
Characterization, Evolutionary Analysis, and Expression Pattern Analysis of the Heat Shock Transcription Factors and Drought Stress Response in Heimia myrtifolia
Abstract
:1. Introduction
2. Materials and Methods
2.1. HSF Gene Characterization and Sequencing Analysis in H. myrtifolia
2.2. Prediction of the Physicochemical Properties and Protein Structure of HmHSF
2.3. Phylogenetic Tree Construction and Sequence Comparison
2.4. GO Database Annotation and Enrichment Analysis
2.5. Plant Material, Methods, and qRT-PCR Validation
3. Results
3.1. Identification of HSF Genes in H. myrtifolia
3.2. HmHSF Protein Sequence Analysis
3.3. Phylogenetic Analysis of HmHSF Protein
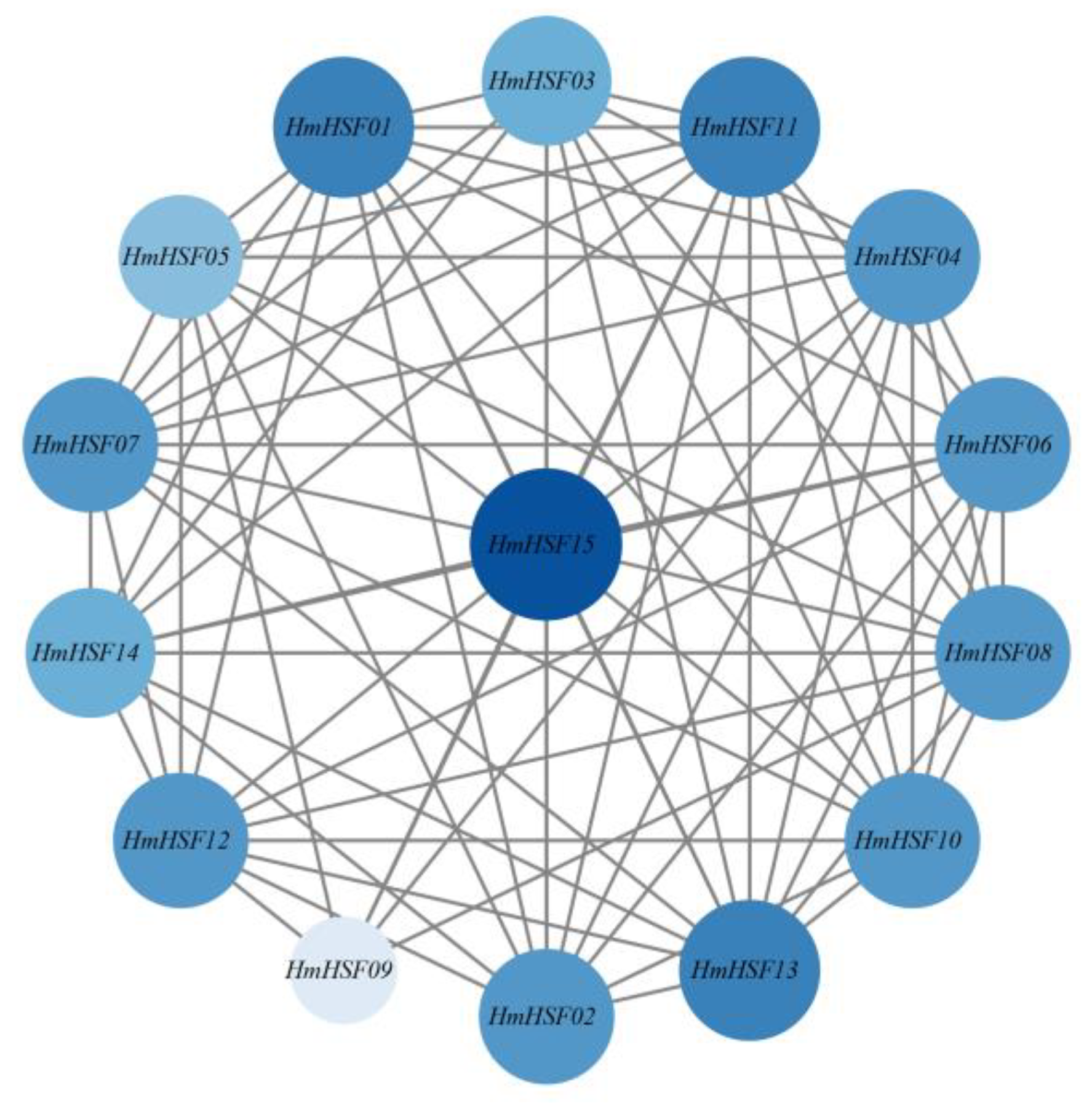
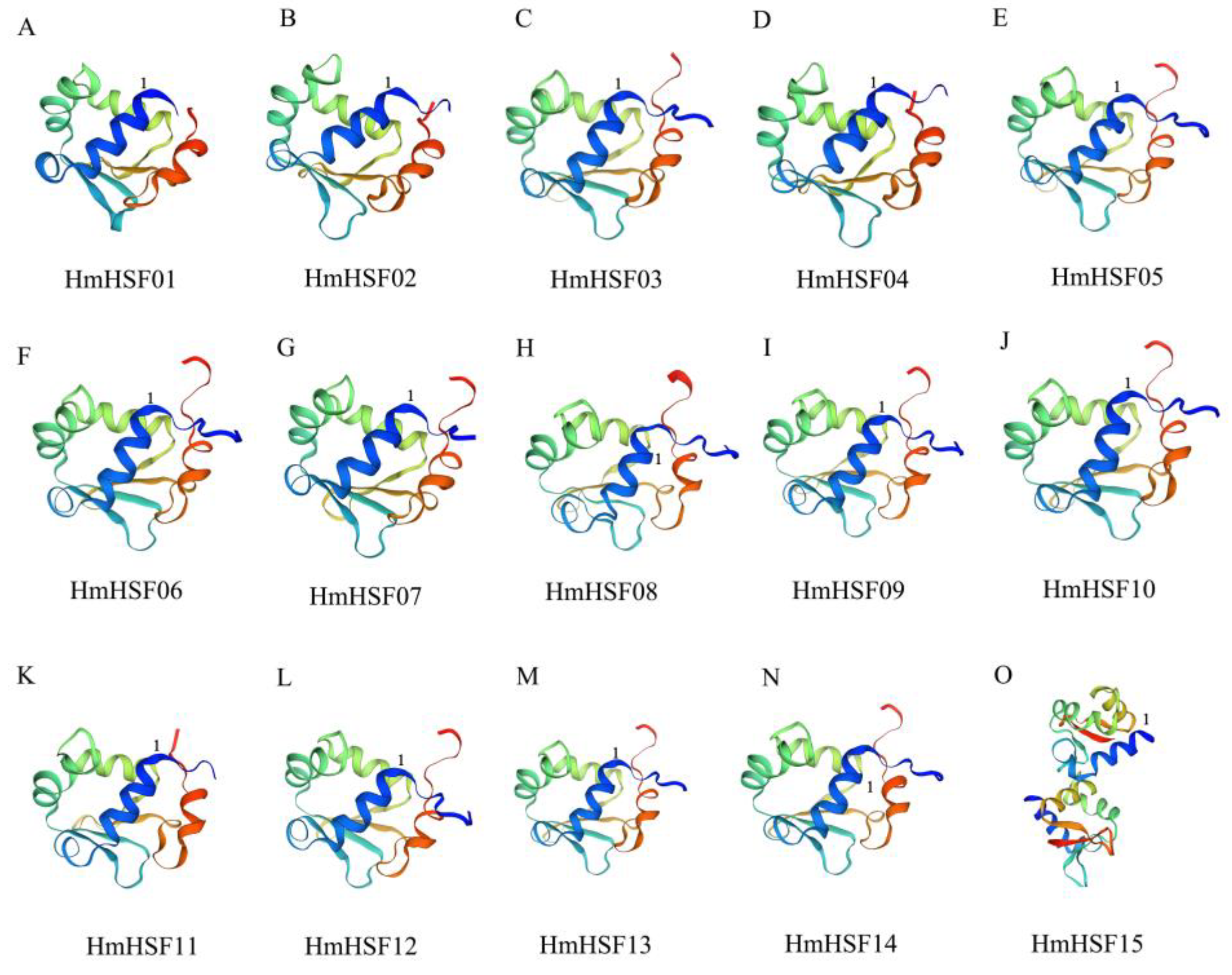
3.4. Structural and Interaction Network Analysis of the HmHSF Protein
3.5. Analysis of the GO Annotation and Enrichment
3.6. Expression and K-Means Clustering Analysis of HmHSF Gene
3.7. Expression of HSF Genes under Drought Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription Factors and Plants Response to Drought Stress: Current Understanding and Future Directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, M.; Nisar, M.; Khan, N.; Hazrat, A.; Khan, A.H.; Hayat, K.; Fahad, S.; Khan, A.; Ullah, A. Drought Tolerance Strategies in Plants: A Mechanistic Approach. J. Plant Growth Regul. 2021, 40, 926–944. [Google Scholar] [CrossRef]
- Batool, M.; El-Badri, A.M.; Hassan, M.U.; Haiyun, Y.; Chunyun, W.; Zhenkun, Y.; Jie, K.; Wang, B.; Zhou, G. Drought Stress in Brassica napus: Effects, Tolerance Mechanisms, and Management Strategies. J. Plant Growth Regul. 2022, 42, 21–45. [Google Scholar] [CrossRef]
- Zhang, X.; Lei, L.; Lai, J.; Zhao, H.; Song, W. Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings. BMC Plant Biol. 2018, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crop. Prod. 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Leng, P.; Zhao, J. Transcription factors as molecular switches to regulate drought adaptation in maize. Theor. Appl. Genet. 2020, 133, 1455–1465. [Google Scholar] [CrossRef]
- Zhang, D.-F.; Zeng, T.-R.; Liu, X.-Y.; Gao, C.-X.; Li, Y.-X.; Li, C.-H.; Song, Y.-C.; Shi, Y.-S.; Wang, T.-Y.; Li, Y. Transcriptomic profiling of sorghum leaves and roots responsive to drought stress at the seedling stage. J. Integr. Agr. 2019, 18, 1980–1995. [Google Scholar] [CrossRef]
- Yao, T.; Zhang, J.; Xie, M.; Yuan, G.; Tschaplinski, T.J.; Muchero, W.; Chen, J.-G. Transcriptional Regulation of Drought Response in Arabidopsis and Woody Plants. Fron. Plant Sci. 2021, 11, 572137. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, J.; Zheng, J.; Gong, Z.; Li, Z.; Ai, X.; Li, X.; Chen, Q. Genome-Wide Comparative Analysis of Heat Shock Transcription Factors Provides Novel Insights for Evolutionary History and Expression Characterization in Cotton Diploid and Tetraploid Genomes. Front. Genet. 2021, 12, 658847. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zheng, S.; Liu, R.; Lu, J.; Lu, L.; Zhang, C.; Liu, Z.; Luo, C.; Zhang, L.; Yant, L.; et al. Genome-wide identification, phylogenetic and expression analysis of the heat shock transcription factor family in bread wheat (Triticum aestivum L.). BMC Genomics. 2019, 20, 505. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yu, M.; Zhang, S.; Song, T.; Zhang, M.; Zhou, H.; Wang, Y.; Xiang, J.; Zhang, X. Transcriptomic Identification of Wheat AP2/ERF Transcription Factors and Functional Characterization of TaERF-6-3A in Response to Drought and Salinity Stresses. Int. J. Mol. Sci. 2022, 23, 3272. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wan, X.-L.; Yu, J.-Y.; Wang, K.-L.; Zhang, J. Genome-Wide Identification, Classification, and Expression Analysis of the Hsf Gene Family in Carnation (Dianthus caryophyllus). Int. J. Mol. Sci. 2019, 20, 5233. [Google Scholar] [CrossRef]
- Liu, X.; Meng, P.; Yang, G.; Zhang, M.; Peng, S.; Zhai, M.Z. Genome-wide identification and transcript profiles of walnut heat stress transcription factor involved in abiotic stress. BMC Genom. 2020, 21, 474. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Tang, R.; Wang, L.; Chen, C.; Ren, Z. Genome-wide identification and expression analysis of Hsf and Hsp gene families in cucumber (Cucumis sativus L.). Plant Growth Regul. 2021, 95, 223–239. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, C.; Wang, C.; Yun, L.; Song, L.; Idrees, M.; Liu, H.; Zhang, Q.; Yang, J.; Zheng, X.; et al. OsHsfB4b Confers Enhanced Drought Tolerance in Transgenic Arabidopsis and Rice. Int. J. Mol. Sci. 2022, 23, 10830. [Google Scholar] [CrossRef]
- Song, X.; Liu, G.; Duan, W.; Liu, T.; Huang, Z.; Ren, J.; Li, Y.; Hou, X. Genome-wide identification, classification and expression analysis of the heat shock transcription factor family in Chinese cabbage. Mol. Genet. Genom. 2014, 289, 541–551. [Google Scholar] [CrossRef]
- Lal, S.V.; Brahma, B.; Gohain, M.; Mohanta, D.; De, B.C.; Chopra, M.; Dass, G.; Vats, A.; Upadhyay, R.C.; Datta, T.K.; et al. Splice variants and seasonal expression of buffalo HSF genes. Cell Stress Chaperones 2015, 20, 545–554. [Google Scholar] [CrossRef]
- Chen, J.; Gao, T.; Wan, S.; Zhang, Y.; Yang, J.; Yu, Y.; Wang, W. Genome-Wide Identification, Classification and Expression Analysis of the HSP Gene Superfamily in Tea Plant (Camellia sinensis). Int. J. Mol. Sci. 2018, 19, 2633. [Google Scholar] [CrossRef]
- Jiang, L.; Hu, W.; Qian, Y.; Ren, Q.; Zhang, J. Genome-wide identification, classification and expression analysis of the Hsf and Hsp70 gene families in maize. Gene 2021, 770, 145348. [Google Scholar] [CrossRef] [PubMed]
- Andrási, N.; Pettkó-Szandtner, A.; Szabados, L. Diversity of plant heat shock factors: Regulation, interactions, and functions. J. Exp. Bot. 2020, 72, 1558–1575. [Google Scholar] [CrossRef] [PubMed]
- Jahan, K.; Nie, H.; Yan, X. Revealing the potential regulatory relationship between HSP70, HSP90 and HSF genes under temperature stress. Fish Shellfish Immunol. 2023, 134, 108607. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Wang, J.; Wang, Q.; Ji, M.; Hong, S.; Shang, L.; Zhang, G.; Zhao, Y.; Ma, Q.; Gu, C. Transcriptome Approach Reveals the Response Mechanism of Heimia myrtifolia (Lythraceae, Myrtales) to Drought Stress. Fron. Plant Sci. 2022, 13, 877913. [Google Scholar] [CrossRef]
- Giorno, F.; Guerriero, G.; Baric, S.; Mariani, C. Heat shock transcriptional factors in Malus domestica: Identification, classification and expression analysis. BMC Genom. 2012, 13, 639. [Google Scholar] [CrossRef]
- Huang, B.; Huang, Z.; Ma, R.; Chen, J.; Zhang, Z.; Yrjälä, K. Genome-wide identification and analysis of the heat shock transcription factor family in moso bamboo (Phyllostachys edulis). Sci. Rep. 2021, 11, 16492. [Google Scholar] [CrossRef]
- Zhang, H.; Li, G.; Fu, C.; Duan, S.; Hu, D.; Guo, X. Genome-wide identification, transcriptome analysis and alternative splicing events of Hsf family genes in maize. Sci. Rep. 2020, 10, 8073. [Google Scholar] [CrossRef]
- Liu, G.; Chai, F.; Wang, Y.; Jiang, J.; Duan, W.; Wang, Y.; Wang, F.; Li, S.; Wang, L. Genome-wide Identification and Classification of HSF Family in Grape, and Their Transcriptional Analysis under Heat Acclimation and Heat Stress. Hortic. Plant J. 2018, 4, 133–143. [Google Scholar] [CrossRef]
- MMallick, B.; Kumari, M.; Pradhan, S.K.; Acharya, G.C.; Naresh, P.; Das, B.; Shashankar, P. Genome-wide analysis and characterization of heat shock transcription factors (Hsfs) in common bean (Phaseolus vulgaris L.). Funct. Integr. Genom. 2022, 22, 743–756. [Google Scholar] [CrossRef]
- Huang, Y.; Li, M.-Y.; Wang, F.; Xu, Z.-S.; Huang, W.; Wang, G.-L.; Ma, J.; Xiong, A.-S. Heat shock factors in carrot: Genome-wide identification, classification, and expression profiles response to abiotic stress. Mol. Biol. Rep. 2015, 42, 893–905. [Google Scholar] [CrossRef]
- Liu, Z.-W.; Wu, Z.-J.; Li, X.-H.; Huang, Y.; Li, H.; Wang, Y.-X.; Zhuang, J. Identification, classification, and expression profiles of heat shock transcription factors in tea plant (Camellia sinensis) under temperature stress. Gene 2016, 576, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Chidambaranathan, P.; Jagannadham, P.T.K.; Satheesh, V.; Kohli, D.; Basavarajappa, S.H.; Chellapilla, B.; Kumar, J.; Jain, P.K.; Srinivasan, R. Genome-wide analysis identifies chickpea (Cicer arietinum) heat stress transcription factors (Hsfs) responsive to heat stress at the pod development stage. J. Plant Res. 2018, 131, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.R.; Goswami, S.; Singh, K.; Dubey, K.; Rai, G.K.; Singh, B.; Singh, S.; Grover, M.; Mishra, D.; Kumar, S.; et al. Characterization of novel heat-responsive transcription factor (TaHSFA6e) gene involved in regulation of heat shock proteins (HSPs)—A key member of heat stress-tolerance network of wheat. J. Biotechnol. 2018, 279, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bin, J.; Zhu, M.; Ding, H.; Zai, Z.; Shi, T.; Wang, L.; Yang, X.; Yue, Y. New Insights into the Roles of Osmanthus Fragrans Heat-Shock Transcription Factors in Cold and Other Stress Responses. Horticulturae 2022, 8, 80. [Google Scholar] [CrossRef]
- Xu, P.; Guo, Q.; Pang, X.; Zhang, P.; Kong, D.; Liu, J. New Insights into Evolution of Plant Heat Shock Factors (Hsfs) and Expression Analysis of Tea Genes in Response to Abiotic Stresses. Plants 2020, 9, 311. [Google Scholar] [CrossRef]
- Shyamli, P.S.; Pradhan, S.; Panda, M.; Parida, A. De novo Whole-Genome Assembly of Moringa oleifera Helps Identify Genes Regulating Drought Stress Tolerance. Fron. Plant Sci. 2021, 12, 766999. [Google Scholar] [CrossRef]
- Li, P.-S.; Yu, T.-F.; He, G.-H.; Chen, M.; Zhou, Y.-B.; Chai, S.-C.; Xu, Z.-S.; Ma, Y.-Z. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses. BMC Genom. 2014, 15, 1009. [Google Scholar] [CrossRef]
- Ma, J.; Xu, Z.-S.; Wang, F.; Tan, G.-F.; Li, M.-Y.; Xiong, A.-S. Genome-wide analysis of HSF family transcription factors and their responses to abiotic stresses in two Chinese cabbage varieties. Acta Physiol. Plant. 2014, 36, 513–523. [Google Scholar] [CrossRef]
- Ma, G.; Shen, J.; Yu, H.; Huang, X.; Deng, X.; Hu, Z.; Amee, M.; Chen, L.; Cao, L. Genome-wide identification and functional analyses of heat shock transcription factors involved in heat and drought stresses in ryegrass. Environ. Exp. Bot. 2022, 201, 104968. [Google Scholar] [CrossRef]
- Liu, M.; Huang, Q.; Sun, W.; Ma, Z.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide investigation of the heat shock transcription factor (Hsf) gene family in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2019, 20, 871. [Google Scholar] [CrossRef]
- Gong, C.; Pang, Q.; Li, Z.; Li, Z.; Chen, R.; Sun, G.; Sun, B. Genome-Wide Identification and Characterization of Hsf and Hsp Gene Families and Gene Expression Analysis under Heat Stress in Eggplant (Solanum melongema L.). Horticulturae 2021, 7, 149. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Size (aa) | MW (kDa) | pI | Stability | A.I | GRAVY | Predicted Location |
|---|---|---|---|---|---|---|---|---|
| HmHSF01 | Gene.80304 | 446 | 50501.11 | 5.83 | U | 65.88 | −0.869 | Nuclear |
| HmHSF02 | Gene.33060 | 363 | 40958.06 | 4.81 | U | 81.87 | −0.515 | Nuclear |
| HmHSF03 | Gene.82041 | 290 | 32889.93 | 6.35 | U | 59.55 | −0.808 | Nuclear |
| HmHSF04 | Gene.67227 | 391 | 44075.58 | 5.11 | U | 61.10 | −0.931 | Nuclear |
| HmHSF05 | Gene.83400 | 339 | 38308.34 | 8.23 | U | 70.44 | −0.522 | Nuclear |
| HmHSF06 | Gene.78293 | 325 | 36158.11 | 4.64 | U | 65.38 | −0.677 | Nuclear |
| HmHSF07 | Gene.92484 | 372 | 41725.87 | 9.07 | U | 71.05 | −0.756 | Nuclear |
| HmHSF08 | Gene.77897 | 303 | 34137.49 | 6.19 | U | 68.58 | −0.549 | Nuclear |
| HmHSF09 | Gene.93333 | 253 | 28667.25 | 9.33 | U | 61.70 | −0.836 | Nuclear |
| HmHSF10 | Gene.82043 | 295 | 32617.38 | 8.31 | U | 66.88 | −0.69 | Nuclear |
| HmHSF11 | Gene.98726 | 427 | 48851.44 | 5.16 | U | 69.13 | −0.784 | Nuclear |
| HmHSF12 | Gene.96475 | 392 | 44531.09 | 6.04 | U | 77.83 | −0.663 | Nuclear |
| HmHSF13 | Gene.44218 | 320 | 34804.22 | 5.33 | U | 72.28 | −0.46 | Nuclear |
| HmHSF14 | Gene.51133 | 279 | 30884.61 | 7.02 | U | 62.26 | −0.785 | Nuclear |
| HmHSF15 | Gene.86874 | 492 | 53592.58 | 4.85 | U | 61.20 | −0.643 | Nuclear |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Gu, C.; Ye, Y.; Zhao, Y.; Shang, L.; Shao, W.; Hong, S.; Ma, J. Characterization, Evolutionary Analysis, and Expression Pattern Analysis of the Heat Shock Transcription Factors and Drought Stress Response in Heimia myrtifolia. Horticulturae 2023, 9, 588. https://doi.org/10.3390/horticulturae9050588
Zhang G, Gu C, Ye Y, Zhao Y, Shang L, Shao W, Hong S, Ma J. Characterization, Evolutionary Analysis, and Expression Pattern Analysis of the Heat Shock Transcription Factors and Drought Stress Response in Heimia myrtifolia. Horticulturae. 2023; 9(5):588. https://doi.org/10.3390/horticulturae9050588
Chicago/Turabian StyleZhang, Guozhe, Cuihua Gu, Yacheng Ye, Yu Zhao, Linxue Shang, Weili Shao, Sidan Hong, and Jin Ma. 2023. "Characterization, Evolutionary Analysis, and Expression Pattern Analysis of the Heat Shock Transcription Factors and Drought Stress Response in Heimia myrtifolia" Horticulturae 9, no. 5: 588. https://doi.org/10.3390/horticulturae9050588