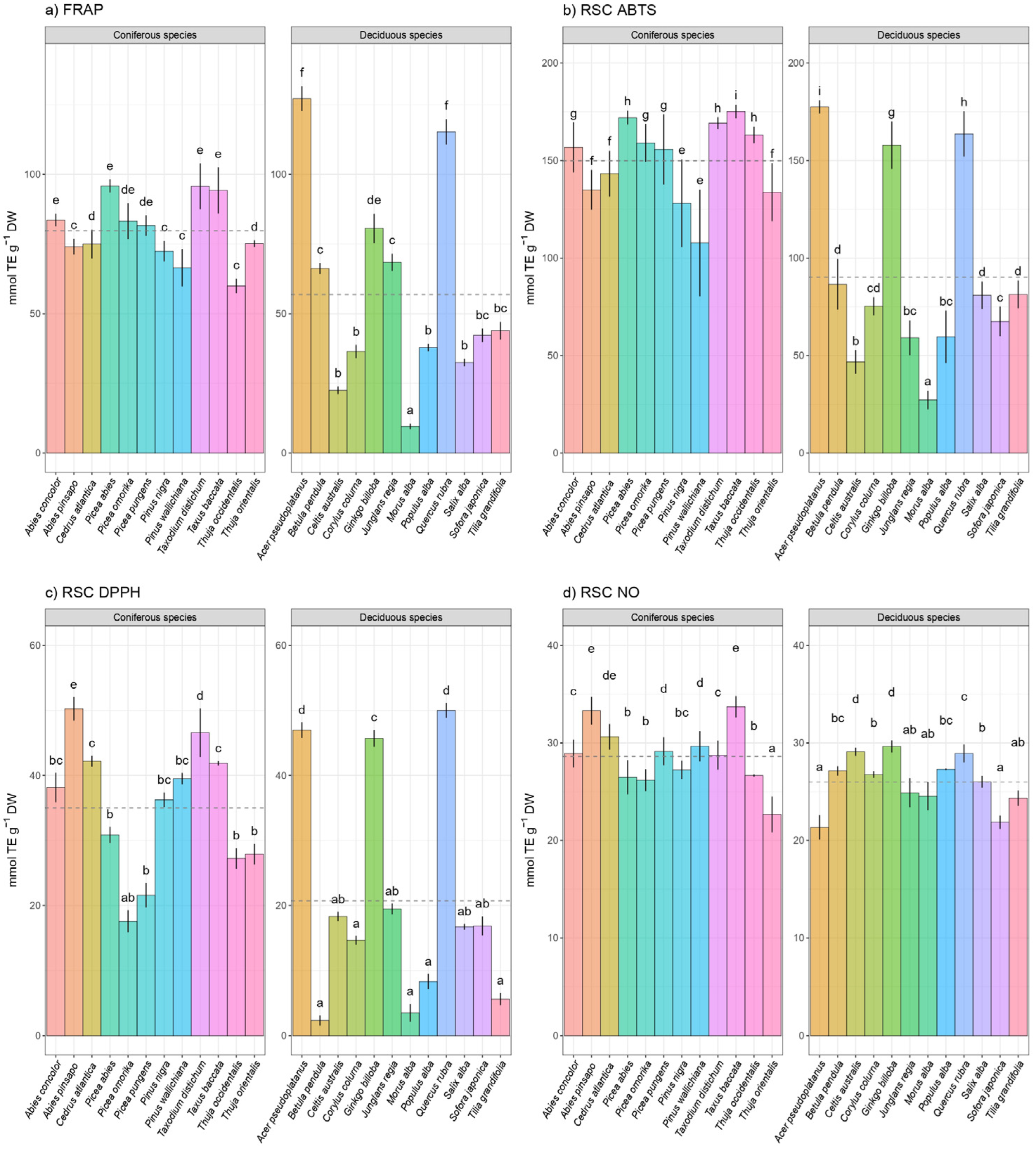
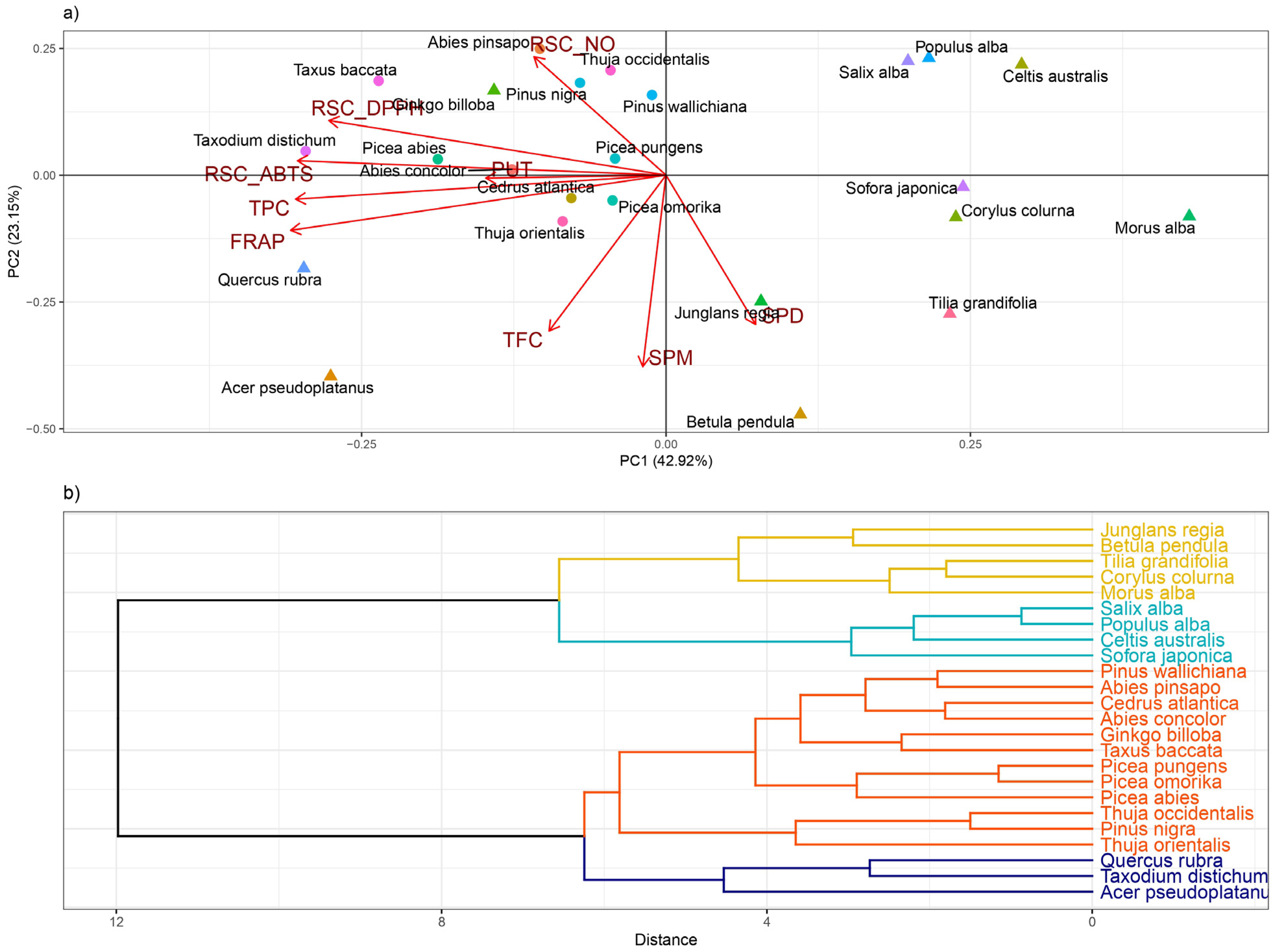
3. Results
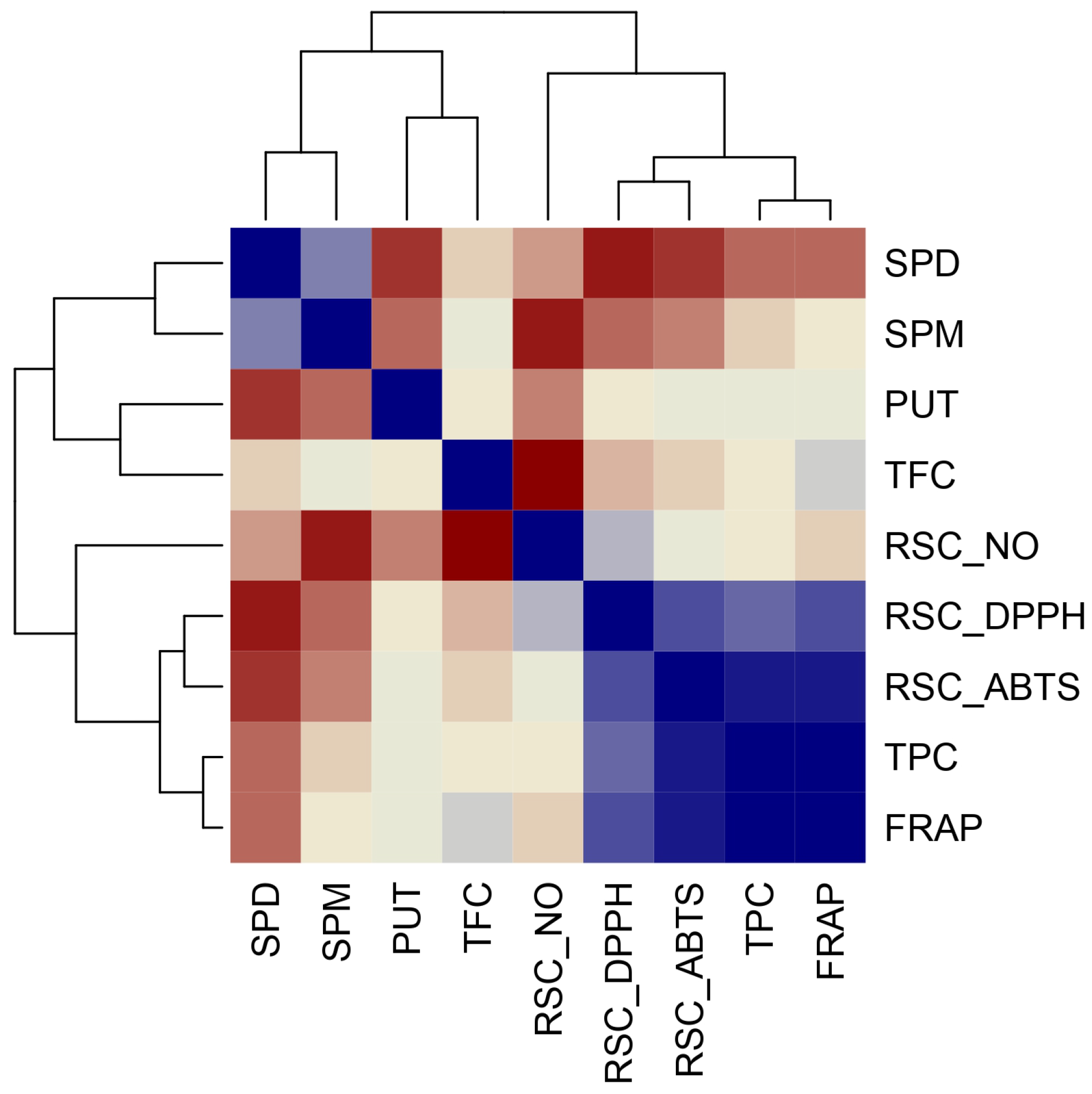
Hierarchical Claster Analysis, Principal Component Analysis (PCA), and Correlation Matrix
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Esperon-Rodriguez, M.; Rymer, P.D.; Power, S.A.; Challis, A.; Marchin, R.M.; Tjoelker, M.G. Functional adaptations and trait plasticity of urban trees along a climatic gradient. Urban For. Urban Green. 2020, 54, 126771. [Google Scholar] [CrossRef]
- Coumou, D.; Robinson, A. Historic and future increase in the global land area affected by monthly heat extremes. Environ. Res. Lett. 2013, 8, 034018. [Google Scholar] [CrossRef]
- Sharma, A.; Andhikaputra, G.; Wang, Y.C. Heatwaves in South Asia: Characterization, Consequences on Human Health, and Adaptation Strategies. Atmosphere 2022, 13, 734. [Google Scholar] [CrossRef]
- Ulpiani, G. On the linkage between urban heat island and urban pollution island: Three-decade literature review towards a conceptual framework. Sci. Total Environ. 2021, 751, 141727. [Google Scholar] [CrossRef] [PubMed]
- Pukowiec-Kurda, K. The urban ecosystem services index as a new indicator for sustainable urban planning and human well-being in cities. Ecol. Indic. 2022, 144, 109532. [Google Scholar] [CrossRef]
- Rahmonov, O.; Pukowiec-Kurda, K.; Banaszek, J.; Brom, K. Floristic diversity in selected city parks in southern Poland. Environ. Prot. Nat. Resour. 2020, 30, 8–17. [Google Scholar] [CrossRef]
- McPherson, E.G.; Berry, A.M.; van Doorn, N.S. Performance testing to identify climate-ready trees. Urban For. Urban Green. 2018, 29, 28–39. [Google Scholar] [CrossRef]
- Kesić, L.; Vuksanović, V.; Karaklić, V.; Vaštag, E. Variation of Leaf Water Potential and Leaf Gas Exchange Parameters of Seven Silver Linden (Tilia tomentosa Moench) Genotypes in Urban Environment. Topola 2020, 205, 15–24. [Google Scholar] [CrossRef]
- Kebert, M.; Rapparini, F.; Neri, L.; Bertazza, G.; Orlović, S.; Biondi, S. Copper-Induced Responses in Poplar Clones are Associated with Genotype-and Organ-Specific Changes in Peroxidase Activity and Proline, Polyamine, ABA, and IAA Levels. J. Plant Growth Regul. 2017, 36, 131–147. [Google Scholar] [CrossRef]
- De la Sota, C.; Ruffato-Ferreira, V.J.; Ruiz-García, L.; Alvarez, S. Urban green infrastructure as a strategy of climate change mitigation. A case study in northern Spain. Urban For. Urban Green. 2019, 40, 145–151. [Google Scholar] [CrossRef]
- Vastag, E.; Kesić, L.; Karaklić, V.; Zorić, M.; Vuksanović, V.; Stojnić, S. Physiological performance of sweetgum (Liquidambar stryraciflua L.) and norway maple (Acer platanoides L.) under drought condition in urban environment. Topola 2019, 204, 17–27. [Google Scholar]
- Esperon-Rodriguez, M.; Tjoelker, M.G.; Lenoir, J.; Baumgartner, J.B.; Beaumont, L.J.; Nipperess, D.A.; Power, S.A.; Richard, B.; Rymer, P.D.; Gallagher, R.V. Climate change increases global risk to urban forests. Nat. Clim. Chang. 2022, 12, 950–955. [Google Scholar] [CrossRef]
- Dale, A.G.; Frank, S.D. Warming and Drought Combine to Increase Pest Insect Fitness on Urban Trees. PLoS ONE 2017, 12, e0173844. [Google Scholar] [CrossRef]
- Calfapietra, C.; Peñuelas, J.; Niinemets, Ü. Urban plant physiology: Adaptation-mitigation strategies under permanent stress. Trends Plant Sci. 2015, 20, 72–75. [Google Scholar] [CrossRef]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant. Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef]
- Tanwir, K.; Amna; Javed, M.T.; Shahid, M.; Akram, M.S.; Ali, Q. Antioxidant Defense Systems in Bioremediation of Organic Pollutants. In Handbook of Bioremediation: Physiological; Mirza, H., Majeti, N.V.P., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 505–521. ISBN 9780128193822. [Google Scholar]
- Kebert, M.; Kostić, S.; Stojnić, S.; Čapelja, E.; Markić, A.G.; Zorić, M.; Kesić, L.; Flors, V. A Fine-Tuning of the Plant Hormones, Polyamines and Osmolytes by Ectomycorrhizal Fungi Enhances Drought Tolerance in Pedunculate Oak. Int. J. Mol. Sci. 2023, 24, 7510. [Google Scholar] [CrossRef]
- Kebert, M.; Kostić, S.; Čapelja, E.; Vuksanović, V.; Stojnić, S.; Markić, A.G.; Zlatković, M.; Milović, M.; Galović, V.; Orlović, S. Ectomycorrhizal Fungi Modulate Pedunculate Oak’s Heat Stress Responses through the Alternation of Polyamines, Phenolics, and Osmotica Content. Plants 2022, 11, 3360. [Google Scholar] [CrossRef]
- Kebert, M.; Vuksanović, V.; Stefels, J.; Bojović, M.; Horák, R.; Kostić, S.; Kovačević, B.; Orlović, S.; Neri, L.; Magli, M.; et al. Species-Level Differences in Osmoprotectants and Antioxidants Contribute to Stress Tolerance of Quercus robur L., and Q. cerris L. Seedlings under Water Deficit and High Temperatures. Plants 2022, 11, 1744. [Google Scholar] [CrossRef] [PubMed]
- Biondi, S.; Antognoni, F.; Marincich, L.; Lianza, M.; Tejos, R.; Ruiz, K.B. The polyamine “multiverse” and stress mitigation in crops: A case study with seed priming in quinoa. Sci. Hortic. 2022, 304, 111292. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Gondor, O.K.; Janda, T. Unfinished story of polyamines: Role of conjugation, transport and light-related regulation in the polyamine metabolism in plants. Plant Sci. 2021, 308, 110923. [Google Scholar] [CrossRef] [PubMed]
- Aktar, F.; Islam, M.S.; Milon, M.A.-A.; Islam, N.; Islam, M.A. Polyamines: An Essentially Regulatory Modulator of Plants to Abiotic Stress Tolerance: A Review. Asian J. Appl. Sci. 2021, 9, 195–204. [Google Scholar] [CrossRef]
- Sobieszczuk-Nowicka, E.; Paluch-Lubawa, E.; Mattoo, A.K.; Arasimowicz-Jelonek, M.; Gregersen, P.L.; Pacak, A. Polyamines—A new metabolic switch: Crosstalk with networks involving senescence, crop improvement, and mammalian cancer therapy. Front. Plant Sci. 2019, 10, 460298. [Google Scholar] [CrossRef] [PubMed]
- Kebert, M.; Kostić, S.; Zlatković, M.; Stojnic, S.; Čapelja, E.; Zorić, M.; Kiprovski, B.; Budakov, D.; Orlović, S. Ectomycorrhizal Fungi Modulate Biochemical Response against Powdery Mildew Disease in Quercus robur L. Forests 2022, 13, 1491. [Google Scholar] [CrossRef]
- Milošević, D. Application and Evaluation of Classification System of Local Climate Zones Using Automatic Model and Bioclimate Analysis. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2018. [Google Scholar]
- Bajsanski, I.; Stojakovic, V.; Jovanovic, M. Effect of tree location on mitigating parking lot insolation. Comput. Environ. Urban Syst. 2016, 56, 59–67. [Google Scholar] [CrossRef]
- Pekeč, S.; Marković, M.; Kebert, M.; Karaklić, V. Osobine zemljišta na području Futoškog parka u Novom Sadu. Šumarstvo 2020, 2020, 111–118. [Google Scholar]
- Kostić, S.; Kebert, M.; Todorović, H.; Pekeč, S.; Zorić, M.; Stojanović, D.; Orlović, S. Soil horizon-dependent heavy metals, and micro-and macro-elements distributions: A case study of Futoški park (Novi Sad, Serbia). Topola 2022, 9, 15–27. [Google Scholar] [CrossRef]
- Kostić, S.; Čukanović, J.; Orlović, S.; Ljubojević, M.; Mladenović, E. Allometric Relations of Sycamore Maple (Acer pseudoplatanus) and its Red Leaf Cultivar (A. pseudoplatanus “ Atropurpureum ”) in Street and Park Habitats of Novi Sad (Serbia, Europe). J. For. 2019, 117, 114–127. [Google Scholar] [CrossRef]
- Scaramagli, S.; Blondi, S.; Torrigiani, P. Methylglyoxal(bis-guanylhydrazone) inhibition of organogenesis is not due to S-adenosylmethionine decarboxylase inhibition/polyamine depletion in tobacco thin layers. Physiol. Plant. 1999, 107, 353–360. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Factors Influencing the Antioxidant Activity Determined by the ABTS•+ Radical Cation Assay. Free. Radic. Res. 2009, 26, 195–199. [Google Scholar] [CrossRef]
- Arnao, M.B. Some methodological problems in the determination of antioxidant activity using chromogen radicals: A practical case. Trends Food Sci. Technol. 2000, 11, 419–421. [Google Scholar] [CrossRef]
- Hensley, K.; Floyd, R.A.; Hensley, K.; Mou, S.; Pye, Q.N. Nitrite Determination by Colorimetric and Fluorometric Griess Diazotization Assays. In Methods in Biological Oxidative Stress; Hensley, K., Floyd, R.A., Eds.; Humana Press: Totowa, NJ, USA, 2003; pp. 185–193. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colometric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Tebartz, F.; Scherer, G.F. Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 2011, 181, 593–603. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef] [PubMed]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Free and cell wall-bound polyamines under long-term water stress applied at different growth stages of × Triticosecale Wittm. PLoS ONE 2015, 10, e0135002. [Google Scholar] [CrossRef]
- Del Duca, S.; Aloisi, I.; Parrotta, L.; Cai, G. Cytoskeleton, transglutaminase and gametophytic self-incompatibility in the Malinae (Rosaceae). Int. J. Mol. Sci. 2019, 20, 209. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Sengupta, A.; Chakraborty, M.; Gupta, B. Hydrogen peroxide and polyamines act as double edged swords in plant abiotic stress responses. Front. Plant Sci. 2016, 7, 1343. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Bhardwaj, R.; Gupta, B.D.; Dutt, P.; Gupta, R.K.; Kanwar, M.; Biondi, S. Enhancing effects of 24-epibrassinolide and putrescine on the antioxidant capacity and free radical scavenging activity of Raphanus sativus seedlings under Cu ion stress. Acta Physiol. Plant. 2011, 33, 1319–1333. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, E.I.S.; Scherer, G.F. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef]
- Sánchez-Vicente, I.; Lorenzo, O. Nitric oxide regulation of temperature acclimation: A molecular genetic perspective. J. Exp. Bot. 2021, 72, 5789–5794. [Google Scholar] [CrossRef]
- Parankusam, S.; Adimulam, S.S.; Bhatnagar-Mathur, P.; Sharma, K.K. Nitric oxide (NO) in plant heat stress tolerance: Current knowledge and perspectives. Front. Plant Sci. 2017, 8, 1582. [Google Scholar] [CrossRef] [PubMed]
- Tiburcio, A.F.; Altabella, T.; Bitrián, M.; Alcázar, R. The roles of polyamines during the lifespan of plants: From development to stress. Planta 2014, 240, 1–18. [Google Scholar] [CrossRef]
- Škrbić, B.; Đurišić-Mladenović, N.; Živančev, J.; Tadić, Đ. Seasonal occurrence and cancer risk assessment of polycyclic aromatic hydrocarbons in street dust from the Novi Sad city, Serbia. Sci. Total Environ. 2019, 647, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.P.; Pang, X.M.; Matsuda, N.; Kita, M.; Inoue, H.; Hao, Y.J.; Honda, C.; Moriguchi, T. Over-expression of the apple spermidine synthase gene in pear confers multiple abiotic stress tolerance by altering polyamine titers. Trans. Res. 2008, 17, 251–263. [Google Scholar] [CrossRef]
- Wen, X.P.; Ban, Y.; Inoue, H.; Matsuda, N.; Kita, M.; Moriguchi, T. Antisense inhibition of a spermidine synthase gene highlights the role of polyamines for stress alleviation in pear shoots subjected to salinity and cadmium. Environ. Exp. Bot. 2011, 72, 157–166. [Google Scholar] [CrossRef]
- Zou, Y.-N.; Zhang, F.; Srivastava, A.K.; Wu, Q.-S.; Kuča, K. Arbuscular Mycorrhizal Fungi Regulate Polyamine Homeostasis in Roots of Trifoliate Orange for Improved Adaptation to Soil Moisture Deficit Stress. Front. Plant Sci. 2021, 11, 600792. [Google Scholar] [CrossRef]
- Salo, H.M.; Sarjala, T.; Jokela, A.; Häggman, H.; Vuosku, J. Moderate stress responses and specific changes in polyamine metabolism characterize Scots pine somatic embryogenesis. Tree Physiol. 2016, 36, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Newton, R.J. Polyamines promote root elongation and growth by increasing root cell division in regenerated Virginia pine (Pinus virginiana Mill.) plantlets. Plant Cell Rep. 2005, 24, 581–589. [Google Scholar] [CrossRef]
- Sharma, K.; Gupta, S.; Thokchom, S.D.; Jangir, P.; Kapoor, R. Arbuscular Mycorrhiza-Mediated Regulation of Polyamines and Aquaporins During Abiotic Stress: Deep Insights on the Recondite Players. Front. Plant Sci. 2021, 12, 642101. [Google Scholar] [CrossRef] [PubMed]
- Tailor, A.; Bhatla, S.C. Polyamine homeostasis modulates plasma membrane-and tonoplast-associated aquaporin expression in etiolated salt-stressed sunflower (Helianthus annuus L.) seedlings. Protoplasma 2021, 258, 661–672. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines are important in abiotic stress signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef]
- Huang, D.; Boxin, O.U.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Urban Area | Natural Area | ||
|---|---|---|---|---|
| Temperature (°C) | Relative Humidity (%) | Temperature (°C) | Relative Humidity (%) | |
| January | 1.3 | 85.1 | −0.2 | 91.6 |
| February | 7.5 | 79.5 | 5.7 | 88.0 |
| March | 10.5 | 69.8 | 8.9 | 78.3 |
| April | 14.2 | 58.8 | 12.6 | 69.2 |
| May | 18.8 | 68.4 | 17.2 | 78.0 |
| June | 23.4 | 64.4 | 21.6 | 77.5 |
| July | 25.0 | 61.6 | 23.0 | 76.2 |
| August | 23.7 | 67.4 | 21.6 | 81.3 |
| September | 19.5 | 74.0 | 17.8 | 86.8 |
| October | 12.5 | 82.6 | 11.3 | 90.4 |
| November | 9.2 | 80.0 | 7.4 | 88.4 |
| December | 3.5 | 87.0 | 2.0 | 93.4 |
| Average | 14.09 | 73.22 | 12.41 | 83.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kebert, M.; Kostić, S.; Rašeta, M.; Stojanović, D.V.; Stojnić, S.; Orlović, S. Species-Specific Level Variation in Polyamines in Coniferous and Deciduous Woody Plant Species in Urban Areas. Horticulturae 2023, 9, 1157. https://doi.org/10.3390/horticulturae9101157
Kebert M, Kostić S, Rašeta M, Stojanović DV, Stojnić S, Orlović S. Species-Specific Level Variation in Polyamines in Coniferous and Deciduous Woody Plant Species in Urban Areas. Horticulturae. 2023; 9(10):1157. https://doi.org/10.3390/horticulturae9101157
Chicago/Turabian StyleKebert, Marko, Saša Kostić, Milena Rašeta, Dejan V. Stojanović, Srđan Stojnić, and Saša Orlović. 2023. "Species-Specific Level Variation in Polyamines in Coniferous and Deciduous Woody Plant Species in Urban Areas" Horticulturae 9, no. 10: 1157. https://doi.org/10.3390/horticulturae9101157