
BcAMT1;5 Mediates Nitrogen Uptake and Assimilation in Flowering Chinese Cabbage and Improves Plant Growth When Overexpressed in Arabidopsis
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
2.2. Cloning of BcAMT1;5 and Bioinformatics Analysis
2.3. Sub-Cellular Location
2.4. qPCR
2.5. Functional Complementation in Yeast Mutant 31019b
2.6. Construction of pCAMBIA1391-BcAMT1;5pro::GUS for Arabidopsis Transformation and Glucorinidase (GUS) Assays
2.7. Construction of BcAMT1;5-Overexpressing Lines in Arabidopsis
2.8. Plant Culture for Growth Test, NH4+ Uptake, Ion Fluxes and Gene Expression
2.9. Statistical Analysis
3. Results
3.1. Molecular Identification of AMT1;5 Homolog from Flowering Chinese Cabbage
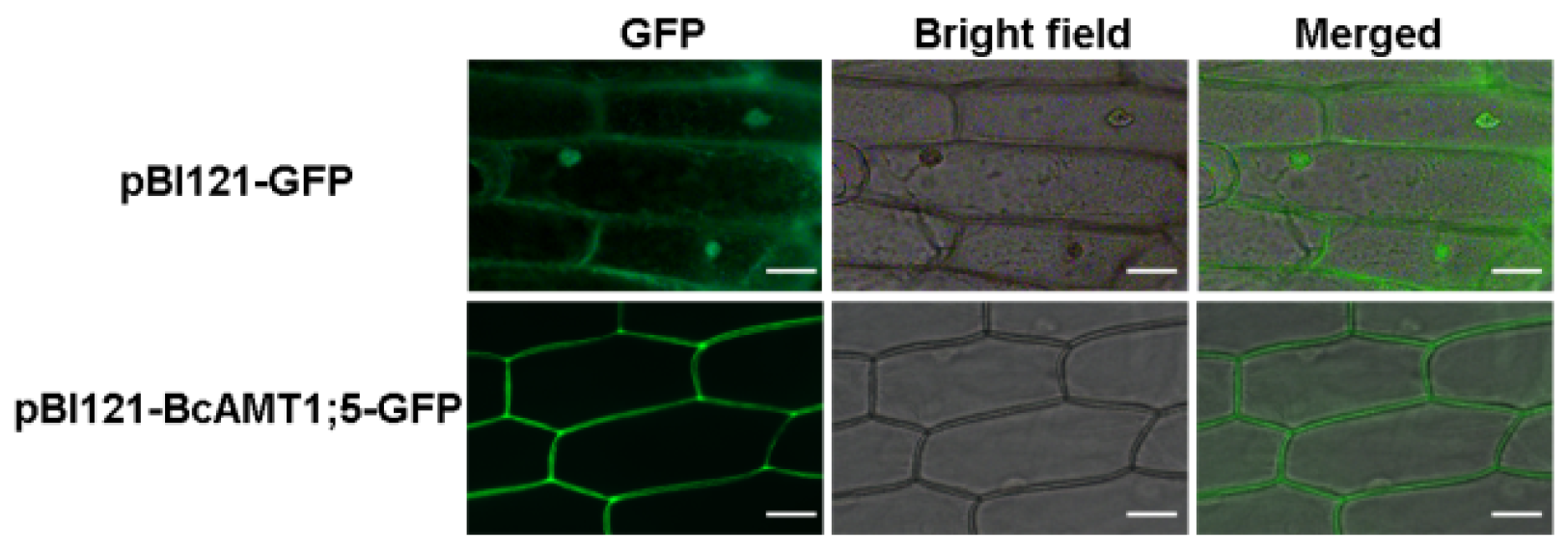
3.2. Subcellular Localization of BcAMT1;5 and its Functional Complementation Analysis in Yeast Mutant Cells
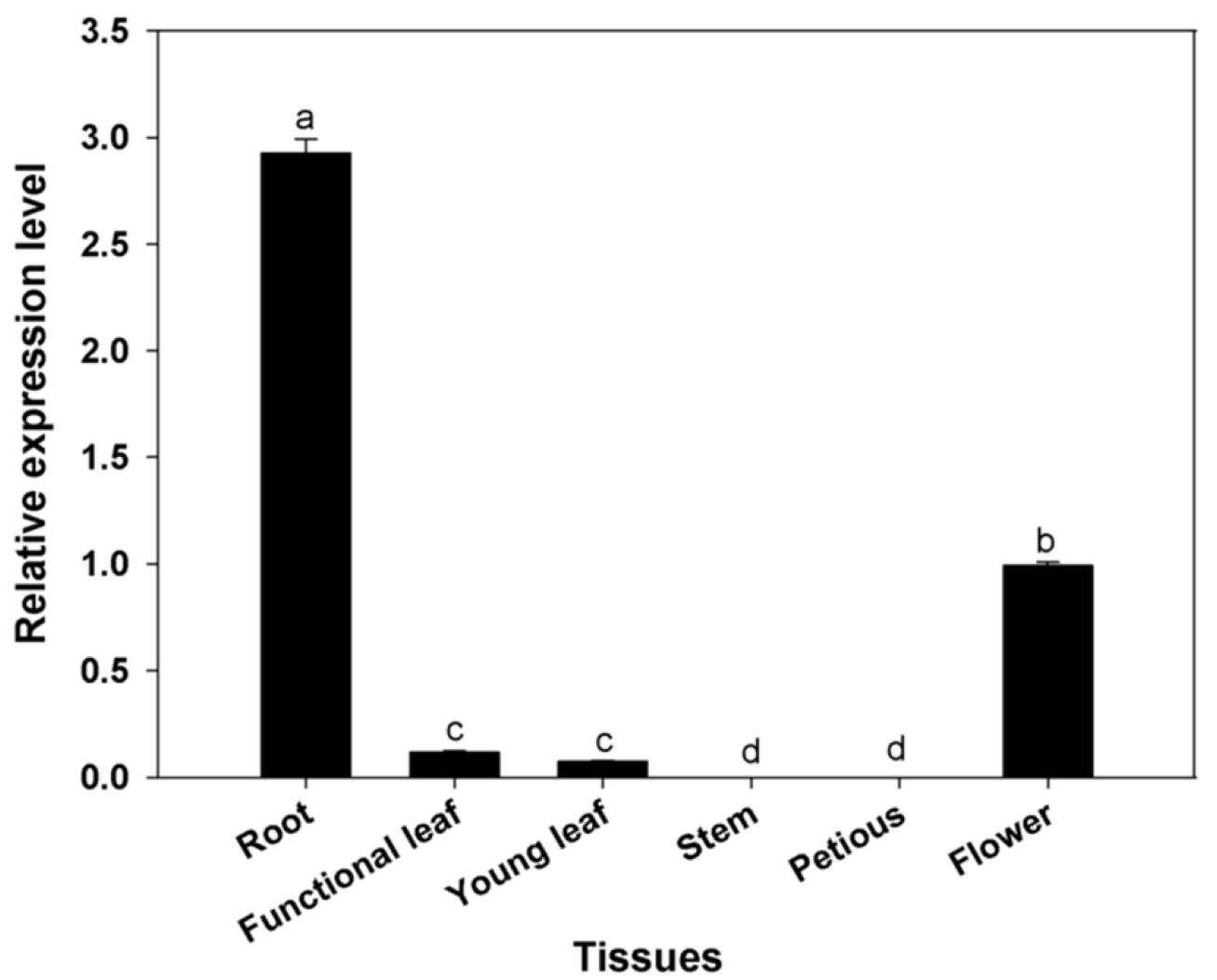
3.3. Expression Profiles for BcAMT1;5 Gene in Different Tissues of Flowering Chinese Cabbage
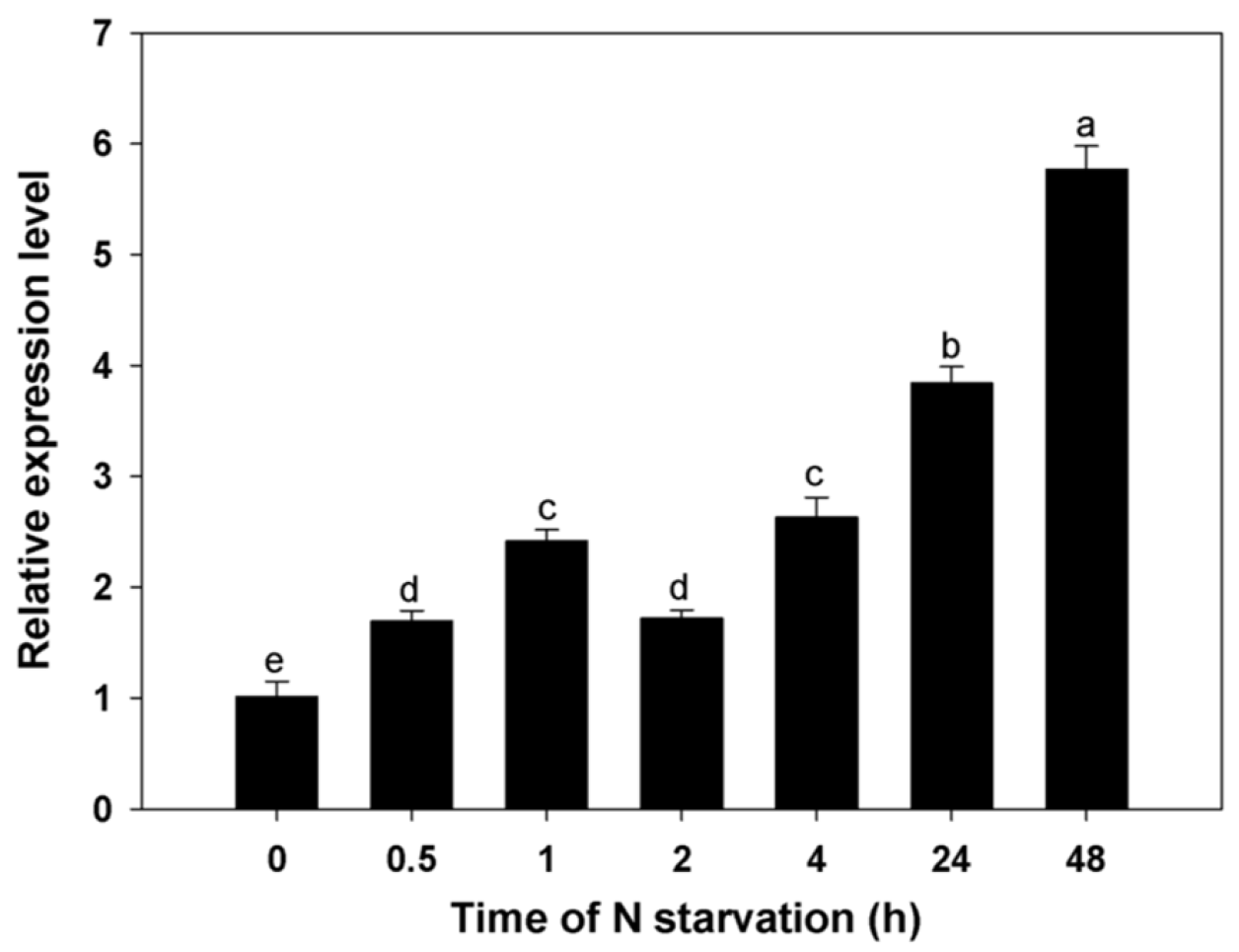
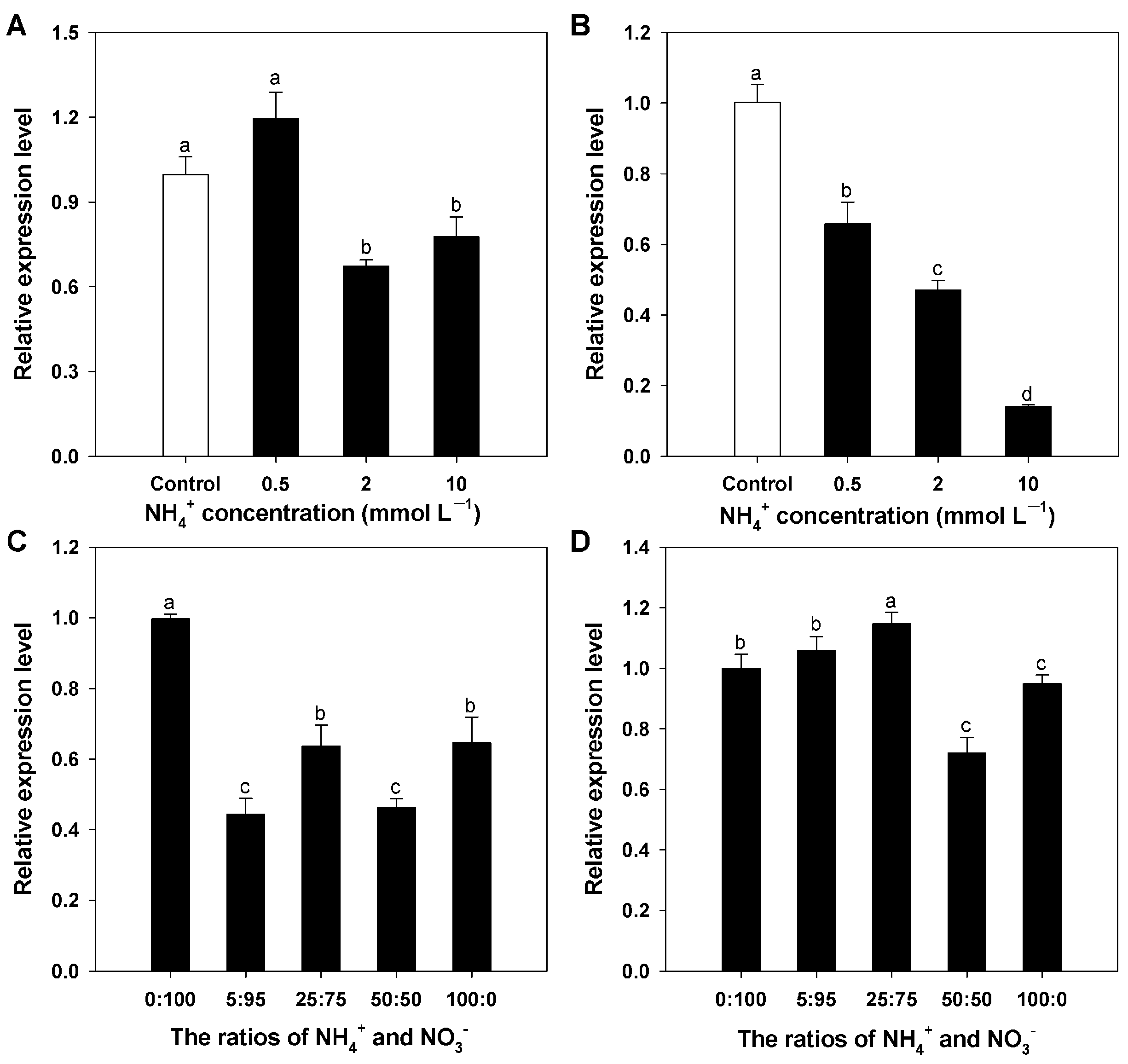
3.4. Expression Profiles for BcAMT1;5 Gene in Different N Regimes
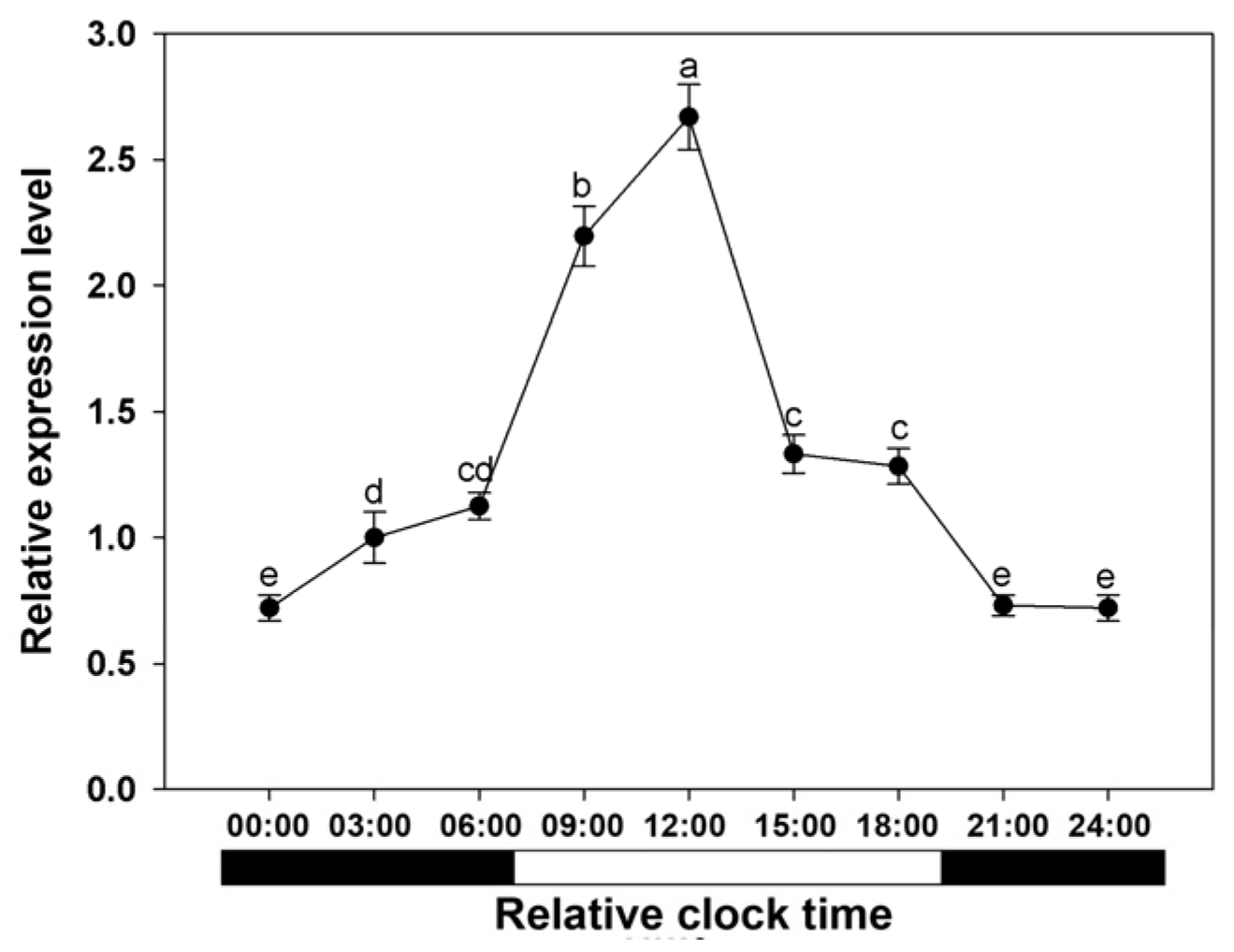
3.5. Expression of BcAMT1;5 Gene in Light Regimes
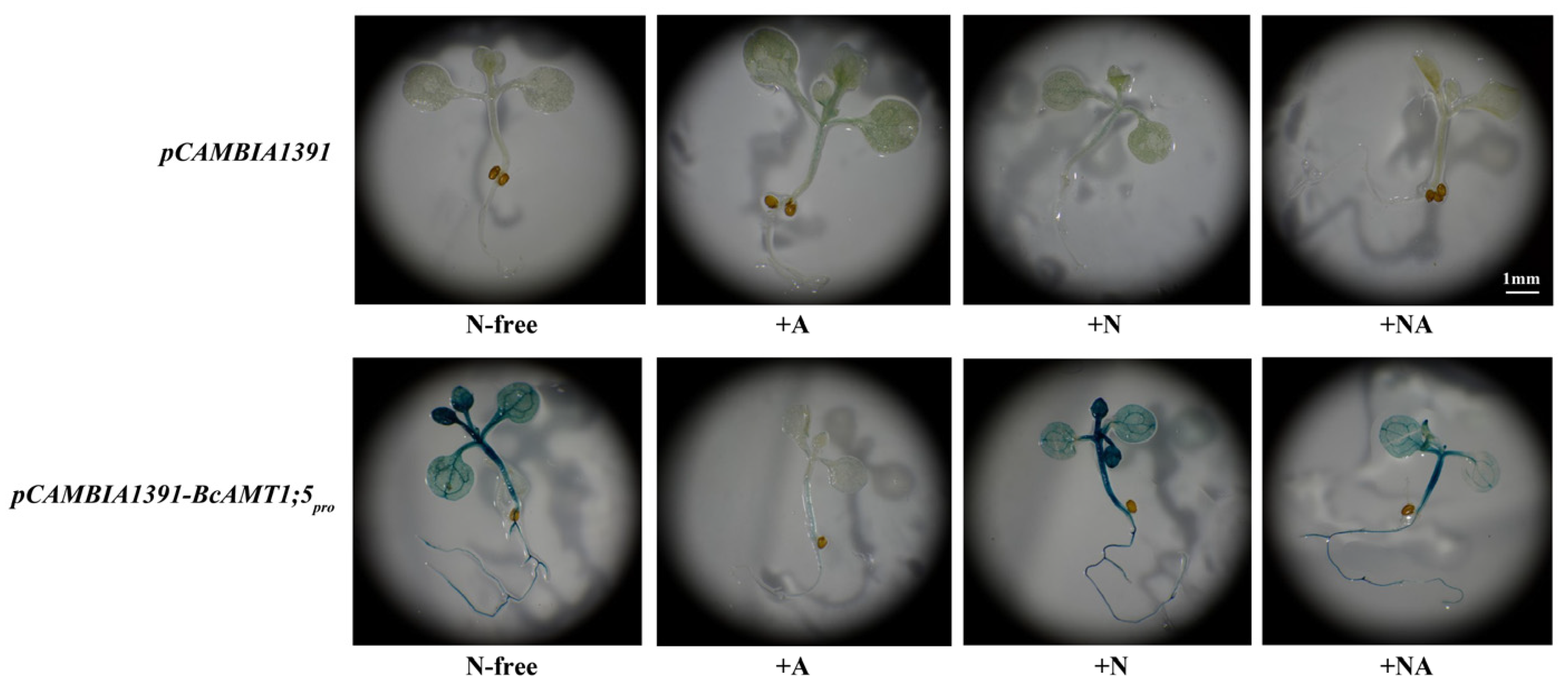
3.6. The GUS Activity of BcAMT1;5pro::GUS in Response to Different N Conditions
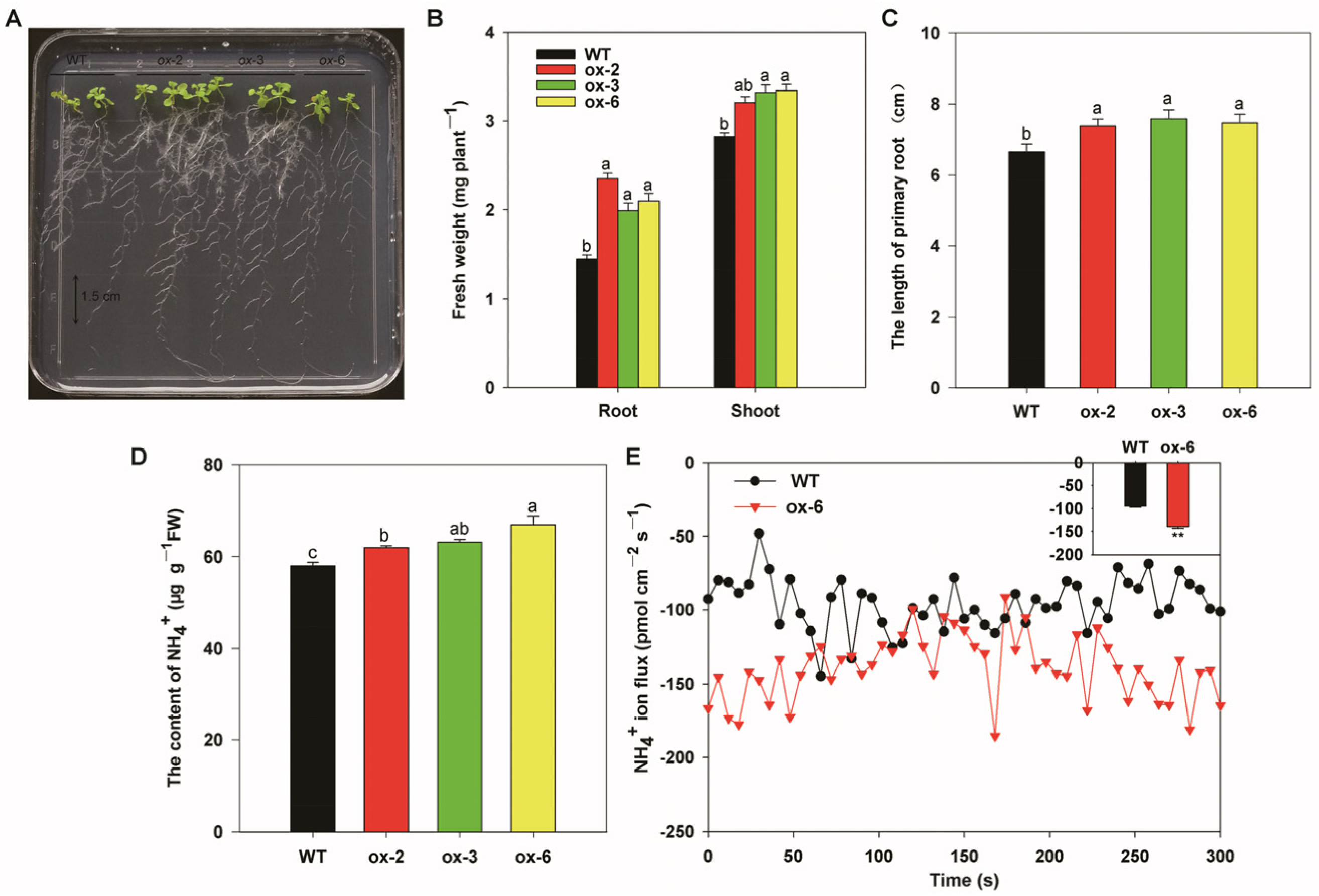
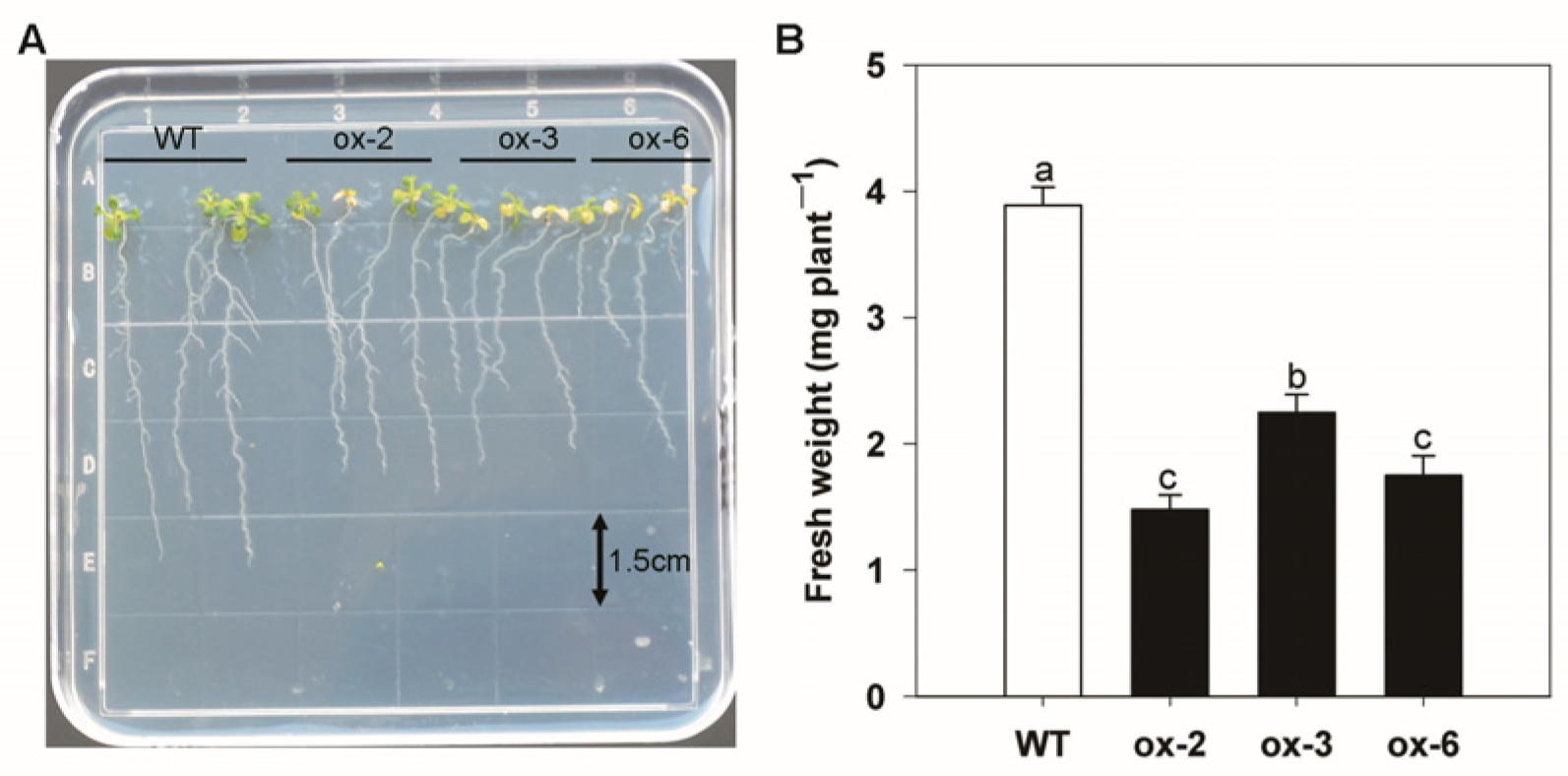
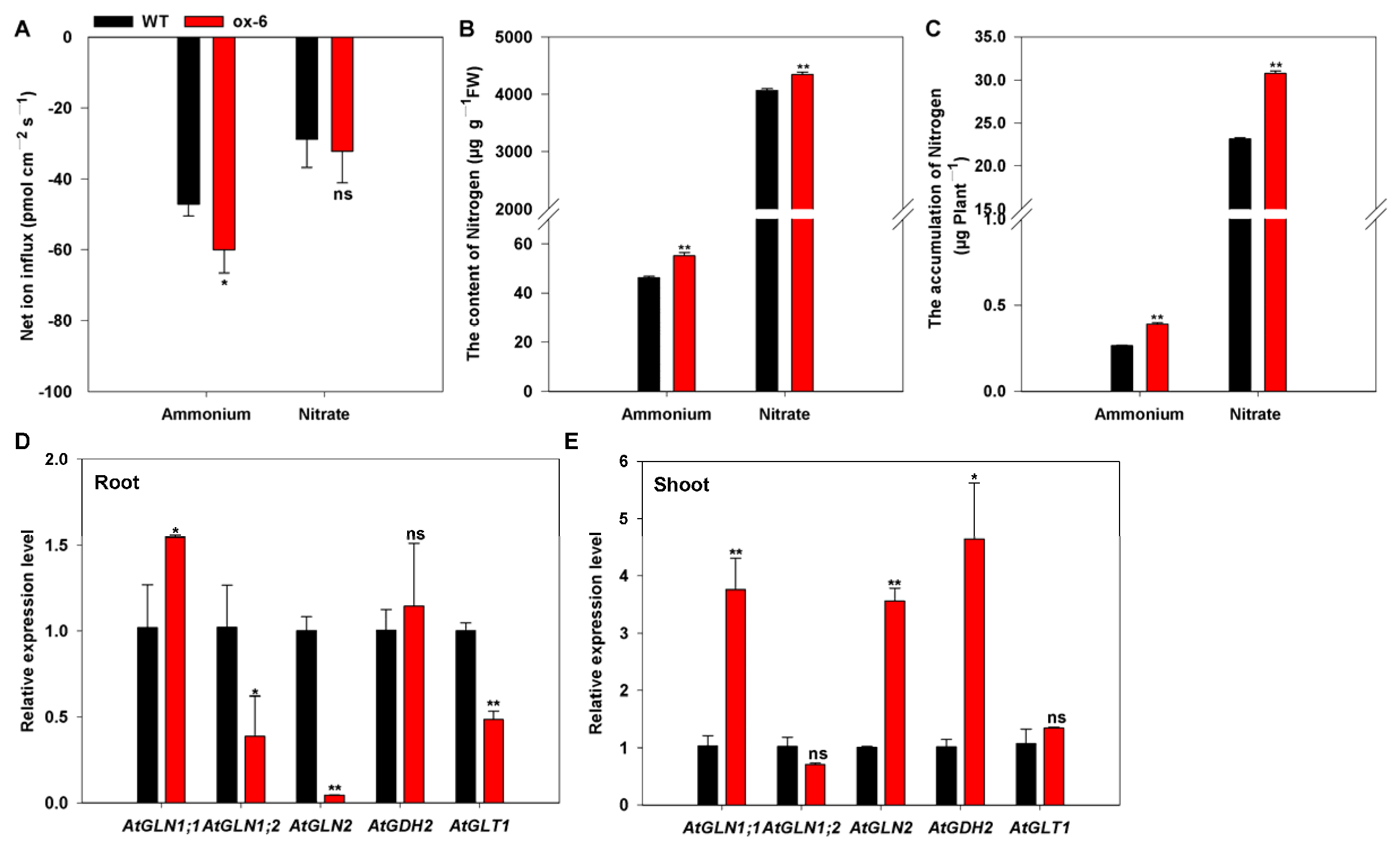
3.7. Heterologous Expression of BcAMT1;5 in Arabidopsis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gu, J.; Yang, J. Nitrogen (N) transformation in paddy rice field: Its effect on N uptake and relation to improved N management. Crop Environ. 2022, 1, 7–14. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Asensio, J.S.R.; Cousins, A.B. Carbon dioxide enrichment inhibits nitrate assimilation in wheat and Arabidopsis. Science 2010, 328, 899–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhou, W.; Wu, J.; Xie, K.; Li, X. LjAMT2;2 Promotes ammonium nitrogen transport during arbuscular mycorrhizal fungi symbiosis in Lotus japonicus. Int. J. Mol. Sci. 2022, 23, 9522. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wiren, N. Three functional transporters for constitutive, diurnally regulated, and starvation-induced uptake of ammonium into Arabidopsis roots. Plant Cell 1999, 11, 937–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, D.; Yang, S.; Huang, Y.; Su, Y. Identification of structural elements involved in fine-tuning of the transport activity of the rice ammonium transporter OsAMT1;3. Plant Physiol. Biochem. 2016, 108, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, D.; Zhou, J.; Yang, S.; Qi, W.; Yang, K.; Su, Y. Function and regulation of ammonium transporters in plants. Int. J. Mol. Sci. 2020, 21, 3557. [Google Scholar] [CrossRef] [PubMed]
- McDonald, T.R.; Ward, J.M. Evolution of electrogenic ammonium transporters (AMTs). Front. Plant Sci. 2016, 7, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wirén, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-Type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; von Wirén, N. AtAMT1;4, a pollen-specific high-affinity ammonium transporter of the plasma membrane in Arabidopsis. Plant Cell Physiol. 2009, 50, 13–25. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef]
- Ijato, T.; Porras Murillo, R.; Ganz, P.; Ludewig, U.; Neuhäuser, B. Concentration-dependent physiological and transcriptional adaptations of wheat seedlings to ammonium. Physiol. Plant. 2021, 171, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Konishi, N.; Ma, J.F. Three polarly localized ammonium transporter 1 members are cooperatively responsible for ammonium uptake in rice under low ammonium condition. New Phytol. 2021, 232, 1778–1792. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, J.; Zhang, B.; Hao, Y.; Ma, F. Genome-wide identification and expression analysis of AMT gene family in apple (Malus domestica Borkh.). Horticulturae 2022, 8, 457. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, X.; Hao, Y.; Su, W.; Liu, H.; Sun, G.; Chen, R.; Song, S. Ammonium transporter (BcAMT1.2) mediates the interaction of ammonium and nitrate in Brassica campestris. Front. Plant Sci. 2020, 10, 1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, L.; Huang, X.; Zhu, Y.; Kou, E.; Liu, H.; Sun, G.; Chen, R.; Song, S. Characterization and expression analysis of BcAMT1;4, an ammonium transporter gene in flowering Chinese cabbage. Hortic. Environ. Biotechnol. 2019, 60, 563–572. [Google Scholar] [CrossRef]
- Ludewig, U. Ion transport versus gas conduction: Function of AMT/Rh-type proteins. Transfus. Clin. Biol. 2006, 13, 111–116. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Marini, A.; Springael, J.; Frommer, W.B.; André, B. Cross-talk between ammonium transporters in yeast and interference by the soybean SAT1 protein. Mol. Microbiol. 2000, 35, 378–385. [Google Scholar] [CrossRef]
- Wiktorek-Smagur, A.; Hnatuszko-Konka, K.; Kononowicz, A.K. Flower bud dipping or vacuum infiltration-two methods of Arabidopsis thaliana transformation. Russ. J. Plant Physl. 2009, 56, 560–568. [Google Scholar] [CrossRef]
- Ivančič, I.; Degobbis, D. An optimal manual procedure for ammonia analysis in natural waters by the indophenol blue method. Water Res. 1984, 18, 1143–1147. [Google Scholar] [CrossRef]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytol. 2007, 174, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; He, Z.; Huang, X.; Zhong, L.; Liu, H.; Sun, G.; Chen, R. cloning and characterization of the ammonium transporter genes BaAMT1;1 and BaAMT1;3 from Chinese kale. Hortic. Environ. Biotechnol. 2017, 58, 178–186. [Google Scholar] [CrossRef]
- Li, H.; Yang, Q.S.; Liu, W.; Lin, J.; Chang, Y.H. The AMT1 family genes from Malus robusta display differential transcription features and ammonium transport abilities. Mol. Biol. Rep. 2017, 44, 379–390. [Google Scholar] [CrossRef]
- Ninnemann, O.; Jauniaux, J.C.; Frommer, W.B. Identification of a high affinity NH4+ transporter from plants. EMBO J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, T.; Sakakibara, H. Interactions between nitrate and ammonium in their uptake, allocation, assimilation, and signaling in plants. J. Exp. Bot. 2017, 68, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Jian, S.; Liao, Q.; Song, H.; Liu, Q.; Lepo, J.E.; Guan, C.; Zhang, J.; Ismail, A.M.; Zhang, Z.; Abdelbagi, M.; et al. NRT1.1-related NH4+ toxicity is associated with a disturbed balance between NH4+ uptake and assimilation. Plant Physiol. 2018, 178, 1473–1488. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Cong, Y.; Chang, Y.; Lin, J. Two AMT2-Type ammonium transporters from Pyrus betulaefolia demonstrate distinct expression characteristics. Plant Mol. Biol. Rep. 2016, 34, 707–719. [Google Scholar] [CrossRef]
- Hachiya, T.; Mizokami, Y.; Miyata, K.; Tholen, D.; Watanabe, C.K.; Noguchi, K. Evidence for a nitrate-independent function of the nitrate sensor NRT1.1 in Arabidopsis thaliana. J. Plant Res. 2011, 124, 425–430. [Google Scholar] [CrossRef]
- Camañes, G.; Cerezo, M.; Primo-Millo, E.; Gojon, A.; García-Agustín, P. Ammonium transport and CitAMT1 expression are regulated by N in Citrus plants. Planta 2009, 229, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Hao, Y.; Liu, H.; Sun, G.; Chen, R.; Song, S. Identification and characterization of two ammonium transporter genes in flowering Chinese cabbage (Brassica campestris). Plant Biotechnol. 2018, 35, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.; Zhu, M.; Jia, L.; Xie, Y.; Wang, Z.; Xuan, W. Ammonium transporters cooperatively regulate rice crown root formation responding to ammonium nitrogen. J. Exp. Bot. 2022, 73, 3671–3685. [Google Scholar] [CrossRef] [PubMed]
- Bittsánszky, A.; Pilinszky, K.; Gyulai, G.; Komives, T. Overcoming ammonium toxicity. Plant Sci. 2015, 231, 184–190. [Google Scholar] [CrossRef] [PubMed]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting nitrogen metabolism and transport processes to improve plant nitrogen use efficiency. Front. Plant Sci. 2021, 11, 628336. [Google Scholar] [CrossRef]
- Liu, Y.; von Wirén, N. Ammonium as a signal for physiological and morphological responses in plants. J. Exp. Bot. 2017, 68, 2581–2592. [Google Scholar] [CrossRef] [Green Version]
- Guan, M.; Thomas, C.D.B.; Carsten, P.; Schjoerring, J.K. Cytosolic glutamine synthetase Gln1;2 is the main isozyme contributing to GS1 activity and can be up-regulated to relieve ammonium toxicity. Plant Physiol. 2016, 171, 1921–1933. [Google Scholar] [CrossRef] [Green Version]
- Pereira, E.G.; Sperandio, M.V.L.; Santos, L.A.; Bucher, C.A.; Coelho, C.P.; Fernandes, M.S. Rice varieties with contrasting nitrogen use efficiency present different expression of amino acid transporters and ammonium transporters. Arch. Agron. Soil Sci. 2022, 1–15. [Google Scholar] [CrossRef]
- Ranathunge, K.; El-kereamy, A.; Gidda, S.; Bi, Y.; Rothstein, S.J. AMT1;1 transgenic rice plants with enhanced NH4+ permeability show superior growth and higher yield under optimal and suboptimal NH4+ conditions. J. Exp. Bot. 2014, 65, 965–979. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Marmagne, A.; Park, J.; Fabien, C.; Yim, Y.; Kim, S.J.; Kim, T.H.; Lim, P.O.; Masclaux Daubresse, C.; Nam, H.G. Concurrent activation of OsAMT1;2 and OsGOGAT1 in rice leads to enhanced nitrogen use efficiency under nitrogen limitation. Plant J. 2020, 103, 7–20. [Google Scholar] [CrossRef]
- Hui, J.; Liu, Z.; Duan, F.; Zhao, Y.; Li, X.; An, X.; Wu, X.; Yuan, L. Ammonium-dependent regulation of ammonium transporter ZmAMT1s expression conferred by glutamine levels in roots of maize. J. Integr. Agr. 2022, 21, 2413–2421. [Google Scholar] [CrossRef]
- Hachiya, T.; Inaba, J.; Wakazaki, M.; Sato, M.; Toyooka, K.; Miyagi, A.; Kawai-Yamada, M.; Sugiura, D.; Nakagawa, T.; Kiba, T.; et al. Excessive ammonium assimilation by plastidic glutamine synthetase causes ammonium toxicity in Arabidopsis thaliana. Nat. Commun. 2021, 12, 4944. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Zhong, L.; Huang, X.; Su, W.; Liu, H.; Sun, G.; Song, S.; Chen, R. BcAMT1;5 Mediates Nitrogen Uptake and Assimilation in Flowering Chinese Cabbage and Improves Plant Growth When Overexpressed in Arabidopsis. Horticulturae 2023, 9, 43. https://doi.org/10.3390/horticulturae9010043
Zhu Y, Zhong L, Huang X, Su W, Liu H, Sun G, Song S, Chen R. BcAMT1;5 Mediates Nitrogen Uptake and Assimilation in Flowering Chinese Cabbage and Improves Plant Growth When Overexpressed in Arabidopsis. Horticulturae. 2023; 9(1):43. https://doi.org/10.3390/horticulturae9010043
Chicago/Turabian StyleZhu, Yunna, Lihua Zhong, Xinmin Huang, Wei Su, Houcheng Liu, Guangwen Sun, Shiwei Song, and Riyuan Chen. 2023. "BcAMT1;5 Mediates Nitrogen Uptake and Assimilation in Flowering Chinese Cabbage and Improves Plant Growth When Overexpressed in Arabidopsis" Horticulturae 9, no. 1: 43. https://doi.org/10.3390/horticulturae9010043