

Trewia nudiflora Linn, a Medicinal Plant: Allelopathic Potential and Characterization of Bioactive Compounds from Its Leaf Extracts

Abstract
:1. Introduction
2. Materials and Methods
2.1. Trewia nudiflora Samples and Test Plant Species
2.2. Extraction of Trewia nudiflora Leaves for the Bioassay Experiment
2.3. Method for Isolating and Purifying Active Allelopathic Substances
2.4. Bioactivity of the Identified Substances
2.5. Analysis
3. Results
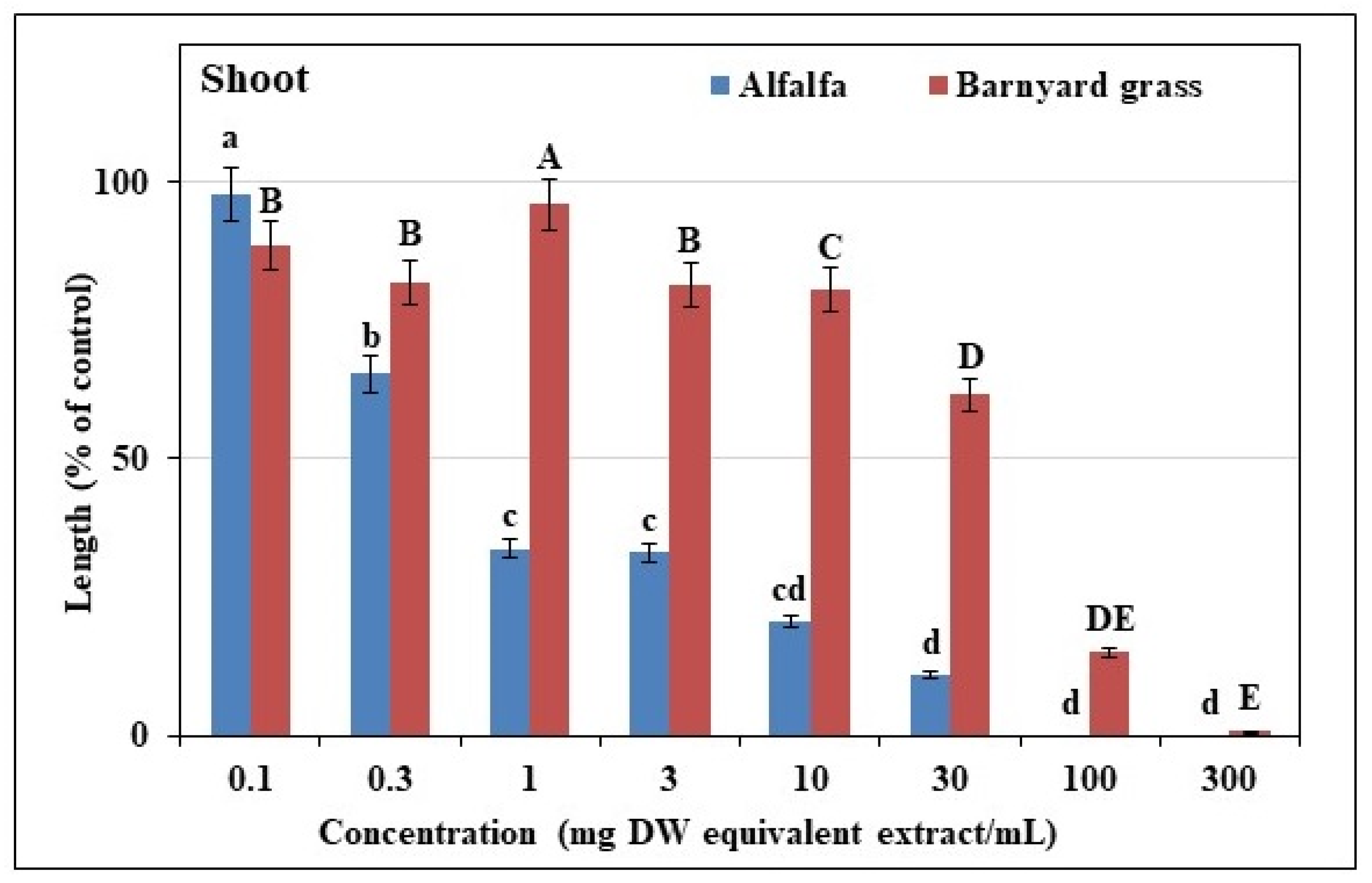
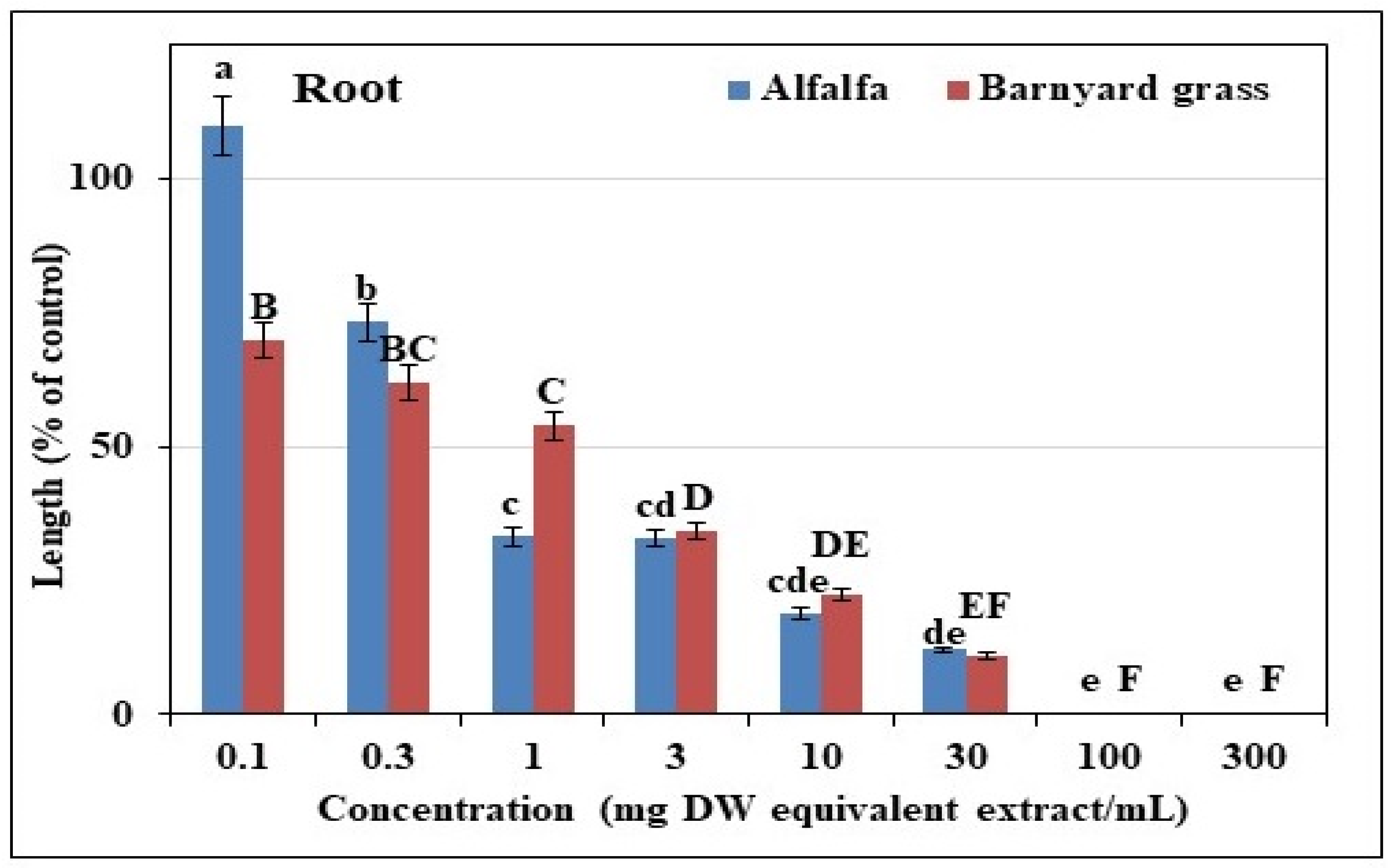
3.1. Effects of Extracts from Trewia nudiflora Leaves on Test Plants
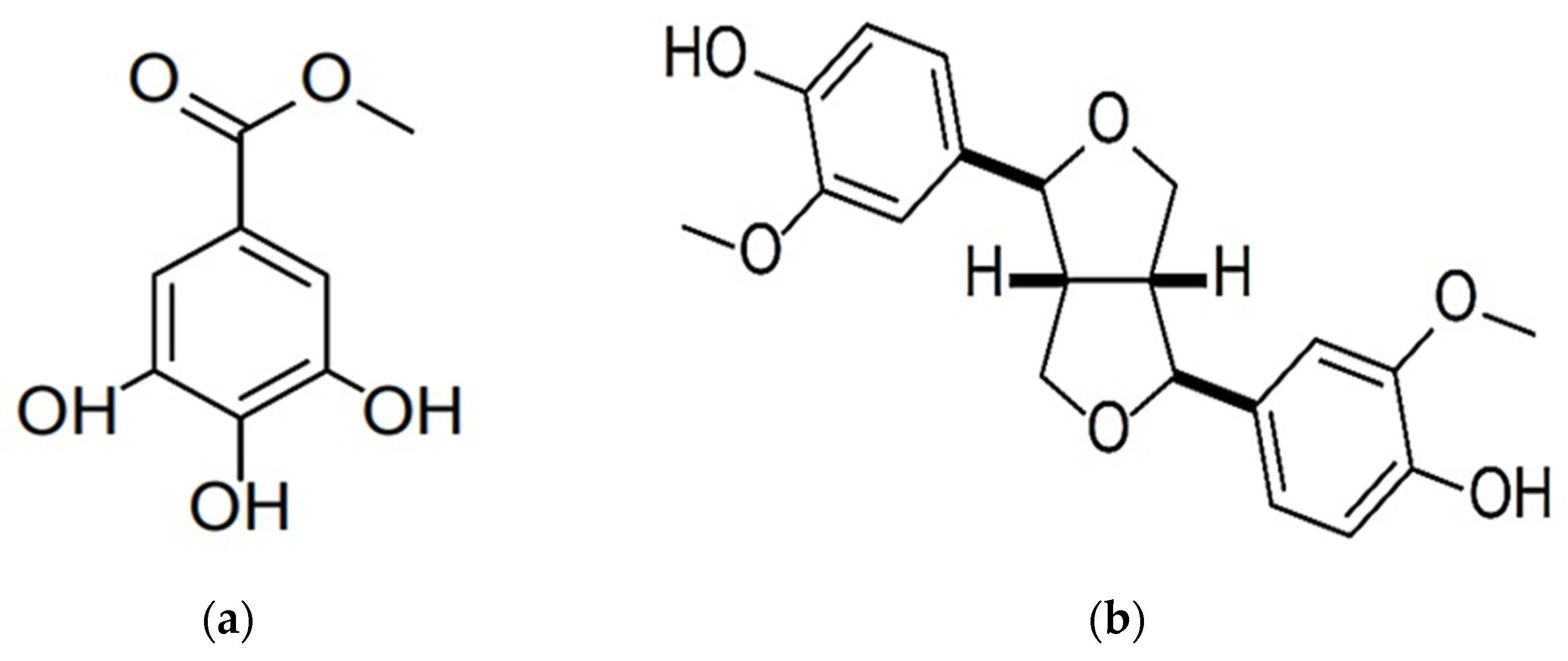
3.2. Characterization of the Isolated Compounds
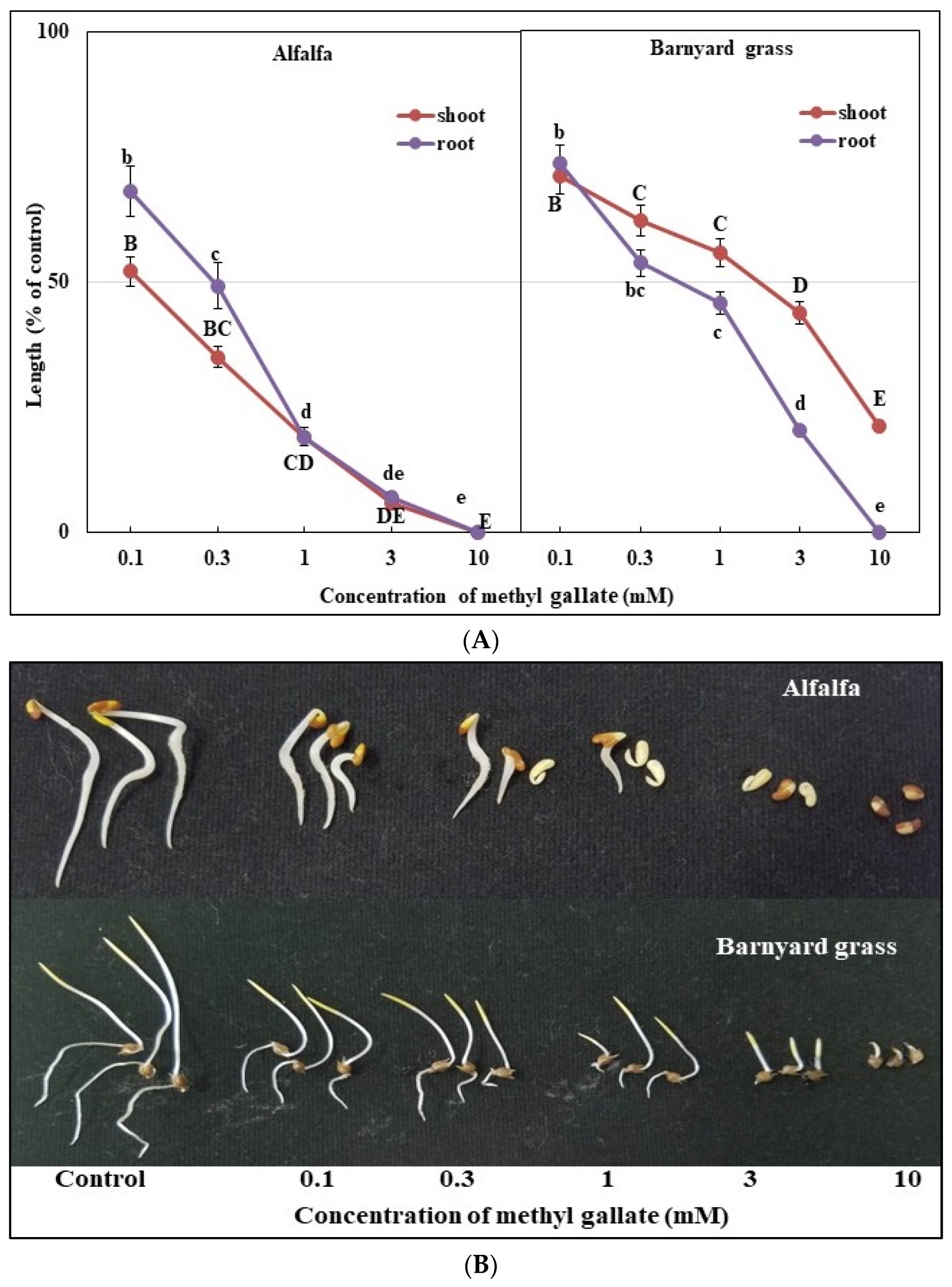
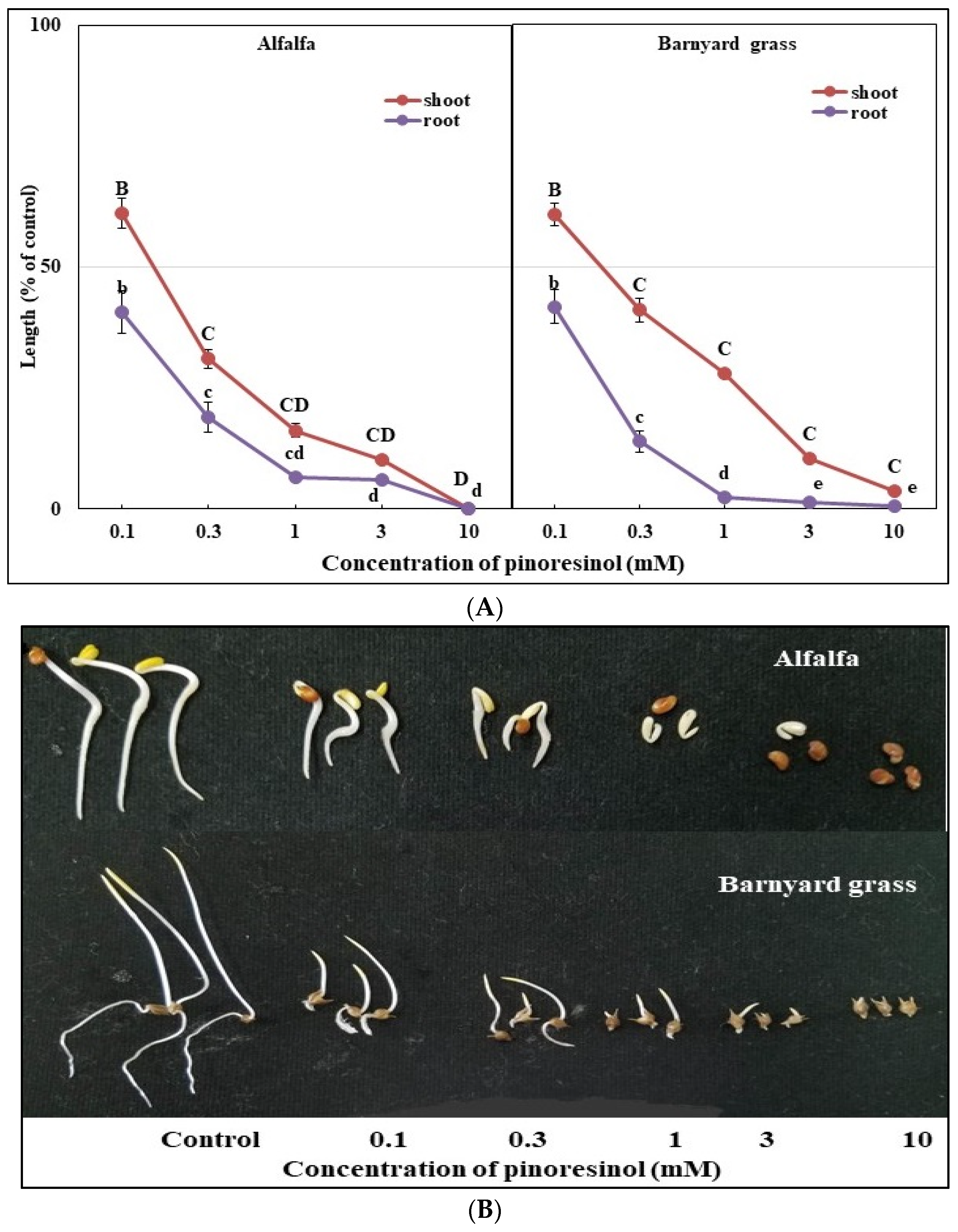
3.3. Bioactivities of the Two Substances (Methyl Gallate and Pinoresinol)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ram, R.; Mehrotra, B.N.; Sinha, S.; Pant, P.; Sheth, R. Compendium of Indian Medicinal Plants; CDRI Lucknow and National Institute of Science Communication: New Delhi, India, 2004. [Google Scholar]
- Kulju, K.K.M.; Sierra, S.E.C.; Van Welzen, P.C. Re-shaping Mallotus [part 2]: Inclusion of Neotrewia, Octospermum and Trewia in Mallotus ss (Euphorbiaceae ss). Blumea-Biodiversity. Evol. Biogeogr. 2007, 52, 115–136. [Google Scholar] [CrossRef] [Green Version]
- Trewia nudiflora. Flora of China. Available online: https://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=200013883 (accessed on 2 June 2021).
- Sultana, R.; Milon, M.M.M.; Kader, M.A.; Parvin, S.; Masud, G.M. Trewia nudiflora: A potential source of new drugs. J. Phytopharm. 2022, 11, 421–424. [Google Scholar] [CrossRef]
- Nasrine, S.; El-taher, S.E.D.H. Allelopathic effect of Euphorbia guyoniana aqueous extract and their potential uses as natural herbicides. Sains Malays. 2013, 42, 1501–1504. [Google Scholar]
- Da Silva, U.P.; Furlani, G.M.; Demuner, A.J.; da Silva, O.L.M.; Varejão, E.V.V. Allelopathic activity and chemical constituents of extracts from roots of Euphorbia heterophylla L. Nat. Prod. Res. 2018, 33, 2681–26844. [Google Scholar] [CrossRef]
- Ripa, F.A.; Hossain, M.J.; Munira, M.S.; Roy, A.; Riya, F.H.; Alam, F.; Khidir, E.B. Phytochemical and pharmacological profiling of Trewia nudiflora Linn. leaf extract deciphers therapeutic potentials against thrombosis, arthritis, helminths, and insects. Open Chem. 2022, 20, 1304–1312. [Google Scholar] [CrossRef]
- Powell, R.G.; Weisleder, D.; Smith, C.R., Jr. Novel maytansinoid tumor inhibitors from Trewia nudiflora: Trewiasine, de-hydrotrewiasine, and demethyltrewiasine. J. Org. Chem. 1981, 46, 4398–4403. [Google Scholar] [CrossRef]
- Kumar, K.P.; Sastry, V.G. Protective of Trewia nudiflora against ischemic stroke in experimental rats. Inter. J. Pharmacother. 2012, 2, 7–12. [Google Scholar]
- Balakrishnan, N.; Srivastava, M.; Tiwari, P. Preliminary phytochemical analysis and DPPH free radical scavenging activity of Trewia nudiflora Linn. roots and leaves. Pak. J. Biol. Sci. 2013, 16, 1403–1406. [Google Scholar] [CrossRef] [Green Version]
- Esan, V.; Elanchezhiyan, C.; Mahboob, S.; Al-Ghanim, K.A.; Al-Misned, F.; Ahmed, Z.; Marimuthu, G. Toxicity of Trewia nudiflora-mediated silver nanoparticles on mosquito larvae and non-target aquatic fauna. Toxin Rev. 2022, 41, 229–236. [Google Scholar] [CrossRef]
- Begum, Y. Antibacterial, antioxidant and cytotoxic activities of Trewia nudiflora. Pharma Tutor. 2016, 4, 37–41. Available online: http://www.pharmatutorjournal.com/index.php/pt/article/view/332 (accessed on 1 January 2016).
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984. [Google Scholar]
- Kruse, M.; Strandberg, M.; Strandberg, B. Ecological Effects of Allelopathic Plants—A Review; NERI Technical Report; National Environmental Research Institute: Roskilde, Denmark, 2000; p. 315. [Google Scholar]
- Bertin, C.; Yang, X.H.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Weir, T.L.; Park, S.W.; Vivanco, J.M. Biochemical and physiological mechanisms mediated by allelochemicals. Curr. Opin. Plant Biol. 2004, 7, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; Van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Yuan, L.; Li, J.; Van Kleunen, M. Competition induces negative conspecific allelopathic effects on seedling recruitment. Ann. Bot. 2022, 130, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Bhadoria, P.B.S. Allelopathy: A natural way towards weed management. Am. J. Exp. Agric. 2011, 1, 7–20. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowska, A. Allelochemicals as Bioherbicides—Present and Perspectives. In Herbicides—Current Research and Case Studies in Use; IntechOpen: London, UK, 2013; Volume Chapter 20, pp. 518–542. [Google Scholar] [CrossRef] [Green Version]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Zhang, J.; An, M.; Wu, H.; Liu, D.L.; Stanton, R. Phytotoxic activity and chemical composition of aqueous volatile fractions from Eucalyptus species. PLoS ONE 2014, 9, e93189. [Google Scholar] [CrossRef] [Green Version]
- Al Harun, M.A.Y.; Johnson, J.; Uddin, M.N.; Robinson, R.W. Identification and phytotoxicity assessment of phenolic compounds in Chrysanthemoides monilifera subsp. monilifera (Boneseed). PLoS ONE 2015, 10, e0139992. [Google Scholar] [CrossRef] [Green Version]
- Devasinghe, D.A.U.D.; Premarathne, K.P.; Sangakkara, U.R. Weed management by rice straw mulching in direct seeded lowland rice (Oryza sativa L.). Trop. Agric. Res. 2011, 22, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Khatun, R.; Kato-Noguchi, H. Piper longum L. leaf extracts, a candidate allelopathic plant that suppressed the growth of six test plants, could be a source of potent phytotoxic compounds. Res. Crop. 2022, 23, 874–880. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, version 16.0; IBM Corp: Armonk, NY, USA, 2007. [Google Scholar]
- Kiss, A.K.; Derwińska, M.; Dawidowska, A.; Naruszewicz, M. Novel biological properties of Oenothera paradoxa defatted seed extracts: Effects on metallopeptidase activity. J. Agric. Food Chem. 2008, 56, 7845–7852. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, S.; Kishimoto, T.; Uraki, Y.; Okamoto, T.; Ubukata, M. Low molecular weight lignin suppresses activation of NF-κB and HIV-1 promoter. Bioorg. Med. Chem. 2008, 16, 2645–2650. [Google Scholar] [CrossRef]
- Islam, A.M.; Kato-Noguchi, H. Allelopathic prospective of Ricinus communis and Jatropha curcas for bio-control of weeds. Acta Agric. Scand. B Soil Plant Sci. 2013, 63, 731–739. [Google Scholar] [CrossRef]
- Das, K.R.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Evaluation of phytotoxic potential and identification of phytotoxic substances in Cassia alata Linn. leaves. Acta Agric. Scand. B Soil Plant Sci. 2019, 69, 479–488. [Google Scholar] [CrossRef]
- Mousavi, S.S.; Karami, A.; Haghighi, T.M.; Alizadeh, S.; Maggi, F. Phytotoxic potential and phenolic profile of extracts from Scrophularia striata. Plants 2021, 10, 135. [Google Scholar] [CrossRef]
- Krumsri, R.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Assessment of allelopathic potential of Senna garrettiana leaves and identification of potent phytotoxic substances. Agronomy 2022, 12, 139. [Google Scholar] [CrossRef]
- Khatun, M.R.; Tojo, S.; Teruya, T.; Kato-Noguchi, H. The allelopathic effects of Trewia nudiflora leaf extracts and its identified substances. Plants 2023, 12, 1375. [Google Scholar] [CrossRef]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.; Sarada, S. Role of phenolics in allelopathic interactions. Allelopath. J. 2012, 29, 215–230. [Google Scholar]
- Fatholahi, S.; Karimmojeni, H.; Ehsanzadeh, P. Phenolic compounds and allelopathic activities of ancient emmer wheats: Perspective for non-chemical weed control scenarios. Acta Physiol. Plant. 2020, 42, 135. [Google Scholar] [CrossRef]
- Favaretto, A.; Schefer-Basso, S.M.; Perez, N.B. Allelopathy in Poaceae species present in Brazil. A review. Agron. Sustain. Dev. 2018, 38, 22. [Google Scholar] [CrossRef] [Green Version]
- Rob, M.M.; Hossen, K.; Khatun, M.R.; Iwasaki, K.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Identification and application of bioactive compounds from Garcinia xanthochymus Hook. for weed management. Appl. Sci. 2021, 11, 2264. [Google Scholar] [CrossRef]
- Šćepanović, M.; Košćak, L.; Šoštarčić, V.; Pismarović, L.; Milanović-Litre, A.; Kljak, K. Selected phenolic acids inhibit the initial growth of Ambrosia artemisiifolia L. Biology 2022, 11, 482. [Google Scholar] [CrossRef]
- Calzada, F.; Cerda-García-Rojas, C.M.; Meckes, M.; Cedillo-Rivera, R.; Bye, R.; Mata, R. Geranins A and B, new antiprotozoal A-type proanthocyanidins from Geranium niveum. J. Nat. Prod. 1999, 62, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Park, D.J.; Jung, H.J.; Park, C.H.; Yokozawa, T.; Jeong, J.C. Root bark of Paeonia suffruticosa extract and its component methyl gallate possess peroxynitrite scavenging activity and anti-inflammatory properties through NF-κB inhibition in LPS-treated mice. Molecules 2019, 24, 3483. [Google Scholar] [CrossRef] [Green Version]
- Abou-Zaid, M.M.; Lombardo, D.A.; Nozzolillo, C. Methyl gallate is a natural constituent of maple (genus Acer) leaves. Nat. Prod. Res. 2009, 23, 1373–1377. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Heredia, N.; del Camacho-Corona, M.R.; García, S. Isolation, characterization and mode of antimicrobial action against Vibrio cholera of methyl gallate isolated from Acacia farnesiana. J. Appl. Microbiol. 2013, 115, 1307–1316. [Google Scholar] [CrossRef]
- Kang, M.S.; Oh, J.S.; Kang, I.C.; Hong, S.J.; Choi, C.H. Inhibitory effect of methyl gallate and gallic acid on oral bacteria. J. Microbiol. 2008, 46, 744–750. [Google Scholar] [CrossRef]
- Suzuki, M.; Khan, M.S.I.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathic potential and an allelopathic substance in mango leaves. Acta. Agric. Scand. B Soil Plant Sci. 2017, 67, 37–42. [Google Scholar] [CrossRef]
- Boonmee, S.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Evaluation of phytotoxic activity of leaf and stem extracts and identification of a phytotoxic substance from Caesalpinia mimosoides Lamk. Theor. Exp. Plant Physiol. 2018, 30, 129–139. [Google Scholar] [CrossRef]
- Yue, F.; Lan, W.; Zhang, L.; Lu, F.; Sun, R.; Ralph, J. Efficient synthesis of pinoresinol, an important lignin dimeric model compound. Front. Energy Res. 2021, 9, 640337. [Google Scholar] [CrossRef]
- Jung, H.W.; Mahesh, R.; Lee, J.G.; Lee, S.H.; Kim, Y.S.; Park, Y.K. Pinoresinol from the fruits of Forsythia koreana inhibits inflammatory responses in LPS-activated microglia. Neurosci. Lett. 2010, 480, 215–220. [Google Scholar] [CrossRef]
- Milder, I.E.; Arts, I.C.; Van de Putte, B.; Venema, D.P.; Hollman, P.C. Lignan contents of Dutch plant foods: A database including lariciresinol, pinoresinol, secoisolariciresinol and matairesinol. Br. J. Nutr. 2005, 93, 393–402. [Google Scholar] [CrossRef]
- Owen, R.W.; Giacosa, A.; Hull, W.E.; Haubner, R.; Spiegelhalder, B.; Bartsch, H. The antioxidant/anticancer potential of phenolic compounds isolated from olive oil. Eur. J. Cancer 2000, 36, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, F.C.; Del Campo, M.L.; Grant, J.B.; Weibel, D.B.; Smedley, S.R.; Bolton, K.L.; Meinwald, J.; Eisner, T. Pinoresinol: A lignol of plant origin serving for defense in a caterpillar. Proc. Natl. Acad. Sci. USA 2006, 103, 15497–15501. [Google Scholar] [CrossRef] [PubMed]
- During, A.; Debouche, C.; Raas, T.; Larondelle, Y. Among plant lignans, pinoresinol has the strongest anti-inflammatory properties in human intestinal Caco-2 cells. J. Nutr. 2012, 142, 1798–1805. [Google Scholar] [CrossRef] [Green Version]
- Fini, L.; Hotchkiss, E.; Fogliano, V.; Graziani, G.; Romano, M.; De Vol, E.B.; Ricciardiello, L. Chemopreventive properties of pinoresinol-rich olive oil involves a selective activation of the ATM–p53 cascade in colon cancer cell lines. Carcinogenesis 2008, 29, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Tebboub, O.; Cotugno, R.; Oke-Altuntas, F.; Bouheroum, M.; Demirtas, Í.; D’Ambola, M.; Vassallo, A. Antioxidant potential of herbal preparations and components from Galactites elegans (All.) Nyman ex Soldano. Evid. Based Complement. Altern. Med. 2018, 2018, 9294358. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.; Lee, J.; Liu, Q.H.; Woo, E.R.; Lee, D.G. Antifungal effect of (+)-pinoresinol isolated from Sambucus williamsii. Molecules 2010, 15, 3507–3516. [Google Scholar] [CrossRef] [Green Version]
- Scavo, A.; Rial, C.; Molinillo, J.M.; Varela, R.M.; Mauromicale, G.; Macias, F.A. The extraction procedure improves the allelopathic activity of cardoon (Cynara cardunculus var. altilis) leaf allelochemicals. Ind. Crops Prod. 2019, 128, 479–487. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Hamada, Y.; Kojima, M.; Kumagai, S.; Iwasaki, A.; Suenaga, K. Allelopathic substances of Osmanthus spp. for developing sustainable agriculture. Plants 2023, 12, 376. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.R.; Tojo, S.; Teruya, T.; Kato-Noguchi, H. Allelopathic activity of Annona reticulata L. leaf extracts and identification of three allelopathic compounds for the development of natural herbicides. Agronomy 2022, 12, 2883. [Google Scholar] [CrossRef]
- Oliva, A.; Moraes, R.M.; Watson, S.B.; Duke, S.O.; Dayan, F.E. Aryltetralin lignans inhibit plant growth by affecting the formation of mitotic microtubular organizing centers. Pestic. Biochem. Physiol. 2002, 72, 45–54. [Google Scholar] [CrossRef]
- Zhou, H.; Ren, J.; Li, Z. Antibacterial activity and mechanism of pinoresinol from Cinnamomum camphora leaves against food-related bacteria. Food Control 2017, 79, 192–199. [Google Scholar] [CrossRef]
- Wu, L.; Guo, X.; Harivandi, M.A. Allelopathic effects of phenolic acids detected in buffalograss (Buchloe dactyloides) clippings on growth of annual bluegrass (Poa annua) and buffalograss seedlings. Environ. Exp. Bot. 1998, 39, 159–167. [Google Scholar] [CrossRef]
- Nishida, N.; Tamotsu, S.; Nagata, N.; Saito, C.; Sakai, A. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: Inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. J. Chem. Ecol. 2005, 31, 1187–1203. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Z.; Lei, X.U.; Jin-jing, S.; Zhang, Z.H.; Fu, M.L.; Teng, H.Y.; Yi, K.K. Allelochemical p-hydroxybenzoic acid inhibits root growth via regulating ROS accumulation in cucumber (Cucumis sativus L.). J. Integr. Agric. 2020, 19, 518–527. [Google Scholar] [CrossRef]
- Kaur, H.; Kaushik, S. Cellular evidence of allelopathic interference of benzoic acid to mustard (Brassica juncea L.) seedling growth. Plant Physiol. Biochem. 2005, 43, 77–81. [Google Scholar] [CrossRef]
- Dos Santos, W.D.; Ferrarese, M.D.L.L.; Finger, A.; Teixeira, A.C.; Ferrarese-Filho, O. Lignification and related enzymes in Glycine max root growth-inhibition by ferulic acid. J. Chem. Ecol. 2004, 30, 1203–1212. [Google Scholar] [CrossRef]
- Reigosa, M.J.; Souto, X.C.; González, L. Effect of phenolic compounds on the germination of six weeds species. Plant Growth Regul. 1999, 28, 83–88. [Google Scholar] [CrossRef]
- Piyatida, P.; Suenaga, K.; Kato-Noguchi, H. Allelopathic potential and chemical composition of Rhinacanthus nasutus extracts. Allelopath. J. 2010, 26, 207–216. [Google Scholar]
- Suzuki, T.; Usui, I.; Tomita-Yokotani, K.; Kono, S.; Tsubura, H.; Miki, Y.; Hasegawa, K. Effects of acid extracts of tomato (Lycopersicon esculentum Mill.) and carrot (Daucus carota L.) wastes from the food industry on the growth of some crops and weeds. Weed Biol. Manag. 2001, 1, 226–230. [Google Scholar] [CrossRef]
- Dayan, F.E.; Romagni, J.G.; Duke, S.O. Investigating the mode of action of natural phytotoxins. J. Chem. Ecol. 2000, 26, 2079–2094. [Google Scholar] [CrossRef]
- Kobayashi, K. Factors affecting phytotoxic activity of allelochemicals in soil. Weed Biol. Manag. 2004, 4, 1–7. [Google Scholar] [CrossRef]
- Maffei, M.; Bertea, C.M.; Garneri, F.; Scannerini, S. Effect of benzoic acid hydroxy-and methoxy-ring substituents during cucumber (Cucumis sativus L.) germination. I. Isocitrate lyase and catalase activity. Plant Sci. 1999, 141, 139–147. [Google Scholar] [CrossRef]
- Sanchez-Maldonado, A.F.; Schieber, A.; Ganzle, M.G. Structure-function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef]
- Synowiec, A.; Mozdzen, K.; Krajewska, A.; Landi, M.; Araniti, F. Carum carvi L. essential oil: A promising candidate for botanical herbicide against Echinochloa crus-galli (L.) P. Beauv. in maize cultivation. Ind. Crops Prod. 2019, 140, 111652. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic Effects and Mechanism of Action of Essential Oils and Terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef]
- Shiffler, A. Natural Weed Killers: A Guide to Organic Herbicides. Available online: https://www.lawnstarter.com/blog/lawn-care-2/natural-weed-killers-a-guide-to-organic-herbicides/ (accessed on 30 December 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Plant Species | I50 Value (mg DW Equivalent of T. nudiflora Extracts, mL−1) | ||
|---|---|---|---|
| Shoot | Root | ||
| Dicot | Alfalfa | 0.53 c | 0.60 c |
| Monocot | Barnyard Grass | 40.48 a | 1.24 b |
| Test Plant | I50 Value (mM) | ||
|---|---|---|---|
| Alfalfa | Methyl Gallate | Pinoresinol | |
| Shoot | 0.137 f | 0.149 e | |
| Root | 0.293 c | 0.065 h | |
| Barnyard Grass | Shoot | 2.051 a | 0.216 d |
| Root | 0.610 b | 0.080 g | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatun, M.R.; Tojo, S.; Teruya, T.; Kato-Noguchi, H. Trewia nudiflora Linn, a Medicinal Plant: Allelopathic Potential and Characterization of Bioactive Compounds from Its Leaf Extracts. Horticulturae 2023, 9, 897. https://doi.org/10.3390/horticulturae9080897
Khatun MR, Tojo S, Teruya T, Kato-Noguchi H. Trewia nudiflora Linn, a Medicinal Plant: Allelopathic Potential and Characterization of Bioactive Compounds from Its Leaf Extracts. Horticulturae. 2023; 9(8):897. https://doi.org/10.3390/horticulturae9080897
Chicago/Turabian StyleKhatun, Mst. Rokeya, Shunya Tojo, Toshiaki Teruya, and Hisashi Kato-Noguchi. 2023. "Trewia nudiflora Linn, a Medicinal Plant: Allelopathic Potential and Characterization of Bioactive Compounds from Its Leaf Extracts" Horticulturae 9, no. 8: 897. https://doi.org/10.3390/horticulturae9080897