
Mechanisms Underlying the C3–CAM Photosynthetic Shift in Facultative CAM Plants
Guangxi Key Laboratory of Plant Functional Phytochemicals and Sustainable Utilization, Guangxi Institute of Botany, Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin 541006, China
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(3), 398; https://doi.org/10.3390/horticulturae9030398
Submission received: 6 February 2023
/
Revised: 11 March 2023
/
Accepted: 15 March 2023
/
Published: 19 March 2023
(This article belongs to the Special Issue Physiological and Molecular Biology Research on Ornamental Flower)
Abstract
:Crassulacean acid metabolism (CAM), one of three kinds of photosynthesis, is a water-use efficient adaptation to an arid environment. CAM is characterized by CO2 uptake via open stomata during the nighttime and refixation CO2 via the Calvin cycle during the daytime. Facultative CAM plants can shift the photosynthesis from C3 to CAM and exhibit greater plasticity in CAM expression under different environments. Though leaf thickness is an important anatomical feature of CAM plants, there may be no anatomical feature changes during the C3–CAM transition for all facultative CAM plants. The shift from C3 photosynthesis to CAM in facultative CAM plants is accompanied by significant changes in physiology including stomata opening, CO2 gas exchange and organic acid fluxes; the activities of many decarboxylating enzymes increase during the shift from C3 to CAM; the molecular changes occur during the photosynthesis C3–CAM shift involved DNA hypermethylation, transcriptional regulation, post-transcriptional regulation and protein level regulation. Recently, omics approaches were used to discover more proceedings underling the C3–CAM transition. However, there are few reviews on the mechanisms involved in this photosynthetic shift in facultative CAM plants. In this paper, we summarize the progress in the comparative analysis of anatomical, physiological, metabolic and molecular properties of facultative CAM plants between C3 and CAM photosynthesis. Facultative CAM plants also show the potential for sustainable food crop and biomass production. We also discuss the implications of the photosynthesis transition from C3 to CAM on horticultural crops and address future directions for research.
1. Introduction
In the plant kingdom, there are three kinds of photosynthetic pathways: C3, C4 and crassulacean acid metabolism (CAM). CAM is characterized by CO2 uptake during the nighttime via open stomata, when CO2 is combined with phosphoenolpyruvate (PEP) and stored as organic acids (mainly malic acid). Then, organic acids are decarboxylated in the vacuoles during daytime and CO2 is refixed via the Calvin cycle [1,2]. Some plants can switch their photosynthesis between C3 and CAM, which are referred to as facultative (inducible or C3/CAM intermediate) CAM plants. The first discovered facultative CAM plant was Mesembryanthemum crystallinum, the common iceplant [3]. Other well-known facultative CAM plants include Sedum album [4], Clusia minor [5], Talinum triangulare [6], etc. These species are sampled as models to study other facultative CAM plants, as they can shift their photosynthetic mode in response to water deficit and other environmental stressors [7,8]. Furthermore, recent evidence suggests that facultative CAM plants may be more widespread among vascular plants than previously thought [9].
As we all know, photosynthesis is one of the most important chemical reactions on the earth [10]. Crassulacean acid metabolism (CAM), with higher photosynthetic, water-use and possibly nutrient-use efficiency, represents higher carbon-concentrating mechanisms (CCMs) than C3 photosynthesis in response to a warmer and drier world [11]. The phenomena of CAM is a typical ecophysiological adaptation to arid conditions [12,13,14]. Expression of CAM modes includes obligate CAM, facultative CAM, CAM-idling and CAM-cycling. Facultative CAM mode is one of the four plastic expressions of CAM modes. Facultative CAM plants can struggle with variable environments through the facultative CAM mode [14]. Recently, engineering CAM-related genes to C3 crops to improve water-use efficiency (WUE) has caused extensive attention [15,16,17]. In order to engineer CAM into C3 crops, a deep understanding of CAM-related genes and metabolic pathways is urgently needed [18].
However, the shift mechanism underlying C3 photosynthesis to CAM in facultative CAM plants is complicated, concerning the genetic changes required for the progression and reversion of this shift [19]. In order to engineer CAM into C3 crops to increase the water-use efficiency (WUE), a few facultative CAM species (such as M. crystallinum) were regarded as key tools to identify the genes involved in the CAM pathway and their respective regulation mechanisms [11,15,20,21]. In this paper, we review the signaling stress factors inducing C3-photosynthesis to CAM in facultative CAM plants and assess progress in the analysis of anatomical, physiological, metabolic and molecular differences for facultative CAM plants between the C3 and CAM mode. We also review their implications on horticultural crops and address directions for future research.
2. Signaling Stress Factors
CAM is a plastic photosynthetic adaptation found in plants in abiotic stress environments (such as drought, salinity, extreme temperature, etc.) [13]. Environmental, hormonal and circadian changes can regulate the CAM expression in facultative CAM species [22]. In facultative CAM species, photosynthesis can switch from C3 to CAM modes after induction by abiotic stress, such as atmospheric CO2 concentration, drought, salinity, photoperiod and light [12,23,24,25]. For instance, M. crystallinum switches its photosynthetic mode from C3 to CAM under water or salinity stress [7]. Light intensity and quality also play a crucial role for the C3–CAM transition [12,26]. All these signaling factors connect via a closed network and directly or indirectly affect each other [2].
The exogenous application of ethylene or abscisic acid (ABA) could induce the C3–CAM transition in a few facultative CAM species [27]. The degree of CAM expression was positively correlated with ABA and trans-zeatin, but negatively correlated with cytokinins and jasmonic acid (JA) [28,29]. Exogenous hydrogen peroxide (H2O2) and root signaling also could induce the C3–CAM transition in M. crystallinum, respectively [30,31,32].
3. Anatomical Variations during the C3–CAM Shift in Facultative CAM Plants
Leaf thickness is an important anatomical feature for CAM plants. CAM is often associated with succulent leaves; indeed, the tissue succulence of CAM species has been observed in many plant families, such as Crassulaceae, Orchidaceae and Clusiaceae [33,34,35,36]. Many arid CAM plants with succulent nature are beneficial as they store more water than C3 and C4 species [37]. A study reported, by analyzing the leaf thickness and leaf δ13C values in 173 tropical orchids, that the leaf was the thickest in the strong CAM species [36]. However, some plants with thinner leaves can also fix CO2 through the CAM pathway; for example, Dendrobium bigibbum (a CAM orchid) can yield δ13C with −11.9‰, despite the leaf thickness being only 0.79 mm [34], which proved that the assumption about obligate CAM species possessing more succulent leaves than facultative CAM species was not accurate [38]. In M. crystallinum, leaf succulence increased during the C3 to CAM transition after 5 days of the salt treatment [39], but some CAM species (e.g., bromeliads) do not have succulent photosynthetic organs [40,41]. Recently, it was also hypothesized that the evolution of facultative CAM plants did not require major changes in anatomy [42,43]. Winter thought that strong CAM plants needed significant anatomical modifications, whereas facultative or weak CAM plants may not require them, suggesting there may be no anatomical feature changes during the C3–CAM transition in facultative CAM plants [9]. Investigations showed that leaf anatomy was not correlated to CAM function in Yucca gloriosa (facultative CAM species, a C3+CAM hybrid species) [44], and the relationships between leaf anatomy and degree of CAM expression were not very close [45]. Herrera [39] reported that less succulence is not a typical feature for facultative CAM plants. Thus, the leaf thickness, as an indicator, cannot completely distinguish between plant species, suggesting that the relationship between anatomical leaf features and CAM expression requires further investigation.
That being said, there may be some changes in vacuole and chloroplast anatomy during the photosynthesis shift from C3 to CAM. Malic acid accumulates and releases in the vacuole. More particularly, the fluidity of the tonoplast will reduce after the photosynthesis shift from C3 to CAM, in turn decreasing the vacuolar mobilization of malic acid [46], indicating that vacuole size may increase before the shift to CAM in the leaves [47]. Chloroplasts, the main sites of photosynthesis in plants, can regulate the facultative CAM plants to acclimate to high salinity environments [48], and show a severe thylakoid swelling at midday in CAM plants [49]. However, whether such changes in the ultrastructure level also occur in other facultative CAM species remains unclear. Young leaves in obligate CAM plants take up CO2 by C3 photosynthesis, while mature leaves take up CO2 by CAM. It is worth exploring the changes in ultrastructure level (i.e., vacuole and chloroplast) that occur throughout the development process of obligate CAM plants.
4. Physiological Mechanisms during the C3–CAM Shift in Facultative CAM Plants
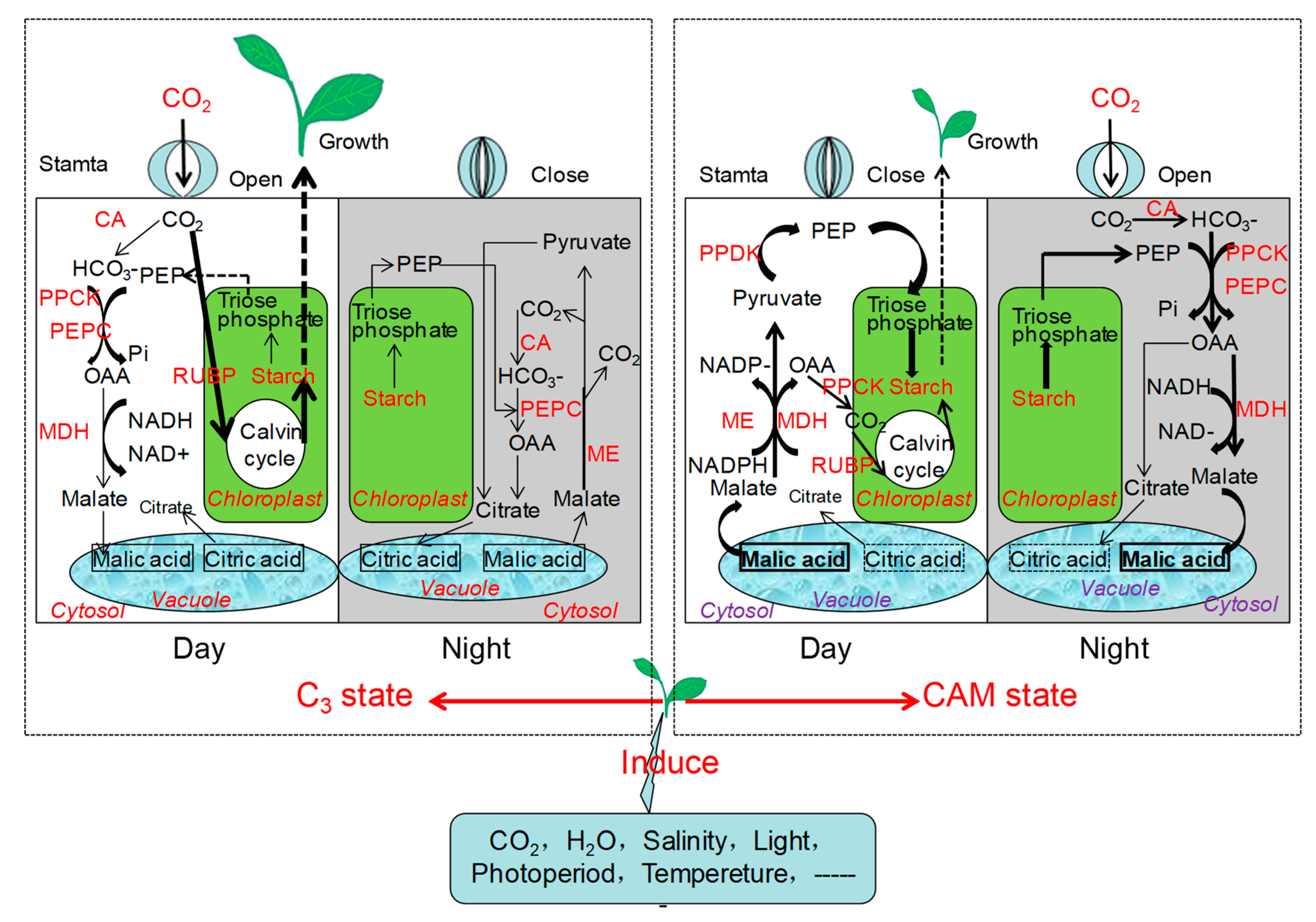
Compared to C3 and C4 metabolism, CAM is characterized by CO2 uptake at night and stomatal closure during the day. For facultative CAM species, the C3 photosynthesis pathway is used to fix CO2 when the plants are under well-conditions. However, the photosynthesis can shift to the CAM mode once induced by abiotic stress; stomata open at night and atmospheric CO2 is absorbed by phosphoenolpyruvate carboxylase (EC 4.1.1.31; PEPC) via oxaloacetate into malic acid. On the following day, stomata close and malic acid is released from the vacuoles and decarboxylated, while CO2 is refixed by the Calvin cycle [1]. Therefore, in facultative CAM plants, the CO2 gas exchange exhibits a difference between daytime and nighttime, as well as the changes in the activities of PEPC and Rubisco, which are important features to distinguish the C3 from the CAM mode. Figure 1 shows the physiological and metabolic shift of the facultative CAM plants from C3 to CAM photosynthesis.
Since this C3 to CAM transition was first recognized, M. crystallinum was used as a model plant to examine the associated changes in enzyme activities [50]. PEPC, one of the key enzymes, is involved in primary carboxylation during both CAM and C4 photosynthesis. PEPC is widely distributed across plants, algae and bacterial species; it catalyzes the irreversible β-carboxylation of phosphoenolpyruvate (PEP) in the presence of HCO3− to yield oxaloacetate (OOA) and Pi. In CAM and C4 photosynthesis, this enzyme is responsible for the primary fixation of inorganic carbon. PEPC also is a major anaplerotic enzyme in most non-photosynthetic organs and the leaves of C3 plants [51,52]. Furthermore, the activities of many decarboxylating enzymes increase during the shift from C3 to CAM, such as the cytosolic NADP-malic enzyme (NADPME, EC 1.1.1.40), the mitochondrial NAD-malic enzyme (NADME, EC 1.1.1.38) and PEP carboxykinase (PEPCK). This increased enzymatic activity is thought to be an indicator of the start of CAM photosynthesis [22,50].
In the plant, reactive oxygen species (ROS), including the superoxide radicals (O2−), the hydroxyl radical (OH) and hydrogen peroxide (H2O2), are always formed in response to environmental stress [53,54]. Oxidative stress could lead to the photosynthesis switch from C3 to CAM in facultative CAM plants [55,56]. Enzymatic antioxidants and non-enzymatic antioxidants are involved to protect the plants from oxidative damage by the scavenging of ROS [57]. During the C3–CAM shift in facultative CAM plants (e.g., Sedum album) induced by water stress, antioxidative enzymes, such as superoxide dismutase (SOD), peroxidase (POD), ascorbate peroxidase (APX), catalase (CAT), etc. [4,58,59]. In M. crystallinum, FeSOD activity increases more rapidly during the first few days before CAM appearance; then, MnSOD and Cu/ZnSOD activity increases after CAM occurs, induced by salt [48,60]. CAT is not only responsible for the removal of H2O2, but presents diel fluctuations [58]. Non-enzymatic antioxidants constituted by low molecular metabolites include ascorbic acid (AsA), glutathione (GSH), carotenoids, γ-tocopherol, etc. [57]. In M. crystallinum, a transition from C3 to CAM induced by H2O2 or salinity, α-tocopherol, polyamines and proline showed accumulation and performed a crucial role in preventing oxidative damage [31,61,62]. Likewise, during the C3–CAM shift in Guzmania monostachia (a facultative CAM plant) induced by water stress under high light PFD, carotenoids were proven to play an important role in the ROS scavenging system [63].
5. Metabolic Mechanism during the C3–CAM Shift in Facultative CAM Plants
CAM was controlled by the circadian clock and metabolites. Organic acids and carbohydrates are two kinds of major metabolites in facultative CAM plants [64]. Large diel changes in malic acid and transitory starch are important characteristics of CAM photosynthetic activity [65,66,67]. Plants, once stressed by adverse environmental factors, trigger their ABA- and Ca2+-dependent signaling pathways, which leads to metabolite changes [28]. These diurnal changes in malic acid and/or citric acid regulate the expression of CAM photosynthesis [68], but the acid accumulation requires metabolic reprogramming [69]. Metabolic fluxes modeling of a starch/sugar-malate cycle was established to test whether a C3–CAM continuum exists in CAM evolution [70].
Carbohydrates, including starch and sucrose or hexose, are produced by the CAM pathway at a high energetic cost [65]. They are converted into PEP and provide substrates for the nocturnal CO2 fixation [71]. In M. crystallinum, this transitory starch reserve is critical for CAM photosynthesis and the oscillation in starch levels between day and night (when the plant is in CAM mode) is 20% greater than in the C3 mode [72,73]. Starch-deficient mutants of this plant are characterized by their inability to photosynthesize in the CAM mode, as these mutant plants are deficient in leaf starch. It was reported that there was a transitory starch breakdown by the hydrolytic pathway when plants were in the C3 mode, but that a shift to the phosphorolytic pathway occurred during CAM photosynthesis [66]. In M. crystallinum, about half of the starch degraded during the night was used to supply PEP to ensure CAM photosynthesis, and the others were exported as soluble sugars used during plant respiration [74]. In another example, the Agave plant (Yucca aloifolia), a CAM plant, uses soluble sugars as a carbohydrates during CAM photosynthesis, while the C3 plant Y. filamentosa is thought to rely on starch pools. However, the diploid hybrid species Y. gloriosa, a facultative CAM species, relies on starch for carbohydrates (like its C3 parent Y. filamentosa), though many features are more similar to that of its CAM parent Y. aloifolia [75,76]. In facultative CAM plants, the proportion of CO2 taken up between nighttime and daytime is influenced by developmental and environmental factors (such as drought, salinity and extreme temperature) [72,77]. Various studies have revealed that many species express a low degree of CAM photosynthesis and a peak in C3-type isotope signatures. Generally, the δ13C value has more C3-type isotope signatures than CAM in facultative CAM plants. In S. nuttallianum, the δ13C value was −30.0‰ but △H+ was significant under controlled conditions [36]. C. minor can shift from C3 to CAM photosynthesis easily and reversibly, and its δ13C value was about −21‰ [78]. These studies showed that carbon was mostly fixed through C3 photosynthesis in facultative CAM plants, and dark CO2 fixation made up, at most, 30% [39].
6. Molecular Mechanisms during C3–CAM Shift in Facultative CAM Plants
The photosynthetic shift from C3–CAM is regulated by some enzymes and metabolite transporters, making this process a part of complex metabolic adaptations to environmental stress [22]. However, the relative expression of genes for enzymes and metabolites is strictly regulated by environmental factors. Genes involved in the shift of C3–CAM may be controlled by a co-expressed circadian master regulator [79]. Therefore, describing the molecular mechanism behind the C3–CAM shift is important to understand the evolution of CAM. Although every documented gene in the CAM also exists in the ancestral C3 species, the timing and magnitude are different between C3 and CAM species [80].
The PEPC gene is one of the most important genes in CAM and C4 photosynthesis. In the past decades, the presence and function of this gene have been investigated in many CAM and facultative CAM plants. PEPC is a ubiquitous enzyme in higher plants and belongs to a small multigene family with several PEPC isozymes [52,81]. Plant PEPC activity is regulated by PEPC kinase (PPCK) [82]. Boxall et al. found that silencing PEPC (Ppc1) in the obligate CAM species Kalanchoë laxiflora can prevent CO2 uptake and malate accumulation at night [83]. Here, the main molecular mechanisms that occur during the photosynthesis transition from C3 to CAM in facultative CAM plants are as follows.
7. DNA Level Regulation
8. Transcriptional Regulation
Between obligate CAM and C3 photosynthesis plants, all of the CAM-related genes exist in ancestral C3 species, but transcriptional regulation cascades are very important for the C3–CAM transition, especially the expression of CAM-specific genes [86,87]. Some enzymes and genes that are involved in some facultative CAM plants during the photosynthesis from C3 to CAM are shown in Table 1. The expression of CAM-specific PEPC, NAD-GAPDH and PPDK is important for the onset during the CAM induction; for example, PEPC mRNA accumulation occurs within 2–3 h, stressed by salinity in M. crystallinum or K. blossfeldiana [27,88,89]. Additionally, there are discrete changes observed in protein sequences [9,75,90]; the promoter regions of these CAM-specific genes contain GT motifs, which may function in light-responsive or ABA-mediated gene expression events [91,92]. The promoter sequences were different in the CAM-specific PEPC (Ppcl) and the C3 “housekeeping” PEPC, that is the former containing TATA and CAAT box motifs, but there was absence in the latter [91]. For CAM-specific genes, there may be the same common cis-acting regulatory elements for regulating the stress-induced expression patterns in different facultative CAM plants. During CAM induction, the distal regions between −977 and −721 control the expression of Ppcl for salt-responsiveness, while the regions between −735 and −675 control the expression of NADGAPDH(GapC1) induced by salt [93]. Therefore, the Ppcl and GapCl promoters in the distal regions share multiple consensus binding sites of transcription factor which control the salt-inducible gene expression. In Kalanchoë, PPC1 is essential for the practice of CAM [83]. In facultative CAM plant Talinum triangulare, transcriptional regulation of the C3–CAM transition revealed that the levels of the CAM-cycle enzyme transcripts are increased in response to drought stress [6]. In facultative CAM plants, during CAM induction induced by abiotic stress, a few transcription factors may control some transcriptional activation events to improve their tolerance [6,94].
Recently, several transcription factors (TF) families take part in regulating CAM induction by salinity or drought stress, such as AP2/ERF, MYB, WRKY, NAC, NF-Y, bZIP and McHB7 [93,95,96,97,98]. In T. triangulare, during CAM induction stressed by ABA, transcription factors such as HSFA2, NF-YA9 and JMJ27 were identified as regulators for the CAM induction [21]. Cushman and Bohnert [99] demonstrated that one factor (designated PCAT-1) binds in the Ppcl promoter, and the PCAT-1 expressed is abundant and may play an important role in the assembly of active transcription complexes during the photosynthesis shift from C3 to CAM.

{kind=link}
Table 1.
Some enzymes and genes involved in some facultative CAM plants during the photosynthesis shift from C3 to CAM.
Table 1.
Some enzymes and genes involved in some facultative CAM plants during the photosynthesis shift from C3 to CAM.
| Enzyme | Gene | Source/Species | Inducer | Reference |
|---|---|---|---|---|
| Phosphenolpyruvate carboxylase | Ppc1 | M. crystallinum | salt, ABA, drought, cytokinin | [25,100] |
| Kb-1, Kb-2 | K. blossfeldiana | short-day, drought | [101] | |
| Ppc 3 | T. triangulare | ABA | [21] | |
| C3-type PEPCs | C. minor | drought | [102] | |
| Alpha Carbonic Anhydrase 1 | ACA1 | T. triangulare | ABA | [21] |
| Beta Carbonic Anhydrase 5 | BCA5 | T. triangulare | ABA | [21] |
| Malic Enzymes | MEs | T. triangulare | ABA | [21] |
| PEPC Kinase | PPCK1 | M. crystallinum T. triangulare | Salt ABA | [21,103,104] |
| Pyruvate orthophosphate dikinase(PPDK) | Ppdk1 | M. crystallinum T. triangulare | salt, ABA | [21,105] |
| Enolase | Pgh1 | M. crystallinum | salt, drought, cold, hypoxia, ABA, 6-BA | [106] |
| phosphoglyceromutase (PGM) | Pgm1 | M. crystallinum | salt, drought, ABA, 6-BA | [107] |
| GAD-Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) | GapC1 | M. crystallinum | salt | [93,108] |
| NADP-Malic enzyme | Mod1 | M. crystallinum T. triangulare | salt ABA | [21,109] |
| Mod4 | T. triangulare | ABA | [21] | |
| NADP-Malate dehydrogenase | MDH1 | M. crystallinum | salt | [80] |
| NAD-Malate dehydrogenase | MDH2 | M. crystallinum | salt | [110] |
| H+-ATPase. c subunit | Atpvc | M. crystallinum, K. daigremontiana | salt, ABA, light | [111,112,113] |
| H+-ATPase, E subunit | AtpvE | M. crystallinum | salt | [114] |
| SNF1 kinase | MK9 | M. crystallinum | salt | [115] |
| RNA-binding protein | Rbp1 | M. crystallinum | salt | [116] |
| Ribosome inactivating proteins | Rip1 | M. crystallinum | salt | [117] |
9. Post-Transcriptional Regulation
Post-transcriptional regulations also take part in regulating CAM expression, such as that Ppc1 may facilitate long-term CAM build up by increasing mRNA stability in M. crystallinum during salt stress [118,119]. In M. crystallinum, many cDNA libraries were constructed with different tissues and stress treatments [120]. Large-scale steady-state mRNA abundance was found to change significantly during CAM induction in plants subjected to salinity-related stress [121]. There was at least one CAM-specific PEPC isoform (Ppc1) responsible for CO2 fixation during the night, which is more abundant than during the day and is responsible for CAM expression [75,122]. During the photosynthesis shifts from C3 to CAM in T. triangulare, PEPC isoform 1(Ppc1) and isoform 3 (Ppc3) transcript abundance increased and PPDK transcripts started to accumulate after 80 min, but the Alpha Carbonic Anhydrase 1 (ACA1) transcript decreased [21]. When M. crystallinum was treated for seven days with salt, Ppc1 and PPCK1 were up-regulated in guard cells [104].
Omics approaches were used by many scientists to explore the underlying molecular mechanism during the transition from C3 to CAM [6,123]. Among monocot species, the partial transcriptomes or genomes in the genus Phalaenopsis have been characterized [124]. The rapid reversible C3–CAM shift in the genus Clusia is based on the C3 isoform of PEPC (a housekeeping gene) [102]. The post-transcriptional regulation of photosynthetic genes is a key driver of C4 leaf ontogeny, determined by using exon–intron split analysis [125]. In order to explore the complex regulatory mechanisms, Heyduk et al. elaborated on a comparative analysis with closely related C3 and CAM species [86]. Aside from PEPC, other enzymes are also important for CAM photosynthesis, which are the products of isogenes. Other studies showed that following CAM induction by salinity-related stress, several CAM-related glycolysis/gluconeogenesis genes showed increasing transcript abundance [106,107,108]. In order to identify the genes involved during the transition from C3 to CAM in M. crystallinum, genomics and transcriptomics analyses were combined; twenty genes encoding six main enzymes were identified and one of four MDH genes presented a specific function in CAM photosynthesis [126]. Many CAM-related starch synthesis/degradation genes have been identified in M. crystallinum. During the photosynthesis shift from C3 to CAM, ADP glucose pyrophosphorylase small (Agp1 and Agp2) and large subunit (Agp3) catalyzed the starch biosynthesis and showed an increase in mRNA expression; additionally, there were three genes including AmyA1 (α-amylase isogene), AmyB1 and AmyB2 which exhibited increased remarkable mRNA abundance during nocturnal starch degradation [121].
Many microRNAs (miRNAs) also play a regulatory role in CAM photosynthesis in the leaves of obligate CAM species, such as Ananas comosus [127]. MiRNAs in M. crystallinum seedlings under salinity-related stress were analyzed by RNA sequencing and were found to be involved in the post-transcriptional regulation of salt tolerance [128]. Hu et al. identified some miRNAs that were involved in the regulation of CAM in Kalanchoë and found that the miR530-TZPs module regulates CAM-related gene expression [129].
10. Protein Level Regulation
Regulatory proteins exist in C3, CAM and C4 species and are essential to the C3–CAM photosynthesis shift in facultative CAM plants. In M. crystallinum, salt-stressed, heat shock proteins and early light-inducible proteins were found increased in the cDNA libraries, and a few proteins were found increased in guard cells but decreased in mesophyll cells [130,131]. There were seven proteins with increased expression and four proteins with decreased expression in M. crystallinum induced by salt [98]. Additionally, new major phosphorylation events during the transition from C3 to CAM stressed by salt were identified and characterized using proteomics and phosphoproteomics [132].
11. Implications in Horticultural Crops
CAM photosynthesis enables plants to assimilate carbon under environmental stress conditions. In contrast to C3 and C4 plants, obligate CAM plants have a higher transpiration efficiency and a lower photosynthetic rate; hence, obligate CAM plants often grow more slowly than their C3 and C4 counterparts [133]. Thus, CAM is not the best choice for highly productive plants [14]. However, facultative CAM plants can function in a C3 mode to increase photosynthetic rates and growth when there are no physical limitations present, and shift to the CAM mode to decrease water loss and overcome environmental stressors. For facultative CAM plants, the photosynthesis switching between C3 and CAM has important ecological implications. Therefore, engineering CAM into C3 crops can improve their WUE and sustain crop productivity in hot and dry climates [15,16,134].
CAM plants are widely distributed within the plant kingdom, i.e., 343 genera in 34 families, approximately 6.5% of flowering plant species [135]. In fact, most crops practice either C3 or C4 photosynthesis, but not CAM photosynthesis, such as wheat, rice and maize, which have higher production. However, many horticultural plants belong to obligate CAM plants or facultative CAM plants; some of them (such as pineapple, an obligate CAM plant, practice a facultative C3/CAM metabolism in the first 2 months of growth) can also be very productive when cultivated under well-conditions [55]. However, there was no exact number in facultative CAM plants. Winter (2019) reported that facultative CAM plants exist in at least 15 families, and he thought there may be over 1000 facultative CAM species in Aizoaceae alone [9]. Many orchids (Dendrobium spp., Oncidium spp. and Phalaenopsis spp.) [52,124,136,137], with higher ornamental values or medicinal values, were identified as facultative CAM plants since they could switch the pathway between C3 and CAM according to the environmental condition. For example, D. officinale, an important traditional herb with higher commercial value in China, uses the facultative CAM pathway to increase its drought tolerance [138]. Many species of Portulacaceae belong to facultative CAM plants [9]. Jatropha curcas, an oil crop, could also practice CAM photosynthesis for survival in response to environmental stress [139]. Thus, this plastic photosynthetic adaptation results in important implications for many horticultural crops.
12. Future Perspectives
For the facultative CAM plants, the photosynthesis switches from C3 to CAM have important ecological implications. Facultative CAM can prevent CO2 loss and favors plant growth and reproduction in responses to environmental stress. During the C3–CAM shift in facultative CAM plants, although there are some progresses in anatomy, physiological, metabolic and molecular properties of facultative CAM plants, in the near future, there are still some works that should be carried out. First, the argument on anatomical variations should be comprehensive and ongoing, and studied with more genera that include obligate C3, facultative CAM and obligate CAM plants. For example, many characters and mechanisms can be explored in the genera Clusia (Clusiaceae), Dendrobium and Oncidium (Orchidaceae) and Yucca (Asparagaceae), which are well known for containing the obligate C3, facultative CAM and obligate CAM plants in a single genus [19,76,102,136]. Second, molecular mechanisms underlying the transition from C3 to CAM in plants is still limited. Recently, omics approaches including transcriptomic, genomic, proteomic, metabolomics and ionomics were used frequently to reveal the molecular changes during the C3–CAM transition (See Table 2). For example, altered gene regulatory networks and expression profiles were found in the transition from C3 to CAM in Erycina (Orchidaceae) and Yucca (Asparagaceae) [76,86], which will benefit the clarification of the key molecular switches underlying this transition of C3 to CAM in facultative CAM plants. Third, understanding the functional genomics of CAM plants is important to elucidate the relationship between genotype and phenotype [9]. Synthetic biology toolboxes, such as the CRISPR/Cas 9 system which was confirmed to be effective for genome editing in K. fedtschenkoi, will accelerate the ongoing research about CAM and the C3 to CAM transition mechanisms [17,140]. Assisted with all this prior knowledge, we should obtain more genetic transformation information on facultative CAM plants and transfer more CAM-related genes into C3 types of horticultural crops in future.
Table 2.
Omics approaches involved to reveal the molecular changes during the C3–CAM transition in facultative CAM plants.
Table 2.
Omics approaches involved to reveal the molecular changes during the C3–CAM transition in facultative CAM plants.
| Omics Approaches | Source/Species | Photosynthesis Type | Year | References |
|---|---|---|---|---|
| Proteomics, Metabolomics | M. crystallinum | facultative CAM plants | 2013 | [141] |
| Transcriptomics | M. crystallinum | facultative CAM plants | 2015 | [142] |
| Metabolomics | M. crystallinum | facultative CAM plants | 2015 | [143] |
| Transcriptomics | M. crystallinum | facultative CAM plants | 2015 | [144] |
| Proteomics, Ionomics | M. crystallinum | facultative CAM plants | 2016 | [145] |
| Transcriptomics, Metabolomics | T. triangulare | facultative CAM plants | 2016 | [6] |
| Transcriptomics | D. catenatum | facultative CAM plants | 2016 | [124] |
| Transcriptomics | Agave (CAM), Polianthes (weak CAM), Manfreda (CAM), Beschorneria (weak CAM) | CAM plants | 2018 | [95] |
| Transcriptomics | D. catenatum | facultative CAM plants | 2018 | [137] |
| Transcriptomics | Erycina pusilla (CAM), Erycina crista-galli(C3), | CAM plants, C3 plants | 2019 | [86] |
| Transcriptomics, Metabolomics | T. triangulare | facultative CAM plants | 2019 | [21] |
| Metabolomics Transcriptomics | Y. gloriosa (C3+ CAM), Y. filamentosa (C3), Y. aloifolia (CAM) | facultative CAM plants, C3 plants, obligate CAM plants | 2019 | [76] |
| Genomics | Sedum album | facultative CAM plants | 2019 | [123] |
| Transcriptomics | M. crystallinum | facultative CAM plants | 2020 | [104] |
| Proteomics, Metabolomics | M. crystallinum | facultative CAM plants | 2021 | [131] |
| Proteomics | M. crystallinum | facultative CAM plants | 2021 | [98] |
| Proteomics, Phosphoproteomics | M. crystallinum | facultative CAM plants | 2022 | [132] |
| Transcriptomics | Tamarix ramosissima | facultative CAM plants | 2022 | [146] |
| Transcriptomics Genomics | M. crystallinum | facultative CAM plants | 2022 | [126] |
| Transcriptomics | 11 species of Agavoideae | facultative CAM plants, C3 plants, obligate CAM plants | 2022 | [147] |
Author Contributions
Conceptualization, Z.Z. and S.Q.; writing and revision of the manuscript, S.Q.; revision and editing of the manuscript, K.X.; Y.Y. and Q.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (31560567), National Natural Science Foundation of Guangxi (2020GXNSFAA297260), Start-up Fund of Innovation Team of Guangxi Academy of Sciences for Innovation and Utilization of Germplasm in Horticultural Crops (CQZ-E-1919), Fundamental Research Fund of Guangxi Institute of Botany (23011) and the fund of Guangxi Key Laboratory of Plant Functional Phytochemicals and Sustainable Utilization (ZRJJ2022-5 and ZRJJ2023-1).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ABA | Abscisic acid |
| CAM | Crassulacean acid metabolism |
| APX | Ascorbate peroxidase |
| CAT | Catalase |
| H2O2 | Hydrogen peroxide |
| MDH | Malate dehydrogenase |
| ME | Malic enzyme; |
| NO | Nitric oxide |
| PEPC | Phosphoenolpyruvate carboxylase |
| PEPCK | PEP carboxykinase |
| POD | Peroxidase |
| PFD | Photon flux density |
| Rubisco | Ribulosebisphosphate carboxylase/oxygenase |
| SOD | superoxide dismutase (SOD) |
| TF | Transcription factors |
| TZPs | Tandem zinc knuckle/PLU3 domain encoding genes |
| WUE | Water-use efficiency |
References
- Osmond, C.B. Crassulacean acid metabolism: A curiosity in context. Annu. Rev. Plant Physiol. 1978, 29, 379–414. [Google Scholar] [CrossRef]
- Lüttge, U. Ecophysiology of Crassulacean acid metabolism (CAM). Ann. Bot. 2004, 93, 629–652. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.; von Willert, D.J. NaCl-induzierter Crassulaceen säurestoffwechsel bei Mesembryanthemum crystallinum. Z. Pflanzenphysiol. 1972, 67, 166–170. [Google Scholar] [CrossRef]
- Castillo, F.J. Antioxidative protection in the inducible CAM plant Sedum album L. following the imposition of severe water stress and recovery. Oecologia 1996, 107, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Barrera Zambrano, V.A.; Lawson, T.; Olmos, E.; Fernández-García, N.; Borland, A.M. Leaf anatomical traits which accommodate the facultative engagement of Crassulacean acid metabolism in tropical trees of the genus Clusia. J. Exp. Bot. 2014, 65, 3513–3523. [Google Scholar] [CrossRef] [Green Version]
- Brilhaus, D.; Bräutigam, A.; Mettler-Altmann, T.; Winter, K.; Weber, A.P. Reversible burst of transcriptional changes during induction of Crassulacean acid metabolism in Talinum triangulare. Plant Physiology 2016, 170, 102–122. [Google Scholar] [CrossRef] [Green Version]
- Winter, K.; Holtum, J.A.M. Environment or development? Lifetime net CO2 exchange and control of the expression of crassulacean acid metabolism in Mesembryanthemum crystallinum. Plant Physiol. 2007, 143, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Winter, K.; Garcia, M.; Holtum, J.A.M. On the nature of facultative and constitutive CAM: Environmental and developmental control of CAM expression during early growth of Clusia, Kalanchoë, and Opuntia. J. Exp. Bot. 2008, 59, 1829–1840. [Google Scholar] [CrossRef]
- Winter, K. Ecophysiology of constitutive and facultative CAM photosynthesis. J. Exp. Bot. 2019, 70, 6495–6508. [Google Scholar] [CrossRef]
- Johnson, G.N.; Lawson, T.; Murchie, E.H.; Raines, C. Photosynthesis in variable environments. J. Exp. Bot. 2015, 66, 2371–2372. [Google Scholar] [CrossRef] [Green Version]
- DePaoli, H.C.; Borland, A.M.; Tuskan, G.A.; Cushman, J.C.; Yang, X.H. Synthetic biology as it relates to CAM photosynthesis: Challenges and opportunities. J. Exp. Bot. 2014, 65, 3381–3393. [Google Scholar] [CrossRef] [Green Version]
- Grams, T.E.E.; Thiel, S. High light-induced switch from C3-photosynthesis to crassulacean acid metabolism is mediated by UV-A/blue light. J. Exp. Bot. 2002, 53, 1475–1483. [Google Scholar] [CrossRef]
- Cushman, J.C. Crassulacean acid metabolism. A plastic photosynthetic adaptation to arid environments. Plant Physiology 2001, 127, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Tay, S.; He, J.; Yam, T.W. CAM plasticity in epiphytic tropical orchid species responding to environmental stress. Bot. Stud. 2019, 60, 7. [Google Scholar] [CrossRef] [PubMed]
- Borland, A.M.; Hartwell, J.; Weston, D.J.; Schlauch, K.A.; Tschaplinski, T.J.; Tuskan, G.A.; Yang, X.; Cushman, J.C. Engineering crassulacean acid metabolism to improve water-use efficiency. Trends Plant Sci. 2014, 19, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.H.; Cushman, J.C.; Borland, A.M.; Edwards, E.J.; Wullschleger, S.D.; Tuskan, G.A.; Owen, N.A.; Griffiths, H.; Smith, J.A.C.; Paoli, H.C.D.; et al. A roadmap for research on crassulacean acid metabolism (CAM) to enhance sustainable food and bioenergy production in a hotter, drier world. New Phytol. 2015, 207, 491–504. [Google Scholar] [CrossRef]
- Liu, D.G.; Chen, M.; Mendoza, B.; Cheng, H.; Hu, R.B.; Li, L.L.; Trinh, C.T.; Tuskan, G.A.; Yang, X.H. CRISPR/Cas9-mediated targeted mutagenesis for functional genomics research of crassulacean acid metabolism plants. J. Exp. Bot. 2019, 70, 6621–6629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.; Lee, S.; Choi, W.; Yim, W.; Cushman, J. Laying the foundation for crassulacean acid metabolism (CAM) biodesign: Expression of the C4 metabolism cycle genes of CAM in Arabidopsis. Front. Plant Sci. 2019, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Silvera, K.; Neubig, K.M.; Whitten, W.M.W.; Williams, N.H.; Winter, K.; Cushman, J.C. Evolution along the crassulacean acid metabolism continuum. Funct. Plant Biol. 2010, 37, 995–1010. [Google Scholar] [CrossRef] [Green Version]
- Hartwell, J.; Dever, L.V.; Boxall, S.F. Emerging model systems for functional genomics analysis of crassulacean acid metabolism. Curr. Opin. Plant Biol. 2016, 31, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maleckova, E.; Brilhaus, D.; Wrobel, T.J.; Weber, A.P.M. Transcript and metabolite changes during the early phase of abscisic acid-mediated induction of crassulacean acid metabolism in Talinum triangulare. J. Exp. Bot. 2019, 70, 6581–6596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cushman, J.C.; Bohnert, H.J. Crassulacean acid metabolism: Molecular Genetics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 305–332. [Google Scholar] [CrossRef] [PubMed]
- Lüttge, U. Carbon dioxide and water demand. Crassulacean acid metabolism (CAM), a versatile ecological adaptation exemplifying the need for integration in eco-physiological work. New Phytol. 1987, 106, 593–629. [Google Scholar] [CrossRef]
- Borland, A.M.; Griffiths, H. The regulation of CAM and respiratory recycling by water supply and light regime in the C3-CAM intermediate Sedum telephium. Funct. Ecol. 1990, 4, 33–39. [Google Scholar] [CrossRef]
- Cushman, J.C.; Meyer, G.; Michalowski, C.B.; Schmitt, J.M.; Bohnert, H.J. Salt stress leads to differential expression of two isogenes of phosphoenolpyruvate carboxylase during crassulacean acid metabolism induction in the common ice plant. Plant Cell 1989, 1, 715–725. [Google Scholar]
- Broetto, F.; Lüttge, U.; Ratajczak, R. Influence of light intensity and salt-treatment on mode of photosynthesis and enzymes of the antioxidative response system of Mesembryanthemum crystallinum. Funct. Plant Biol. 2002, 29, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Taybi, T.; Sotta, B.; Gehrig, H.; Giiclii, S.; Kluge, M.; Brulfert, J. Differential effects of abscisic acid on phosphoenolpyruvate carboxylase and CAM operation in Kalanchoe blossfeldiana. Bot. Acta 1995, 108, 240–246. [Google Scholar] [CrossRef]
- Freschi, L.; Rodrigues, M.A.; Domingues, D.S.; Purgatto, E.; Van Sluys, M.A.V.; Magalhaes, J.R.; Kaiser, W.M.; Mercier, H. Nitric oxide mediates the hormonal control of crassulacean acid metabolism expression in young Pineapple plants. Plant Physiology 2010, 152, 1971–1985. [Google Scholar] [CrossRef] [Green Version]
- Wakamatsu, A.; Mori, I.C.; Matsuura, T.; Taniwaki, Y.; Yoshida, R. Possible roles for phytohormones in controlling the stomatal behavior of Mesembryanthemum crystallinum during the salt-induced transition from C3 to crassulacean acid metabolism. J. Plant Physiol. 2021, 262, 153448. [Google Scholar] [CrossRef]
- Ślesak, I.; Karpinska, B.; Surówka, E.; Miszalski, Z.; Karpinski, S. Redox changes in the chloroplast and hydrogen peroxide are essential for regulation of C3-CAM transition and photooxidative stress responses in the facultative CAM plant Mesembryanthemum crystallinum L. Plant Cell Physiol. 2003, 44, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Surówka, E.; Dziurka, M.; Kocurek, M.; Goraj, S.; Rapacz, M.; Miszalski, Z. Effects of exogenously applied hydrogen peroxide on antioxidant and osmoprotectant profiles and the C3-CAM shift in the halophyte Mesembryanthemum crystallinum L. J. Plant Physiol. 2016, 200, 102–110. [Google Scholar] [CrossRef]
- Eastmond, P.J.; Ross, J.D. Evidence that the induction of crassulacean acid metabolism by water stress in Mesembryanthemum crystallinum (L.) involves root signalling. Plant Cell Environ. 1997, 20, 1559–1565. [Google Scholar] [CrossRef]
- Teeri, J.A.; Tonsor, S.J.; Turner, M. Leaf thickness and carbon isotope composition in the Crassulaceae. Oecologia. 1981, 50, 367–369. [Google Scholar] [CrossRef]
- Winter, K.; Wallace, B.J.; Stocker, G.C.; Roksandic, Z. Crassulacean acid metabolism in Australian vascular epiphytes and some related species. Oecologia 1983, 57, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Borland, A.M.; Técsi, L.I.; Leegood, R.C.; Walker, R.P. Inducibility of crassulacean acid metabolism (CAM) in Clusia species; physiological/biochemical characterisation and intercellular localisation of carboxylation processes in three species which show different degrees of CAM. Planta 1998, 205, 342–351. [Google Scholar] [CrossRef]
- Silvera, K.; Santiago, L.S.; Winter, K. Distribution of crassulacean acid metabolism in orchids of Panama: Evidence of selection of weak and strong modes. Funct. Plant Biol. 2005, 32, 397–407. [Google Scholar] [CrossRef]
- Heyduk, K. Evolution of crassulacean acid metabolism in response to the environment: Past, present, and future. Plant Physiol. 2022, 190, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Veste, M.; Herppich, W.B.; von Willert, D.J. Variability of CAM in leaf deciduous succulents from the Succulent Karoo (South Africa). Basic Appl. Biol. 2001, 2, 283–288. [Google Scholar]
- Herrera, A. Crassulacean acid metabolism and fitness under water deficit stress: If not for carbon gain, what is facultative CAM good for? Ann. Bot. 2009, 103, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Q.J.; Tan, B.; Kelley, T.M.; Tian, J.K.; Chen, S.X. Physiological changes in Mesembryanthemum crystallinum during the C3 to CAM transition induced by salt stress. Front. Plant Sci. 2020, 11, 283. [Google Scholar] [CrossRef] [Green Version]
- De-Santo, A.V.; Alfani, A.; Russo, G.; Fioretto, A. Relationship between CAM and succulence in some species of Vitaceae and Piperaceae. Bot. Gaz. 1983, 144, 342–346. [Google Scholar] [CrossRef]
- Edwards, E.J. Evolutionary trajectories, accessibility and other metaphors: The case of C4 and CAM photosynthesis. New Phytol. 2019, 223, 1742–1755. [Google Scholar] [CrossRef] [Green Version]
- Heyduk, K.; Moreno-Villena, J.J.; Gilman, I.S.; Christin, P.A.; Edwards, E.J. The genetics of convergent evolution: Insights from plant photosynthesis. Nat. Rev. Genet. 2019, 20, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Heyduk, K.; Ray, J.N.; Leebens-Mack, J. Leaf anatomy is not correlated to CAM function in a C3+CAM hybrid species, Yucca gloriosa. Ann. Bot. 2021, 127, 437–449. [Google Scholar] [CrossRef] [Green Version]
- Herrera, A. Are thick leaves, large mesophyll cells and small intercellular air spaces requisites for CAM? Ann. Bot. 2020, 125, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Kliemchen, A.; Schomburg, M.; Galla, H.J.; Lüttge, U.; Kluge, M. Phenotypic changes in the fluidity of the tonoplast membrane of crassulacean acid metabolism plants in response to temperature and salinity stress. Planta 1993, 189, 403–409. [Google Scholar] [CrossRef]
- Jones, M.B. Effect of leaf age on leaf resistance and CO2 exchange of the CAM plant Bryophyllum fedtschenkoi. Planta 1975, 123, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Ślesak, I.; Miszalski, Z.; Karpinska, B.; Niewiadomska, E.; Ratajczak, R.; Karpinski, S. Redox control of oxidative stress responses in the C3–CAM intermediate plant Mesembryanthemum crystallinum. Plant Physiol. Biochem. 2002, 40, 669–677. [Google Scholar] [CrossRef]
- Niewiadomska, E.; Bilger, W.; Gruca, M.; Mulisch, M.; Miszalski, Z.; Krupinska, K. CAM-related changes in chloroplastic metabolism of Mesembryanthemum crystallinum L. Planta 2011, 233, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Holtum, J.A.M.; Winter, K. Activities of enzymes of carbon metabolism during the induction of crassulacean acid metabolism in Mesembryanthemum crystallinum. Planta 1982, 155, 8–16. [Google Scholar] [CrossRef]
- O’Leary, B.; Park, J.; Plaxton, W.C. The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase): Recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs. Biochem. J. 2011, 436, 15–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvera, K.; Winter, K.; Rodriguez, B.L.; Albion, R.L.; Cushman, J.C. Multiple isoforms of phosphoenolpyruvate carboxylase in the Orchidaceae (subtribe Oncidiinae): Implications for the evolution of Crassulacean acid metabolism. J. Exp. Bot. 2014, 65, 3623–3636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Aragón, C.; Carvalho, L.; González, J.; Escalona, M.; Amâncio, S. The physiology of ex vitro pineapple (Ananas comosus L. Merr. var MD-2) as CAM or C3 is regulated by the environmental conditions. Plant Cell Rep. 2012, 31, 757–769. [Google Scholar] [CrossRef]
- Aragón, C.; Pascual, P.; González, J.; Escalona, M.; Carvalho, L.; Amancio, S. The physiology of ex vitro pineapple (Ananas comosus L. Merr. var MD-2) as CAM or C3 is regulated by the environmental conditions: Proteomic and transcriptomic profiles. Plant Cell Rep. 2013, 32, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Niewiadomska, E.; Miszalski, Z.; Ślesak, I.; Ratajczak, R. Catalase activity during C3-CAM transition in Mesembryanthemum crystallinum L. leaves. Free Radic. Res. 1999, 31, 251–256. [Google Scholar] [CrossRef]
- Nosek, M.; Gawrońska, K.; Rozpądek, P.; Szechyńska-Hebda, M.; Kornaś, A.; Miszalski, Z. Withdrawal from functional crassulacean acid metabolism (CAM) is accompanied by changes in both gene expression and activity of antioxidative enzymes. J. Plant Physiol. 2018, 229, 151–157. [Google Scholar] [CrossRef]
- Miszalski, Z.; Slesak, I.; Niewiadomska, E.; Baczek-Kwinta, R.; Luttge, U.; Ratajczak, R. Subcellular localization and stress responses of superoxide dismutase isoforms from leaves in the C3-CAM intermediate halophyte Mesembryanthemum crystallinum L. Plant Cell Environ. 1998, 21, 169–179. [Google Scholar] [CrossRef]
- Sanada, Y.; Ueda, H.; Kuribayashi, K.; Andoh, T.; Hayashi, F.; Tamai, N.; Wada, K. Novel light-dark change of proline levels in halophyte (Mesembryanthemum crystallinum L.) and glycophytes (Hordeum vulgare L. and Triticum aestivum L.) leaves and roots under salt stress. Plant Cell Physiol. 1995, 36, 965–970. [Google Scholar] [CrossRef]
- Shevyakova, N.I.; Shorina, M.V.; Rakitin, V.Y.; Kuznetsov, V. Stress-dependent accumulation of spermidine and spermine in the halophyte Mesembryanthemum crystallinum under salinity conditions. Russ. J. Plant Physiol. 2006, 53, 739–745. [Google Scholar] [CrossRef]
- Abreu, M.E.; Carvalho, V.; Mercier, H. Antioxidant capacity along the leaf blade of the C3-CAM facultative bromeliad Guzmania monostachia under water deficit conditions. Funct. Plant Biol. 2018, 45, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Borland, A.M.; Taybi, T. Synchronization of metabolic processes in plants with crassulacean acid metabolism. J. Exp. Bot. 2004, 55, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Black, C.C.; Osmond, C.B. Crassulacean acid metabolism photosynthesis: ‘working the night shift’. Photosynth. Res. 2003, 76, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Weise, S.E.; van Wijk, K.J.; Sharkey, T.D. The role of transitory starch in C3, CAM, and C4 metabolism and opportunities for engineering leaf starch accumulation. J. Exp. Bot. 2011, 62, 3109–3118. [Google Scholar] [CrossRef] [PubMed]
- Habibi, G.; Hajiboland, R. Comparison of photosynthesis and antioxidative protection in Sedum album and Sedum stoloniferum (Crassulaceae) under water stress. Photosynthetica 2012, 50, 508–518. [Google Scholar] [CrossRef]
- Haider, M.S.; Barnes, J.D.; Cushman, J.C.; Borland, A.M. A CAM- and starch-deficient mutant of the facultative CAM species Mesembryanthemum crystallinum reconciles sink demands by repartitioning carbon during acclimation to salinity. J. Exp. Bot. 2012, 63, 1985–1996. [Google Scholar] [CrossRef]
- Winter, K.; Smith, J.A.C. CAM photosynthesis: The acid test. New Phytol. 2022, 233, 599–609. [Google Scholar] [CrossRef]
- Tay, I.Y.Y.; Odang, K.B.; Cheung, C.Y.M. Metabolic modeling of the C3-CAM continuum revealed the establishment of a starch/sugar-malate cycle in CAM evolution. Front. Plant Sci. 2021, 11, 573197. [Google Scholar] [CrossRef]
- Christopher, J.T.; Holtum, J.A.M. Patterns of carbon partitioning in leaves of crassulacean acid metabolism species during deacidification. Plant Physiol. 1996, 112, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Dodd, A.N.; Borland, A.M.; Haslam, R.P.; Griffiths, H.; Maxwell, K. Crassulacean acid metabolism: Plastic, fantastic. J. Exp. Bot. 2002, 53, 569–580. [Google Scholar] [CrossRef] [Green Version]
- Cushman, J.C.; Agarie, S.; Albion, R.L.; Elliot, S.M.; Taybi, T.; Borland, A.M. Isolation and characterization of mutants of common ice plant deficient in crassulacean acid metabolism. Plant Physiol. 2008, 147, 228–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borland, A.M.; Dodd, A.N. Carbohydrate partitioning in crassulacean acid metabolism plants: Reconciling potential conflicts of interest. Funct. Plant Biol. 2002, 29, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Abraham, P.E.; Yin, H.; Borland, A.M.; Weighill, D.; Lim, S.D.; De-Paoli, H.C.; Engle, N.; Jones, P.C.; Agh, R.; Weston, D.J.; et al. Transcript, protein and metabolite temporal dynamics in the CAM plant Agave. Nat. Plants 2016, 178, 16178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyduk, K.; Ray, J.N.; Ayyampalayam, S.; Moledina, N.; Borland, A.; Harding, S.A.; Tsai, C.J.; Leebens-Mack, J. Shared expression of crassulacean acid metabolism (CAM) genes pre-dates the origin of CAM in the genus Yucca. J. Exp. Bot. 2019, 70, 6597–6609. [Google Scholar] [CrossRef] [Green Version]
- Cushman, J.C.; Borland, A.M. Induction of Crassulacean acid metabolism by water limitation. Plant Cell Environ. 2002, 25, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Lüttge, U. Clusia: A Woody Neotropical Genus of Remarkable Plasticity and Diversity; Ecological Studies; Springer: Berlin, Germany, 2007; p. 194. [Google Scholar]
- Borland, A.M.; Hartwell, J.; Jenkins, G.I.; Wilkins, M.B.; Nimmo, H.G. Metabolite control overrides circadian regulation of phosphoenolpyruvate carboxylase knase and CO2 fixation in crassulacean acid metabolism. Plant Physiol. 1999, 121, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Cushman, J.C. Molecular cloning and expression of chloroplast NADP-malate dehydrogenase during crassulacean acid metabolism induction by salt stress. Photosynth. Res. 1993, 35, 15–27. [Google Scholar] [CrossRef]
- Gehrig, H.H.; Wood, J.A.; Cushman, M.A.; Virgo, A.; Cushman, J.C.; Winter, K. Research note: Large gene family of phosphoenolpyruvate carboxylase in the crassulacean acid metabolism plant Kalanchoe pinnata (Crassulaceae) characterised by partial cDNA sequence analysis. Funct. Plant Biol. 2005, 32, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Taybi, T.; Patil, S.; Chollet, R.; Cushman, J.C. A minimal serine/threonine protein kinase circadianly regulates phosphoenolpyruvate carboxylase activity in crassulacean acid metabolism-induced leaves of the common ice plant. Plant Physiol. 2000, 123, 1471–1482. [Google Scholar] [CrossRef] [Green Version]
- Boxall, S.F.; Kadu, N.; Dever, L.V.; Kneřová, J.; Waller, J.L.; Gould, P.J.D.; Hartwell, J. Kalanchoë PPC1 is essential for crassulacean acid metabolism and the regulation of core circadian clock and guard cell signaling genes[CC-BY]. Plant Cell 2020, 32, 1136–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyachenko, O.V.; Zakharchenko, N.S.; Shevchuk, T.V.; Bohnert, H.J.; Cushman, J.C.; Buryanov, Y.I. Effect of hypermethylation of CCWGG sequences in DNA of Mesembryanthemum crystallinum plants on their adaptation to salt stress. Biochem. Biokhimiia 2006, 71, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.C.; Li, C.H.; Lee, J.Y.; Yen, H.E. Cytosine methylation changes in the ice plant Ppc1 promoter during transition from C3 to crassulacean acid metabolism. Plant Sci. 2010, 178, 41–46. [Google Scholar] [CrossRef]
- Heyduk, K.; Hwang, M.; Albert, V.A.; Silvera, K.; Lan, T.Y.; Winter, K.; Leebens-Mack, J. Altered gene regulatory networks are associated with the transition from C3 to crassulacean acid metabolism in Erycina (Oncidiinae: Orchidaceae). Front. Plant Sci. 2019, 9, 2000. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.B.; Zhang, J.; Jawdy, S.; Sreedasyam, A.; Sreedasyam, A.; Lipzen, A.; Wang, M.; Ng, V.; Daum, C.; Keymanesh, K.; et al. Comparative genomics analysis of drought response between obligate CAM and C3 photosynthesis plants. J. Plant Physiol. 2022, 277, 153791. [Google Scholar] [CrossRef]
- Schmitt, J.M. Rapid concentration changes of phosphoenolpyruvate carboxylase mRNA in detached leaves of Mesembryanthemum crystallinum. Plant Cell Environ. 1990, 13, 845–850. [Google Scholar] [CrossRef]
- Brulfert, J.; Güclü, S.; Taybi, T.; Pierre, J.N. Enzymatic responses to water stress in detached leaves of the CAM plant Kalanchoë blossfeldiana Poelln. Plant Physiol. Biochem. 1993, 31, 491–497. [Google Scholar]
- Yang, X.H.; Hu, R.B.; Yin, H.F.; Jenkins, J.; Shu, S.Q.; Tang, H.B.; Liu, D.G.; Weighill, D.A.; Yim, W.C.; Ha, J.M.; et al. The Kalanchö genome provides insights into convergent evolution and building blocks of crassulacean acid metabolism. Nat. Commun. 2017, 8, 1899. [Google Scholar] [CrossRef] [Green Version]
- Cushman, J.C.; Vernon, D.M.; Bohnert, H.J. ABA and the transcriptional control of CAM induction during salt stress in the common ice plant. In Control of Plant Gene Expression; Verma, D.P.S., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 287–300. [Google Scholar]
- Schmitt, J.M.; Fißlthaler, B.; Sheriff, A.; Lenz, B.; Bäßler, M.; Meyer, G. Environmental control of CAM induction in Mesembryanthemum crystallinum—A role for cytokinin, abscisic acid and jasmonate? Crassulacean Acid Metabolism. Biochem. Ecophysiol. Evol. 1996, 114, 159–175. [Google Scholar]
- Schaeffer, H.J.; Forstheoefel, N.R.; Cushman, J.C. Identification of enhancer and silencer regions involved in salt-responsive expression of crassulacean acid metabolism (CAM) genes in the facultative halophyte Mesembryanthemum crystallinum. Plant Mol. Biol. 1995, 28, 205–218. [Google Scholar] [CrossRef]
- Amin, A.B.; Rathnayake, K.N.; Yim, W.C.; Garcia, T.M.; Wone, B.; Cushman, J.C.; Wone, B.W.M. Crassulacean acid metabolism abiotic stress-responsive transcription factors: A potential genetic engineering approach for improving crop tolerance to abiotic stress. Front. Plant Sci. 2019, 10, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyduk, K.; Ray, J.N.; Ayyampalayam, S.; Leebens-Mack, J. Shifts in gene expression profiles are associated with weak and strong crassulacean acid metabolism. Am. J. Bot. 2018, 105, 587–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moseley, R.C.; Mewalal, R.; Motta, F.; Tuskan, G.A.; Haase, S.; Yang, X.H. Conservation and diversification of circadian rhythmicity between a model crassulacean acid metabolism plant Kalanchoë fedtschenkoi and a model C3 photosynthesis plant Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1757. [Google Scholar] [CrossRef]
- Yin, H.B.; Guo, H.B.; Weston, D.J.; Borland, A.M.; Ranjan, P.; Abraham, P.E.; Jawdy, S.S.; Wachira, J.; Tuskan, G.A.; Tschaplinski, T.J.; et al. Diel rewiring and positive selection of ancient plant proteins enabled evolution of CAM photosynthesis in Agave. BMC Genom. 2018, 19, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.M.; Tan, B.W.; Zhu, D.; Dufresne, D.; Jiang, T.B.; Chen, S.X. Proteomics of Homeobox7 enhanced salt tolerance in Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2021, 22, 6390. [Google Scholar] [CrossRef]
- Cushman, J.C.; Bohnert, H.J. Salt stress alters A/T-rich DNA-binding factor interactions within the phosphoenolpyruvate carboxylase promoter from Mesembryanthemum crystallinum. Plant Mol. Biol. 1999, 20, 411–424. [Google Scholar] [CrossRef]
- McElwain, E.F.; Bohnert, H.J.; Thomas, J.C. Light moderates the induction of phosphoenolpyruvate carboxylase by NaCl and abscisic acid in Mesembryanthemum crystallinum. Plant Physiol. 1992, 99, 1261–1264. [Google Scholar] [CrossRef] [Green Version]
- Gehrig, H.; Taybi, T.; Kluge, M.; Brulfert, J. Identification of multiple PEPC isogenes in leaves of the facultative crassulacean acid metabolism (CAM) plant Kalanchoe blossfeldiana Poelln. cv.Tom Thumb. FEBS Lett. 1995, 377, 399402. [Google Scholar]
- Vaasen, A.; Begerow, D.; Hampp, R. Phosphoenolpyruvate carboxylase genes in C3, crassulacean acid metabolism (CAM) and C3/CAM intermediate species of the genus Clusia: Rapid reversible C3/CAM switches are based on the C3 housekeeping gene. Plant Cell Environ. 2006, 29, 2113–2123. [Google Scholar] [CrossRef]
- Li, B.; Chollet, R. Salt induction and the partial purification/characterization of phosphoenolpyruvate carboxylaseprotein-serine kinase from an inducible crassulacean acid-metabolism (CAM) plant, Mesembryanthemum crystallinum L. Arch. Biochem. Biophys. 1994, 314, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.W.; Yoo, M.J.; Zhu, D.; Noble, J.D.; Kelley, T.M.; Li, J.; Kirst, M.; Assmann, S.M.; Chen, S.X. Molecular changes in Mesembryanthemum crystallinum guard cells underlying the C3 to CAM transition. Plant. Mol. Biol. 2020, 103, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Fissßthaler, B.; Meyer, G.; Bohnert, H.J.; Schmitt, J.M. Age-dependent induction of pyruvate, orthophosphate dikinase in Mesembryanthemum crystallinum L. Planta 1995, 196, 492–500. [Google Scholar]
- Forsthoefel, N.R.; Cushman, M.A.F.; Cushman, J.C. Posttranscriptional and posttranslational control of enolase expression in the facultative crassulacean acid metabolism plant Mesembryanthemum crystallinum L. Plant Physiol. 1995, 108, 1185–1195. [Google Scholar] [CrossRef] [Green Version]
- Forsthoefel, N.R.; Vernon, D.M.; Cushman, J.C. A salinity-induced gene from the halophyte M. crystallinum encodes a glycolytic enzyme, cofactor-independent phosphoglyceromutase. Plant Mol. Biol. 1995, 29, 213–226. [Google Scholar] [CrossRef]
- Ostrem, J.A.; Vernon, D.M.; Bohnert, H.J. Increased expression of a gene coding for NAD: Glyceraldehyde-3-phosphate dehydrogenase during the transition from C3 photosynthesis to crassulacean acid metabolism in Mesembryanthemum crystallinum. J. Biol. Chem. 1990, 265, 3497–3502. [Google Scholar] [CrossRef]
- Cushman, J.C. Characterization and expression of a NADP-malic enzyme cDNA induced by salt stress from the facultative CAM plant, Mesembryanthemum crystallinum. Eur. J. Biochem. 1992, 208, 259–266. [Google Scholar] [CrossRef]
- Ocheretina, O.; Scheibe, R. Cloning and sequence analysis of cDNAs encoding cytosolic malate dehydrogenase. Gene 1997, 199, 145–148. [Google Scholar] [CrossRef]
- Löw, R.; Rockel, B.; Kirsch, M.; Ratajczak, R.; Hörntensteiner, S. Early salt stress effects on the differential expression of vacuolar H+-ATPase genes in roots and leaves of Mesembryanthemum crystallinum. Plant Physiol. 1996, 110, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Bartholomew, D.M.; Rees, D.J.G.; Rambaut, A.; Smith, J.A.C. Isolation and sequence analysis of a cDNA encoding the c subunit of a vacuolar-type H+-ATPase from the CAM plant Kalanchoë diagremontiana. Plant Mol. Biol. 1996, 31, 435–442. [Google Scholar] [CrossRef]
- Tsiantis, M.S.; Bartholomew, D.M.; Smith, J.A.C. Salt regulation of transcript levels for the c subunit of a leaf vacuolar H+-ATPase in the halophyte Mesembryanthemum crystallinum. Plant J. 1996, 9, 729–736. [Google Scholar] [CrossRef]
- Dietz, K.J.; Arbinger, B. cDNA sequence and expression of subunit E of the vacuolar H(+)-ATPase in the inducible crassulacean acid metabolism plant Mesembryanthemum crystallinum. Biochem. Biophys. Acta Gene Struct. Funct. 1996, 1281, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Baur, B.; Fisher, K.; Winter, K.; Dietz, K.J. cDNA sequences of a protein kinase from the halophyte Mesembryanthemum crystallinum L., encoding a SNF-1 homologue. Plant Physiol. 1994, 106, 1225–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breiteneder, H.; Michalowski, C.B.; Bohnert, H.J. Environmental stress mediated differential 3′ end formation of chloroplast RNA-binding protein transcripts. Plant Mol. Biol. 1994, 26, 833–849. [Google Scholar] [CrossRef]
- Rippmann, J.F.; Michalowski, C.B.; Nelson, D.E.; Bohnert, H.J. Induction of a ribosome-inactivating protein upon environmental stress. Plant Mol. Biol. 1997, 35, 701–709. [Google Scholar] [CrossRef]
- Cushman, J.C.; Michalowski, C.B.; Bohnert, H.J. Developmental control of crassulacean acid metabolism inducibility by salt stress in the common ice plant. Plant Physiol. 1990, 94, 1137–1142. [Google Scholar] [CrossRef] [Green Version]
- Cushman, J.C.; Bohnert, H.J. Transcriptional activation of CAM genes during development and environmental stress. In Ecological Studies; Smith, J.A.C., Winter, A.C., Eds.; Metabolism: Biochemistry, Ecophysiology and Evolution; Springer: Berlin, Germany, 1996; pp. 135–158. [Google Scholar]
- Bohnert, H.J.; Ayoubi, P.; Borchert, C.; Bressan, R.A.; Burnap, R.L.; Cushman, J.C.; Cushman, M.A.; Deyholos, M.; Fischer, R.; Galbraith, D.W.; et al. A genomics approach towards salt stress tolerance. Plant Physiol. Biochem. 2001, 39, 295–311. [Google Scholar] [CrossRef]
- Cushman, J.C.; Tillett, R.L.; Wood, J.A.; Branco, J.M.; Schlauch, K.A. Large-scale mRNA expression profiling in the common ice plant, Mesembryanthemum crystallinum, performing C3 photosynthesis and crassulacean acid metabolism (CAM). J. Exp. Bot. 2008, 59, 1875–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boxall, S.F.; Dever, L.V.; Kneřová, J.; Gould, P.D.; Hartwell, J. Phosphorylation of phosphoenolpyruvate ccarboxylase is essential for maximal and sustained dark CO2 Fixation and core circadian clock operation in the obligate crassulacean acid metabolism species Kalanchoë fedtschenkoi. Plant Cell 2017, 29, 2519–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wai, C.M.; Weise, S.E.; Ozersky, P.; Mockler, T.C.; Michael, T.P.; VanBuren, R. Time of day and network reprogramming during drought induced CAM photosynthesis in Sedum album. PLoS Genet. 2019, 15, e1008209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, H.; Zhang, L.S.; Zhang, G.Q.; Zheng, B.Q.; Liu, Z.J.; Wang, Y. Evolutionary history of PEPC genes in green plants: Implications for the evolution of CAM in orchids. Mol. Phylogenetics Evol. 2016, 94, 559–564. [Google Scholar] [CrossRef]
- Fankhauser, N.; Aubry, S. Post-transcriptional regulation of photosynthetic genes is a key driver of C4 leaf ontogeny. J. Exp. Bot. 2017, 68, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.Q.; Li, N.; Wang, Y.J.; Zhou, R.; Sun, P.C.; Lin, H.; Chen, W.; Yu, T.; Liu, Z.; Wang, Z.Y.; et al. High quality ice plant reference genome analysis provides insights into genome evolution and allows exploration of genes involved in the transition from C3 to CAM pathways. Plant Biotechnol. J. 2022, 20, 2107–2122. [Google Scholar] [CrossRef] [PubMed]
- Wai, C.M.; VanBuren, R.; Zhang, J.S.; Huang, L.X.; Miao, W.; Edger, P.P.; Yim, W.C.; Priest, H.D.; Meyers, B.C.; Mockler, T.; et al. Temporal and spatial transcriptomic and microRNA dynamics of CAM photosynthesis in pineapple. Plant J. Cell Mol. Biol. 2017, 92, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, C.P.; Yim, W.C.; Sun, Y.H.; Ohnishi, M.; Mimura, T.; Cushman, J.C.; Yen, H.E. Identification of ice plant (Mesembryanthemum crystallinum L.) microRNAs using RNA-seq and their putative roles in high salinity responses in seedlings. Front. Plant Sci. 2016, 7, 1143. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.K.; Nie, Z.Y.; Yan, C.; Huang, H.; Ma, X.J.; Wang, Y.P.; Ye, N.; Tuskan, G.A.; Yang, X.H.; Yin, H.F. Transcriptome and degradome profiling reveals a role of miR530 in the circadian regulation of gene expression in Kalanchoë marnieriana. Cells 2021, 10, 1526. [Google Scholar] [CrossRef] [PubMed]
- Kore-eda, S.; Cushman, M.A.; Akselrod, I.; Bufford, D.; Fredrickson, M.; Clark, E.; Cushman, J.C. Transcript profiling of salinity stress responses by large-scale expressed sequence tag analysis in Mesembryanthemum crystallinum. Gene 2004, 341, 83–92. [Google Scholar] [CrossRef]
- Guan, Q.J.; Kong, W.W.; Zhu, D.; Zhu, W.; Dufresne, C.; Tian, J.K.; Chen, S.X. Comparative proteomics of Mesembryanthemum crystallinum guard cells and mesophyll cells in transition from C3 to CAM. J. Proteom. 2021, 231, 104019. [Google Scholar] [CrossRef]
- Perron, N.; Tan, B.; Dufresne, C.P.; Chen, S.X. Proteomics and phosphoproteomics of C3 to CAM transition in the common ice plant. Methods Enzymol. 2022, 676, 347–368. [Google Scholar]
- Nobe, P. High productivities of certain agronomic CAM species. Crassulacean Acid Metabolism. In Biochemistry, Ecophysiology and Evolution; Springer: Berlin/Heidelberg, Germany, 1996; pp. 255–265. [Google Scholar]
- Töpfer, N.; Braam, T.; Shameer, S.; Ratcliffe, R.G.; Sweetlove, L.J. Alternative crassulacean acid metabolism modes provide environment-specific water-saving benefits in a leaf metabolic model. Plant Cell 2020, 32, 3689–3705. [Google Scholar] [CrossRef]
- Winter, K.; Smith, J.A.C. (Eds.) Taxonomic distribution of crassulacean acid metabolism. In Crassulacean Acid Metabolism. Biochemistry, Ecophysiology and Evolution; Springer: Berlin/Heidelberg, Germany, 1996; pp. 427–436. [Google Scholar]
- Qiu, S.; Sultana, S.; Liu, Z.D.; Yin, L.Y.; Wang, C.Y. Identification of obligate C3 photosynthesis in Dendrobium. Photosynthetica 2015, 53, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.H.; Wan, X.; Deng, H.; Zheng, B.Q.; Li, B.J.; Wang, Y. RNA-seq transcriptomic profiling of crassulacean acid metabolism pathway in Dendrobium catenatum. Sci. Data 2018, 5, 180252. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.J.; He, D.X.; Gao, R.F. Concomitant CAM and C3 photosynthetic pathways in Dendrobium officinale plants. J. Am. Soc. Hortic. Sci. 2014, 139, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Winter, K.; Holtum, J.A.M. Cryptic crassulacean acid metabolism (CAM) in Jatropha curcas. Funct Plant Biol. 2015, 42, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.L.; Hassan, M.M.; Liu, D.G.; Lim, S.D.; Yim, W.C.; Cushman, J.C.; Markel, K.; Shih, P.M.; Lu, H.W.; Weston, D.J.; et al. Biosystems Design to Accelerate C3-to-CAM Progression. BioDesign Res. 2020, 2020, 3686791. [Google Scholar] [CrossRef]
- Cosentino, C.; Silvestre, D.D.; Fischer-Schliebs, E.; Homann, U.; Palma, A.D.; Comunian, C.; Mauri, P.L.; Thiel, G. Proteomic analysis of Mesembryanthemum crystallinum leaf microsomal fractions finds an imbalance in V-ATPase stoichiometry during the salt-induced transition from C3 to CAM. Biochem. J. 2013, 450, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, D.H.; Barkla, B.J.; Vera-Estrella, R.; Pantoja, O.; Lee, S.Y.; Bohnert, H.J.; Dassanayake, M. Cell type-specific responses to Psalinity–the epidermal bladder cell transcriptome of Mesembryanthemum crystallinum. New Phytol. 2015, 207, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Barkla, B.J.; Vera-Estrella, R. Single cell-type comparative metabolomics of epidermal bladder cells from the halophyte Mesembryanthemum crystallinum. Front. Plant Sci. 2015, 6, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukagoshi, H.; Suzuki, T.; Nishikawa, K.; Agarie, S.; Ishiguro, S.; Higashiyama, T. RNA-seq analysis of the response of the halophyte, Mesembryanthemum crystallinum (ice plant) to high salinity. PLoS ONE 2015, 10, e0118339. [Google Scholar] [CrossRef]
- Barkla, B.J.; Vera-Estrella, R.; Raymond, C. Single-cell-type quantitative proteomic and ionomic analysis of epidermal bladder cells from the halophyte model plant Mesembryanthemum crystallinum to identify salt-responsive proteins. BMC Plant. Biol. 2016, 16, 110. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Chang, Y.; Zhao, W.J.; Qian, C.J.; Yin, X.Y.; Fan, X.K.; Zhu, X.Y.; Zhao, X.Q.; Ma, X.F. Transcriptome profiling reveals that foliar water uptake occurs with C3 and crassulacean acid metabolism facultative photosynthesis in Tamarix ramosissima under extreme drought. AoB Plants 2022, 14, plab060. [Google Scholar] [CrossRef] [PubMed]
- Heyduk, K.; McAssey, E.V.; Leebens-Mack, J. Differential timing of gene expression and recruitment in independent origins of CAM in the Agavoideae (Asparagaceae). New Phytol. 2022, 235, 2111–2126. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Physiological and Metabolic shift of the facultative CAM plants from C3 to CAM photosynthesis. For the facultative CAM plants, C3 state of photosynthesis is used to fix CO2 under well conditions, when stomata open during the day and close at night. Once the facultative CAM plants stressed by abiotic stress (such as atmospheric CO2 concentration, drought, salinity, photoperiod, light, etc.), the carbon assimilation pathway will be induced to the CAM mode, when CO2 uptakes during the nighttime via open stomata and stores as organic acids (mainly malic acid). Then, organic acids are decarboxylated in the vacuoles during the daytime and CO2 is refixed via Calvin cycle. CA, Carbonic anhydrase; PEP, Phosphoenol pyruvate; PEPC, Phosphoenolpyruvate carboxylase; PPCK, PEPC kinase; PPDK, Pyruvate phosphate kinase; ME, Malic enzyme; MDH, Malic dehydrogenase; OAA, Oxalacetic acid; RUBP, Ribulose-1,5-bisphosphate carboxylase; NAD, Nicotinamide adenine denucleotide; NADP, Nicotinamide adenine denucleotide phosphate.
Figure 1.
Physiological and Metabolic shift of the facultative CAM plants from C3 to CAM photosynthesis. For the facultative CAM plants, C3 state of photosynthesis is used to fix CO2 under well conditions, when stomata open during the day and close at night. Once the facultative CAM plants stressed by abiotic stress (such as atmospheric CO2 concentration, drought, salinity, photoperiod, light, etc.), the carbon assimilation pathway will be induced to the CAM mode, when CO2 uptakes during the nighttime via open stomata and stores as organic acids (mainly malic acid). Then, organic acids are decarboxylated in the vacuoles during the daytime and CO2 is refixed via Calvin cycle. CA, Carbonic anhydrase; PEP, Phosphoenol pyruvate; PEPC, Phosphoenolpyruvate carboxylase; PPCK, PEPC kinase; PPDK, Pyruvate phosphate kinase; ME, Malic enzyme; MDH, Malic dehydrogenase; OAA, Oxalacetic acid; RUBP, Ribulose-1,5-bisphosphate carboxylase; NAD, Nicotinamide adenine denucleotide; NADP, Nicotinamide adenine denucleotide phosphate.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qiu, S.; Xia, K.; Yang, Y.; Wu, Q.; Zhao, Z. Mechanisms Underlying the C3–CAM Photosynthetic Shift in Facultative CAM Plants. Horticulturae 2023, 9, 398. https://doi.org/10.3390/horticulturae9030398
AMA Style
Qiu S, Xia K, Yang Y, Wu Q, Zhao Z. Mechanisms Underlying the C3–CAM Photosynthetic Shift in Facultative CAM Plants. Horticulturae. 2023; 9(3):398. https://doi.org/10.3390/horticulturae9030398
Chicago/Turabian StyleQiu, Shuo, Ke Xia, Yanni Yang, Qiaofen Wu, and Zhiguo Zhao. 2023. "Mechanisms Underlying the C3–CAM Photosynthetic Shift in Facultative CAM Plants" Horticulturae 9, no. 3: 398. https://doi.org/10.3390/horticulturae9030398
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.