

‘Miguang’ Grape Response to Pergola and Single-Curtain Training Systems
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Field Site
2.2. Experimental Design
2.3. Determination of Microclimate
2.4. Determination of Light Interception
2.5. Determination of Spectral Absorptance
2.6. Determination of Leaf Area
2.7. Determination of Chlorophyll and Carotenoid Contents
2.8. Determination of Leaf Net Photosynthetic Rate
2.9. Determination of Shoot Net Photosynthetic Rate
2.10. 13C Labeling and Calculation of 13C
2.11. Determination of Shoot Soluble Sugar and Starch Contents
2.12. Statistical Analysis
3. Results
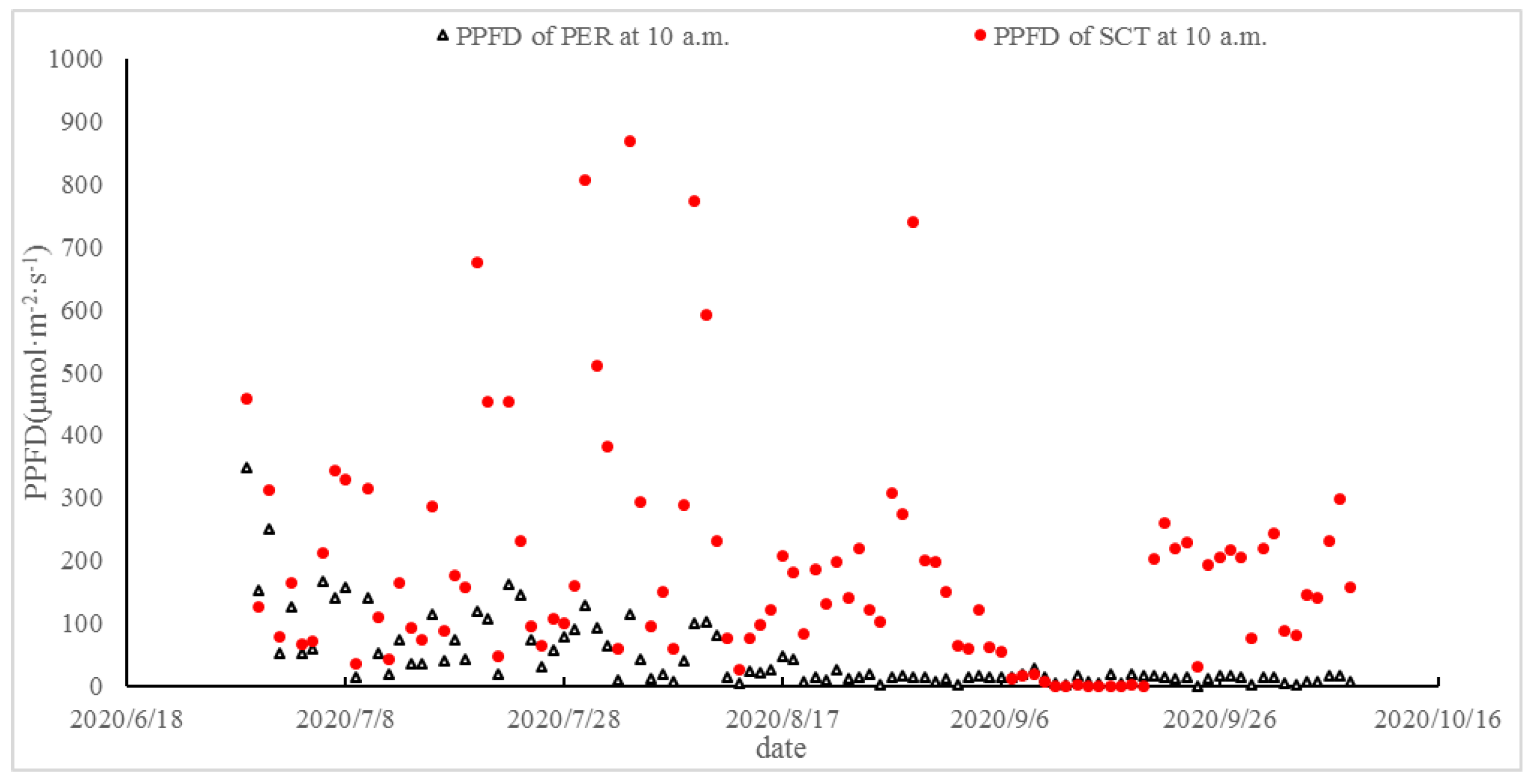
3.1. Effects of PER and SCT on the Light Intensity in the Cluster Zone
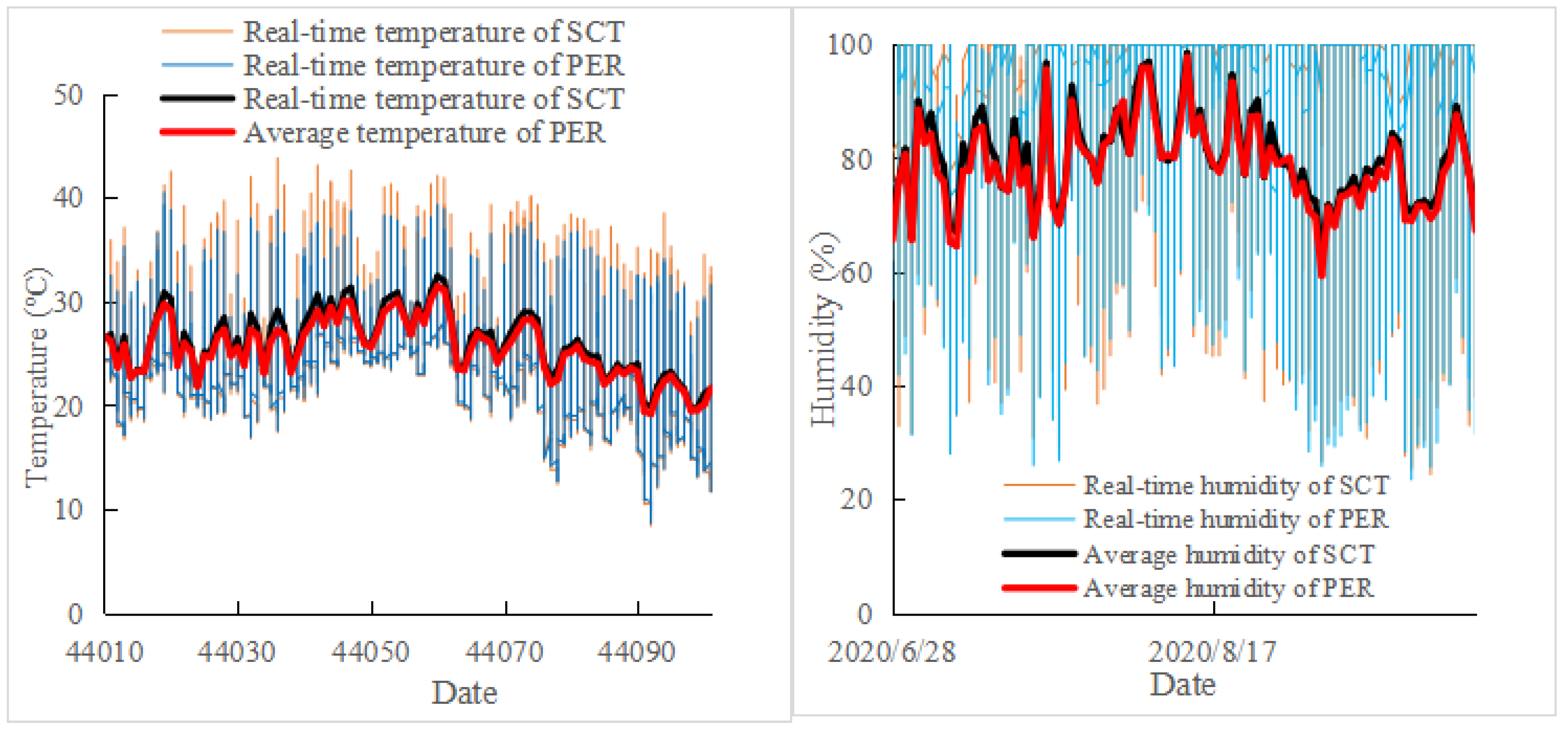
3.2. Effects of PER and SCT on Temperature in the Cluster Zone
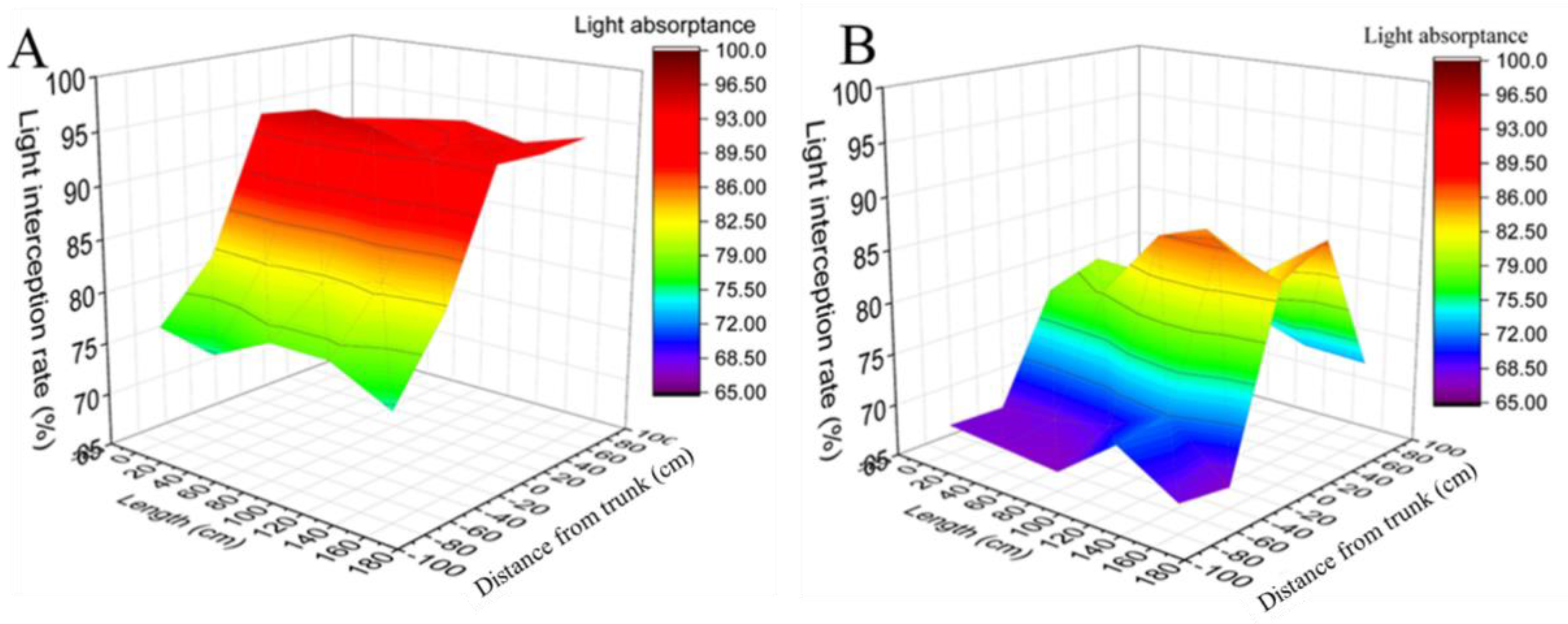
3.3. Effects of PER and SCT on Light Interception
3.3.1. Light Interception Rate
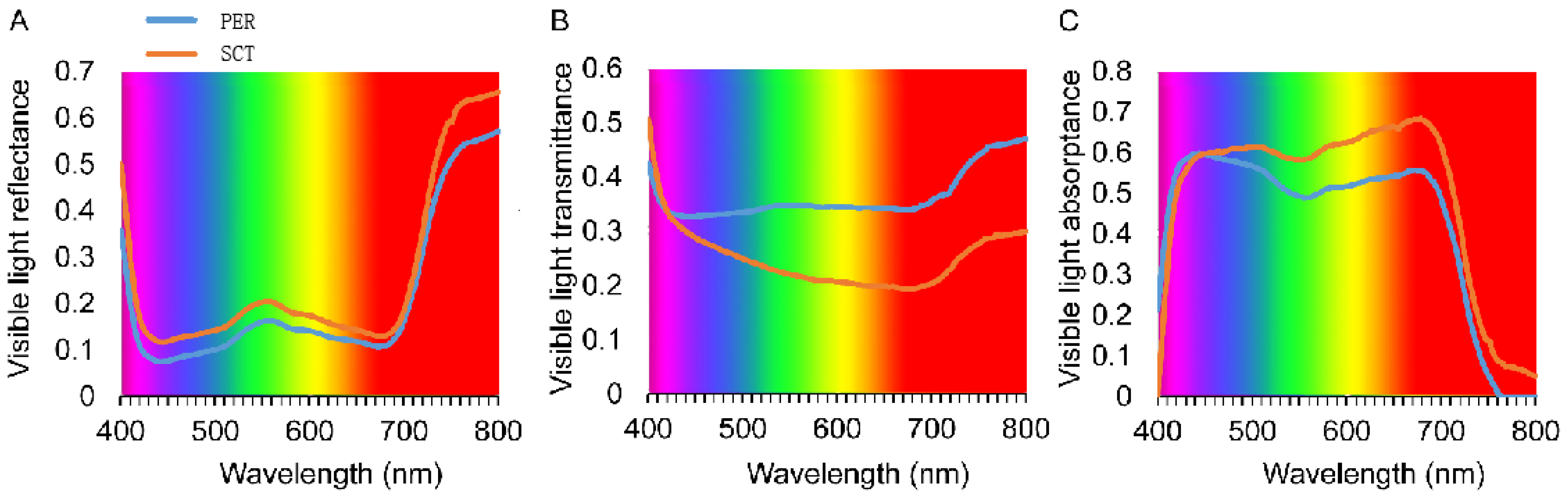
3.3.2. Spectral Reflectance, Transmittance, and Absorptance
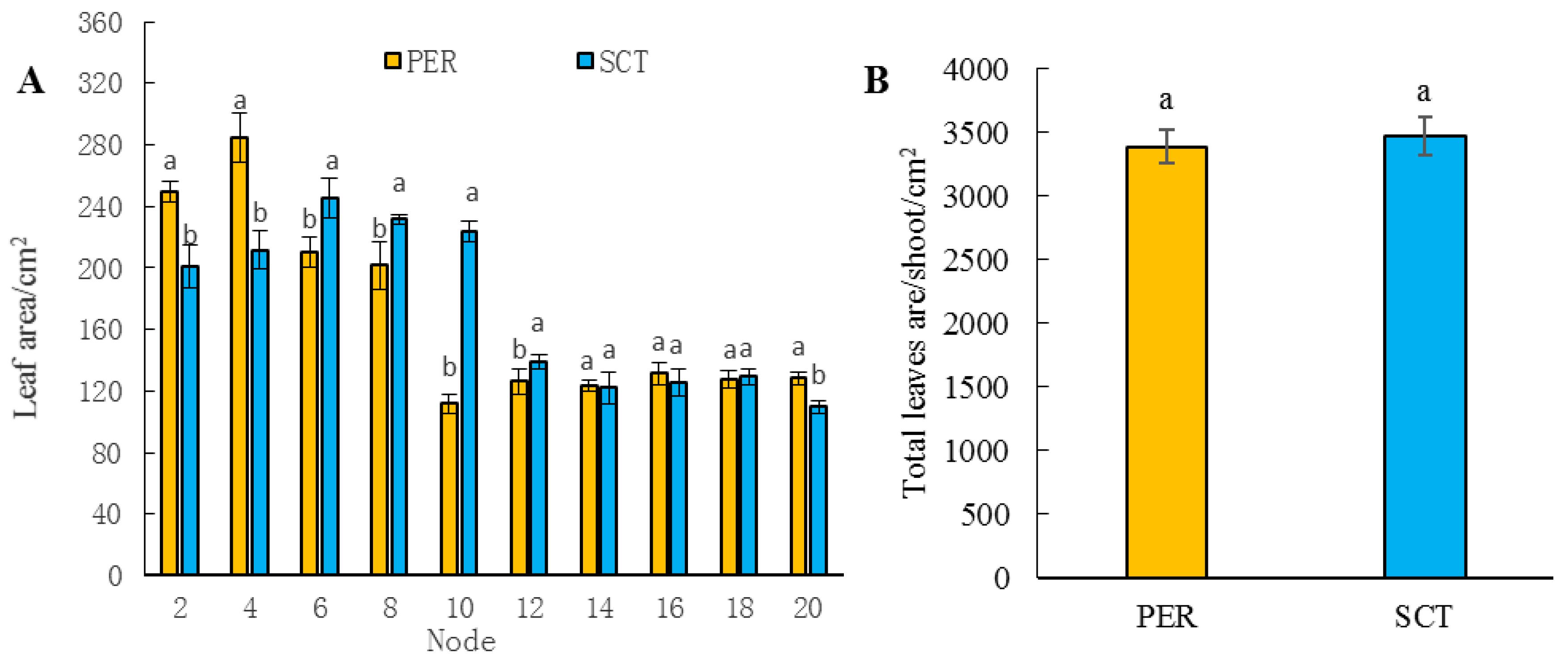
3.4. Effects of PER and SCT on Leaf Area
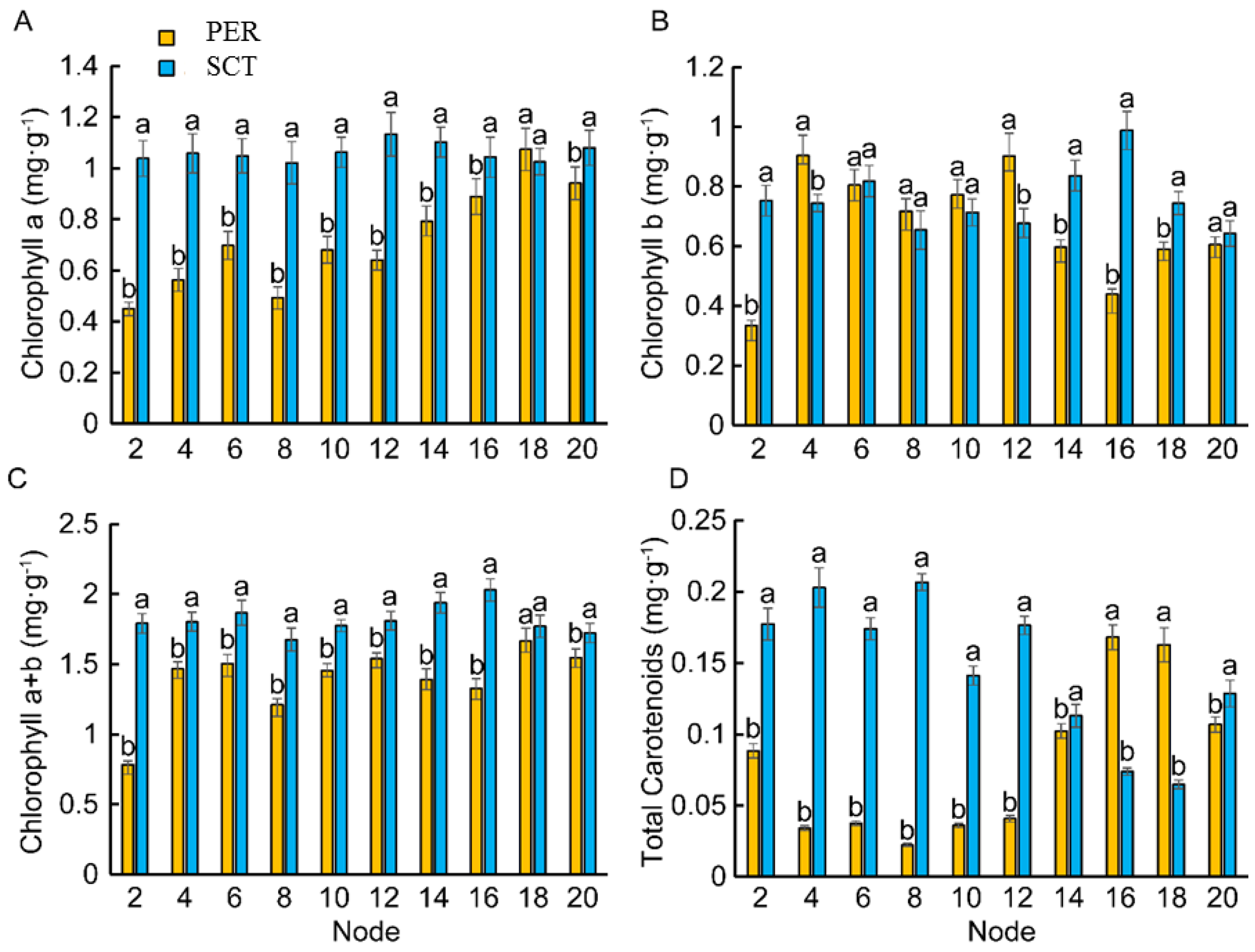
3.5. Effects of PER and SCT on Leaf Chlorophyll Content in Different Nodes
3.6. Effects of PER and SCT on Leaf Photosynthetic Rates in Different Nodes
3.7. Effects of PER and SCT on Assimilate Distribution
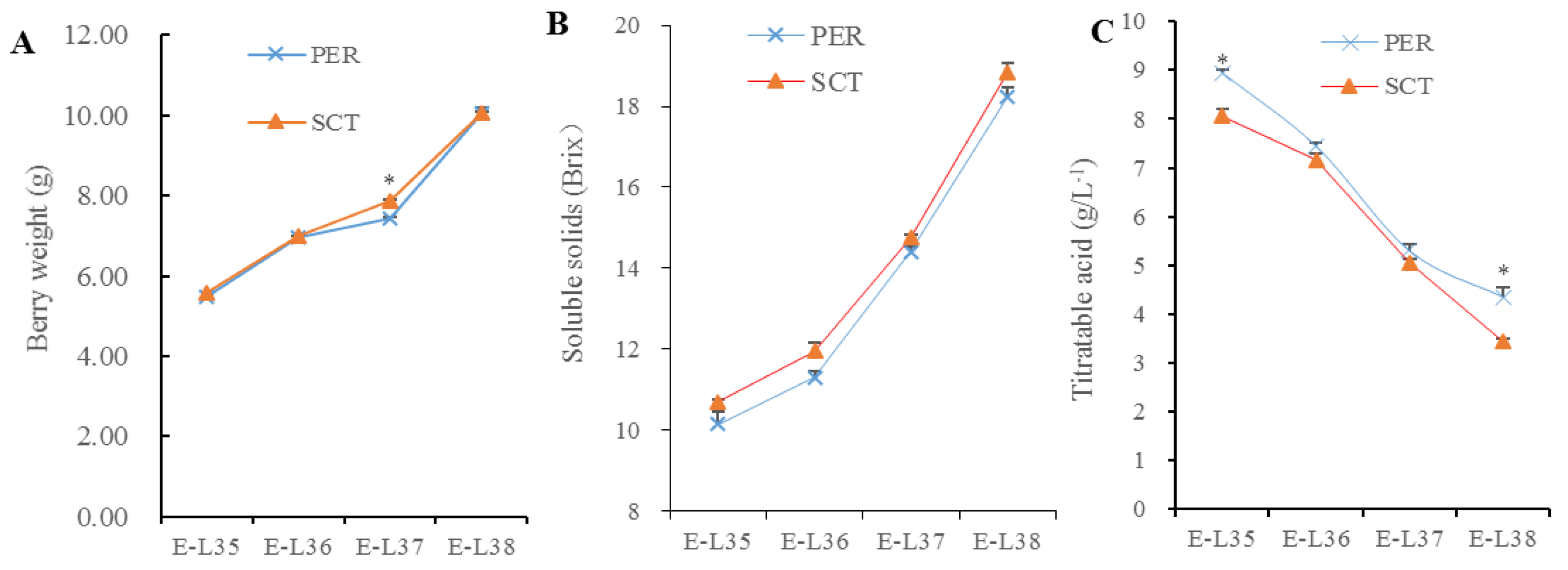
3.8. Effects of PER and SCT on Berry Weight, Soluble Solids, and Titratable Acid Content
3.9. Effects of PER and SCT on Sugar and Starch Storage in Canes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willaume, M.; Lauri, P.; Sinoquet, H. Light interception in apple trees influenced by canopy architecture manipulation. Trees 2004, 18, 705–713. [Google Scholar] [CrossRef]
- Gladstone, E.A.; Dokoozlian, N.K. Influence of leaf area density and trellis/training system on the light microclimate within grapevine canopies. Vitis 2003, 42, 123–131. [Google Scholar]
- Reynolds, A.G.; Heuvel, J.E.V. Influence of Grapevine Training Systems on Vine Growth and Fruit Composition: A Review. Am. J. Enol. Vitic. 2009, 60, 251–268. [Google Scholar] [CrossRef]
- Wolf, T.K.; Dry, P.R.; Iland, P.G.; Botting, D.; Dick, J.; Kennedy, U.; Ristic, R. Response of Shiraz grapevines to five different training systems in the Barossa Valley, Australia. Aust. J. Grape Wine Res. 2003, 9, 82–95. [Google Scholar] [CrossRef]
- Stewart, D.W.; Costa, C.; Dwyer, L.M.; Smith, D.L.; Hamilton, R.I.; Ma, B.L. Canopy Structure, Light Interception, and Photosynthesis in Maize. Agron. J. 2003, 95, 1465–1474. [Google Scholar] [CrossRef]
- Wünsche, J.; Lakso, A.N. Apple tree physiology: Implications for orchard and tree management. Compact. Fruit Tree 2000, 33, 82–88. [Google Scholar]
- Gregan, S.M.; Wargent, J.J.; Liu, L.; Shinkle, J.; Hofann, R.; Winefield, C.; Troutht, M.; Jordan, B. Effects of solar ultraviolet radiation and canopy manipulation on the biochemical composition of Sauvignon Blanc grapes. Aust. J. Grape Wine Res. 2012, 18, 227–238. [Google Scholar] [CrossRef]
- Fira, A.; Joshee, N.; Cristea, V.; Simu, M.; Monica, H.; Pamfil, D.; Clapa, D. Optimization of Micropropagation Protocol for Goji Berry (Lycium barbarum L.). Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Horticulture 2016, 73, 141–150. [Google Scholar]
- Liu, M.Y.; Chi, M.; Tang, Y.H.; Song, C.Z.; Xi, Z.M.; Zhang, Z.W. Effect of Three Training Systems on Grapes in a Wet Region of China: Yield, Incidence of Disease and Anthocyanin Compositions of Vitis vinifera cv. Cabernet Sauvignon. Molecules 2015, 20, 18967–18987. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Q.; Xie, X.L.; Chen, N.; Wang, H.; Li, H. Study on current status and climatic characteristics of wine regions in China. Vitis 2018, 57, 9–16. [Google Scholar]
- Meng, J.F.; Ning, P.F.; Xu, T.F.; Zhang, Z.W. Effect of rain-shelter cultivation of Vitis vinifera cv. Cabernet Gernischet on the phenolic profile of berry skins and the incidence of grape diseases. Molecules 2012, 18, 381–397. [Google Scholar] [CrossRef]
- Di Lorenzo, R.; Gambino, C.; Scafidi, P. Summer pruning in table grape. Adv. Hortic. Sci. 2011, 25, 143–150. [Google Scholar]
- He, L.F.; Li, X.X.; Gao, Y.Y.; Li, B.X.; Mu, W.; Liu, F. Characterization and fungicide sensitivity of Colletotrichum spp. from different hosts in Shandong, China. Plant Dis. 2019, 103, 34–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.Y.; Zhang, Y.; Cao, X.J.; Guo, R.R.; Lin, L.; Cheng, C.F. Introduction Performance of New Labor-Saving Grape Variety Miguang in Southern Guangxi and Its Two-Harvest-A-Year Cultivation Technique. Asian Agric. Res. 2020, 12, 49–51. [Google Scholar]
- Montgomery, R.A.; Chazdon, R.L. Forest Structure, Canopy Architecture, and Light Transmittance in Tropical Wet Forests. Ecology 2001, 82, 2707–2718. [Google Scholar] [CrossRef]
- Gao, X.X.; Dong, S.K.; Li, S.; Xu, Y.D.; Liu, S.L.; Zhao, H.D.; Yeomans, J.; Li, Y.; Shen, H.; Wu, S.N.; et al. Using the random forest model and validated MODIS with the field spectrometer measurement promote the accuracy of estimating aboveground biomass and coverage of alpine grasslands on the Qinghai-Tibetan Plateau. Ecol. Indic. 2020, 112, 106114. [Google Scholar] [CrossRef]
- Serrano, M.A.; Moreno, J.C. Spectral transmission of solar radiation by plastic and glass materials. J. Photochem. Photobiol. B Biol. 2020, 208, 111894. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method Enzymol. 1987, 148, 350–382. [Google Scholar]
- Wang, F.; Sha, J.; Chen, Q.; Xu, X.; Zhu, Z.; Ge, S.; Jiang, Y. Exogenous Abscisic Acid Regulates Distribution of 13C and 15N and Anthocyanin Synthesis in ‘Red Fuji’ Apple Fruit under High Nitrogen Supply. Front. Plant Sci. 2020, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa, L.; Carbonneau, A.; Kelner, J.J. The shoot system architecture of Vitis vinifera ssp. sativa. Sci. Hortic. 2021, 288, 110404. [Google Scholar] [CrossRef]
- Samant, D.; Kishore, K.; Singh, H.S. Branch Bending for Crop Regulation in Guava under Hot and Humid Climate of Eastern India. J. Indian Soc. Coast. Agric. Res. 2016, 34, 92–96. [Google Scholar]
- McIntyre, G. The role of water in the regulation of plant development. Can. J. Bot. 1987, 65, 1287–1298. [Google Scholar] [CrossRef]
- Mamun, A.A.; Rahman, M.H.; Farooque, A.M. Effect of bending and fruit thinning for offseason production of guava. J. Agrof. Environ. 2012, 1, 111–116. [Google Scholar]
- Zheng, Z.L. Carbon and nitrogen nutrient balance signaling in plants. Plant Signal. Behav. 2009, 4, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, H.; Zhang, Y.; Yi, X.; Zhang, X.; Zhang, W. Cotton responds to different plant population densities by adjusting specific leaf area to optimize canopy photosynthetic use efficiency of light and nitrogen. Field Crops Res. 2016, 188, 10–16. [Google Scholar] [CrossRef]
- Buttery, B.R.; Buzzell, R.I. The relationship between chlorophyll content and rate of photosynthesis in soybeans. Can. J. Plant Sci. 1977, 57, 1–5. [Google Scholar] [CrossRef]
- Smart, R.E. Sunlight interception by vineyard. Am. J. Enol. Vitic. 1973, 24, 141–147. [Google Scholar]
- Ferrari, V.; Disegna, E.; Dellacass, E.; Coniberti, A. Influence of timing and intensity of fruit zone leaf removal and kaolin applications on bunch rot control and quality improvement of Sauvignon blanc grapes, and wines, in a temperate humid climate. Sci. Hortic. 2017, 223, 62–71. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Node | Berry Expansion Stage | Veraison | Harvest | |||
|---|---|---|---|---|---|---|
| PER | SCT | PER | SCT | PER | SCT | |
| 2nd | 5.6 ± 0.32b | 8.4 ± 0.43a | 3.4 ± 0.12b | 3.7 ± 0.25a | −2.7 ± 0.18a | −3 ± 0.12a |
| 4th | 7.2 ± 0.55b | 8.8 ± 0.28a | 4.7 ± 0.32a | 5.1 ± 0.43a | 3.6 ± 0.20a | 2.0 ± 0.15b |
| 6th | 10.5 ± 0.43b | 16.1 ± 0.19a | 4.1 ± 0.24a | 4.4 ± 0.36a | 4.1 ± 0.23a | 3.3 ± 0.29b |
| 8th | 9.3 ± 0.46b | 10.8 ± 0.21a | 8.4 ± 0.33a | 6.1 ± 0.42b | 8.3 ± 0.34b | 9.4 ± 0.45a |
| 10th | 9.8 ± 0.49a | 9.5 ± 0.79a | 10.4 ± 0.94b | 13.5 ± 0.85a | 16.5 ± 0.89a | 16.6 ± 0.67a |
| 12th | 8.2 ± 0.58a | 8.8 ± 0.14a | 15.5 ± 0.55b | 18.0 ± 0.76a | 17.2 ± 0.96a | 16.9 ± 1.03a |
| 14th | 7.9 ± 0.67a | 8.2 ± 0.51a | 16.2 ± 0.52b | 17.2 ± 0.68a | 18.0 ± 0.86b | 23.2 ± 0.97a |
| 16th | 12.7 ± 0.94a | 7.9 ± 0.39b | 12.4 ± 0.66b | 17.8 ± 1.06a | 16.6 ± 1.09b | 19.3 ± 1.23a |
| 18th | 13.1 ± 0.68a | 5.6 ± 0.24b | 13.8 ± 0.49a | 14.6 ± 0.34a | 16.3 ± 0.77b | 19.6 ± 0.99a |
| 20th | 6.9 ± 0.33a | 2.3 ± 0.19b | 12.2 ± 0.73a | 10.7 ± 0.55b | 12.2 ± 0.51b | 14.3 ± 0.76a |
| 22nd | —— | —— | 6.6 ± 0.21a | 5.1 ± 0.13b | 13.3 ± 0.34b | 15.9 ± 0.92a |
| 24th | —— | —— | 2.7 ± 0.14a | 2.0 ± 0.09b | 10.3 ± 0.86b | 14.1 ± 0.44a |
| Shoot photosynthetic capacity | 3.1 ± 0.14 b | 5.3 ± 0.22 a | ||||
| PER | SCT | |
|---|---|---|
| Berry weight g | 8.13 ± 1.86a | 7.92 ± 1.58a |
| Soluble solids Brix | 18.47 ± 0.22a | 18.20 ± 0.36a |
| Titratable acid g/L | 5.33 ± 0.21a | 5.21 ± 0.43a |
| Index | Treatment | 4th | 5th | 6th | 7th |
|---|---|---|---|---|---|
| Soluble sugar (mg/g) | PER | 124.63 ± 10.54b | 217.77 ± 7.1b | 233.45 ± 6.92b | 315.42 ± 20.05a |
| SCT | 287.60 ± 19.05a | 299.25 ± 13.27a | 271.34 ± 8.15a | 209.29 ± 13.84b | |
| Starch (mg/g) | PER | 77.03 ± 4.91b | 59.76 ± 2.28a | 49.30 ± 1.99a | 47.93 ± 1.69a |
| SCT | 97.76 ± 5.68a | 47.04 ± 2.97b | 44.88 ± 3.64a | 41.28 ± 0.78b | |
| Internode length (cm) | PER | 9.01 ± 0.24a | 8.56 ± 0.17a | 8.95 ± 0.14a | 8.72 ± 0.34a |
| SCT | 8.04 ± 0.33b | 7.97 ± 0.49b | 8.16 ± 0.24b | 8.89 ± 0.41a | |
| Internode diameter (mm) | PER | 11.67 ± 0.24b | 11.52 ± 0.15a | 11.69 ± 0.63a | 12.05 ± 0.21a |
| SCT | 12.27 ± 0.42a | 11.81 ± 0.38a | 11.43 ± 0.45a | 11.37 ± 0.27b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, W.; Li, S.; Du, T.; Huang, W.; Zhang, Y.; Kang, H.; Yao, Y.; Gao, Z.; Du, Y. ‘Miguang’ Grape Response to Pergola and Single-Curtain Training Systems. Horticulturae 2023, 9, 113. https://doi.org/10.3390/horticulturae9010113
Du W, Li S, Du T, Huang W, Zhang Y, Kang H, Yao Y, Gao Z, Du Y. ‘Miguang’ Grape Response to Pergola and Single-Curtain Training Systems. Horticulturae. 2023; 9(1):113. https://doi.org/10.3390/horticulturae9010113
Chicago/Turabian StyleDu, Wensheng, Shangrui Li, Tingting Du, Wenwei Huang, Yifan Zhang, Hui Kang, Yuxin Yao, Zhen Gao, and Yuanpeng Du. 2023. "‘Miguang’ Grape Response to Pergola and Single-Curtain Training Systems" Horticulturae 9, no. 1: 113. https://doi.org/10.3390/horticulturae9010113