
Grafting Eggplant Onto Underutilized Solanum Species and Biostimulatory Action of Azospirillum brasilense Modulate Growth, Yield, NUE and Nutritional and Functional Traits

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Location and Type of Plants
2.2. Azospirillum brasilense Culture Preparation and Inoculation
2.3. Plant Development and Productive Traits
2.4. Fruit Nutritional Traits, Element Concentration, Functional Compounds and Nitrogen Use Efficiency
2.5. Experimental Layout and Statistics
3. Results
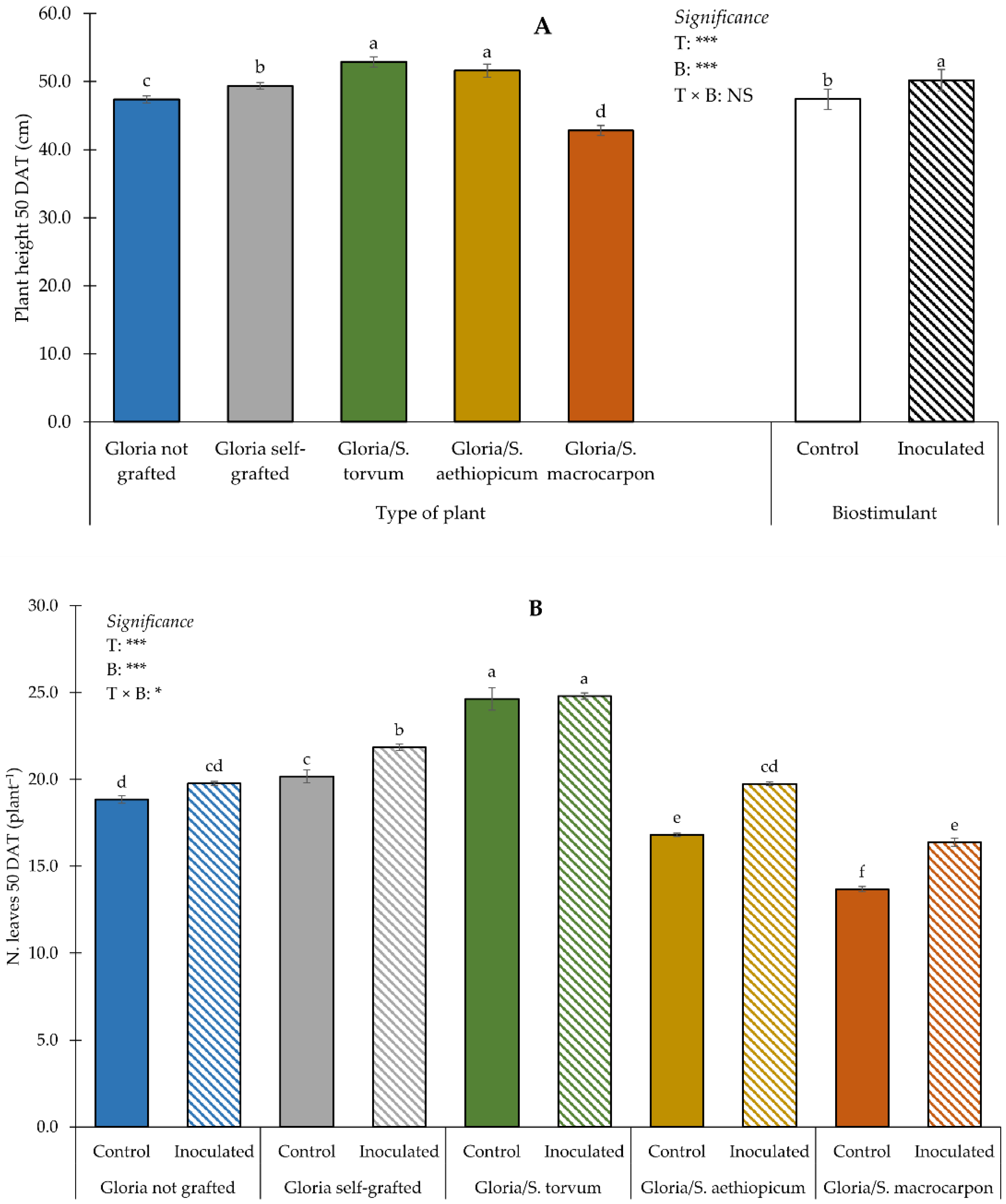
3.1. Plant Growth and Yield
3.2. Fruit Dry Matter, Firmness and Soluble Solids Content
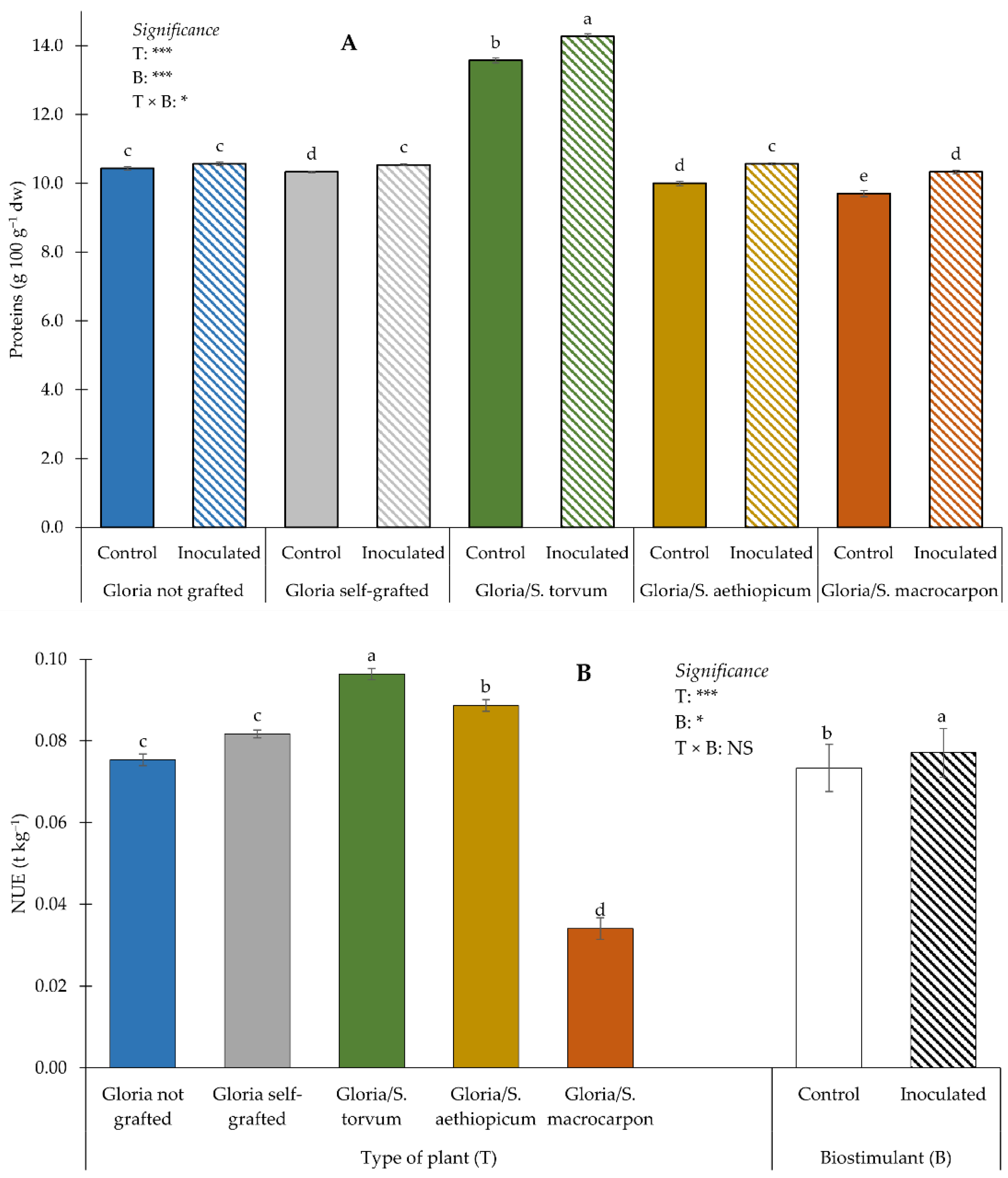
3.3. Minerals, Proteins and NUE
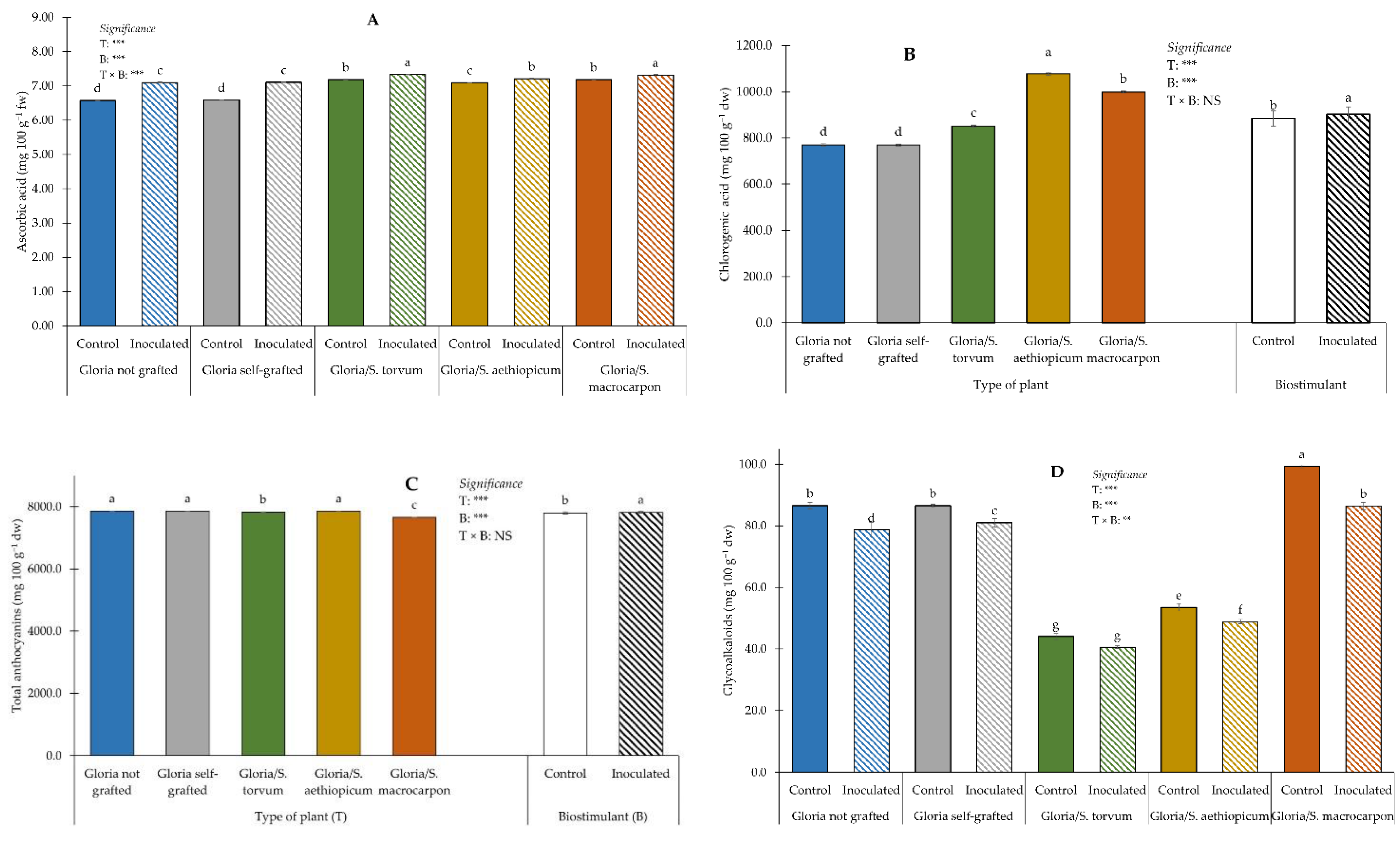
3.4. Fruit Functional Features
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toppino, L.; Prohens, J.; Rotino, G.L.; Plazas, M.; Parisi, M.; Carrizo García, C.; Tripodi, P. Pepper and Eggplant Genetic Resources. In The Wild Solanums Genomes; Carputo, D., Aversano, R., Ercolano, M.R., Eds.; Compendium of Plant Genomes; Springer International Publishing: Cham, Switzerland, 2021; pp. 119–154. [Google Scholar]
- Meyer, R.S.; Karol, K.G.; Little, D.P.; Nee, M.H.; Litt, A. Phylogeographic Relationships among Asian Eggplants and New Perspectives on Eggplant Domestication. Mol. Phylogenet. Evol. 2012, 63, 685–701. [Google Scholar] [CrossRef]
- Lester, R.N.; Daunay, M.C. Diversity of African vegetable Solanum species and its implications for a better understanding of plant domestication. Schr. Genet. Ressour. 2003, 22, 137–152. [Google Scholar]
- Schippers, R.R. African Indigenous Vegetables—An Overview of The Cultivated Species; Natural Resources Institute: Chatham, UK, 2000. [Google Scholar]
- Sunseri, F.; Polignano, G.B.; Alba, V.; Lotti, C.; Visignano, V.; Mennella, G.; D’Alessandro, A.; Bacchi, M.; Riccardi, P.; Fiore, M.C.; et al. Genetic diversity and characterization of African eggplant germplasm collection. Afr. J. Plant. Sci. 2010, 4, 231–241. [Google Scholar]
- Taher, D.; Solberg, S.Ø.; Prohens, J.; Chou, Y.; Rakha, M.; Wu, T. World Vegetable Center eggplant collection: Origin, composition, seed dissemination and utilization in breeding. Front. Plant Sci. 2017, 8, 1484. [Google Scholar] [CrossRef] [Green Version]
- D’Anna, F.; Sabatino, L. Morphological and agronomical characterization of eggplant genetic resources from the Sicily area. J. Food Agric. Environ. 2013, 11, 401–404. [Google Scholar]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; Palazzolo, E.; D’Anna, F. Grafting suitability of Sicilian eggplant ecotypes onto Solanum torvum: Fruit composition, production and phenology. J. Food Agric. Environ. 2013, 11, 1195–1200. [Google Scholar]
- Sabatino, L.; Iapichino, G.; Maggio, A.; D’anna, E.; Bruno, M.; D’Anna, F. Grafting affects yield and phenolic profile of Solanum melongena L. landraces. J. Int. Agric. 2016, 15, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Mauro, R.P.; Rizzo, V.; Leonardi, C.; Mazzaglia, A.; Muratore, G.; Distefano, M.; Sabatino, L.; Giuffrida, F. Influence of Harvest Stage and Rootstock Genotype on Compositional and Sensory Profile of the Elongated Tomato cv. “Sir Elyan”. Agriculture 2020, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; La Bella, S.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C.; Consentino, B.B.; Rouphael, Y. Selenium Biofortification and Grafting Modulate Plant Performance and Functional Features of Cherry Tomato Grown in a Soilless System. Sci. Hortic. 2021, 285, 110095. [Google Scholar] [CrossRef]
- Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S.; Sabatino, L. Agronomic performance and fruit quality in greenhouse grown eggplant are interactively modulated by iodine dosage and grafting. Sci. Hortic. 2022, 295, 110891. [Google Scholar] [CrossRef]
- Bletsos, F.; Thanassoulopoulos, C.; Roupakias, D. Effect of Grafting on Growth, Yield, and Verticillium Wilt of Eggplant. HortScience 2003, 38, 183–186. [Google Scholar] [CrossRef]
- Bogoescu, M.; Doltu, M.; Sora, D. Prevention and control of soilborne diseases and nematodes in eggplants crop by grafting plants combined with soil fumigation. Acta Hortic. 2014, 1044, 331–336. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; D’Anna, F.; Palazzolo, E.; Mennella, G.; Rotino, G.L. Hybrids and allied species as potential rootstocks for eggplant: Effect of grafting on vigour, yield and overall fruit quality traits. Sci. Hortic. 2018, 228, 81–90. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; Rotino, G.L.; Palazzolo, E.; Mennella, G.; D’Anna, F. Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′. Agronomy 2019, 9, 223. [Google Scholar]
- Murata, G.; Uehara, T.; Lee, H.J.; Mizutani, M.; Kadota, Y.; Shinmura, Y.; Uesugi, K. Solanum palinacanthum Dunal as a potential eggplant rootstock resistant to root-knot nematodes. J. Phytopathol. 2022, 170, 185–193. [Google Scholar] [CrossRef]
- Consentino, B.B.; Virga, G.; La Placa, G.G.; Sabatino, L.; Rouphael, Y.; Ntatsi, G.; Iapichino, G.; La Bella, S.; Mauro, R.P.; D’Anna, F.; et al. Celery (Apium graveolens L.) Performances as Subjected to Different Sources of Protein Hydrolysates. Plants 2020, 9, 1633. [Google Scholar] [CrossRef]
- Di Mola, I.; Conti, S.; Cozzolino, E.; Melchionna, G.; Ottaiano, L.; Testa, A.; Sabatino, L.; Rouphael, Y.; Mori, M. Plant-Based Protein Hydrolysate Improves Salinity Tolerance in Hemp: Agronomical and Physiological Aspects. Agronomy 2021, 11, 342. [Google Scholar] [CrossRef]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S. Protein Hydrolysates and Mo-Biofortification Interactively Modulate Plant Performance and Quality of ‘Canasta’ Lettuce Grown in a Protected Environment. Agronomy 2021, 11, 1023. [Google Scholar] [CrossRef]
- La Bella, S.; Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Sabatino, L. Impact of Ecklonia maxima Seaweed Extract and Mo Foliar Treatments on Biofortification, Spinach Yield, Quality and NUE. Plants 2021, 10, 1139. [Google Scholar] [CrossRef]
- Consentino, B.B.; Sabatino, L.; Mauro, R.P.; Nicoletto, C.; De Pasquale, C.; Iapichino, G.; La Bella, S. Seaweed Extract Improves Lagenaria siceraria Young Shoot Production, Mineral Profile and Functional Quality. Horticulturae 2021, 7, 549. [Google Scholar] [CrossRef]
- Consentino, B.B.; Aprile, S.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Alibrandi, P.; Sabatino, L. Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition. Agronomy 2022, 12, 236. [Google Scholar] [CrossRef]
- Sabatino, L.; Consentino, B.B.; Ntatsi, G.; La Bella, S.; Baldassano, S.; Rouphael, Y. Stand-Alone or Combinatorial Effects of Grafting and Microbial and Non-Microbial Derived Compounds on Vigour, Yield and Nutritive and Functional Quality of Greenhouse Eggplant. Plants 2022, 11, 1175. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Carillo, P.; Colla, G.; Graziani, G.; Di Mola, I.; Mori, M.; Kyriacou, M.C.; Rouphael, Y.; Soteriou, G.A.; et al. Plant-Derived Biostimulants Differentially Modulate Primary and Secondary Metabolites and Improve the Yield Potential of Red and Green Lettuce Cultivars. Agronomy 2022, 12, 1361. [Google Scholar] [CrossRef]
- Davison, J. Plant Beneficial Bacteria. Nat. Biotechnol. 1988, 6, 282–286. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Lifshitz, R.; Zablotowicz, R.M. Free-Living Bacterial Inocula for Enhancing Crop Productivity. Trends Biotechnol. 1989, 7, 39–44. [Google Scholar] [CrossRef]
- Cassan, F.; García de Salamone, I. Azospirillum: Cell Physiology, Plant Response, Agronomic and Environmental Research in Argentina; Asociación Argentina de Microbiologia: Dean Funes, Argentina, 2008; ISBN 978-987-98475-8-9. [Google Scholar]
- Tien, T.M.; Gaskins, M.H.; Hubbell, D.H. Plant growth substances produced by Azospirillum brasilense and their effect on the growth of pearl millet (Pennisetum americanum L.). Appl. Environ. Microbiol. 1979, 37, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Bottini, R.; Fulchieri, M.; Pearce, D.; Pharis, R. Identification of gibberelins A1, A3, and iso-A3 in cultures of A. lipoferum. Plant Physiol. 1989, 90, 45–47. [Google Scholar] [CrossRef] [Green Version]
- Strzelczyk, E.; Kamper, M.; Li, C. Cytocinin-like-substances and ethylene production by Azospirillum in media with different carbon sources. Microbiol. Res. 1994, 149, 55–60. [Google Scholar] [CrossRef]
- Rodriguez, H.; Gonzalez, T.; Goire, I.; Bashan, Y. Gluconic Acid Production and Phosphate Solubilization by the Plant Growth-Promoting Bacterium Azospirillum spp. Naturwissenschaften 2004, 91, 552–555. [Google Scholar] [CrossRef]
- Correa, O.S.; Romero, A.M.; Soria, M.A.; de Estrada, M. Azospirillum brasilense-plant genotype interactions modify tomato response to bacterial diseases, and root and foliar microbial communities. In Azospirillum sp.: Cell Physiology, Plant Interactions and Agronomic Research in Argentina; Cassán, F.D., Garcia de Salamone, I., Eds.; Asociación Argentina de Microbiologia: Dean Funes, Argentina, 2008; Volume 266, pp. 85–94. [Google Scholar]
- Weller, D.M.; Thomashow, L.S. Current Challenges in Introducing Beneficial Microorganisms into the Rhizosphere. In Molecular Ecology of Rhizosphere Microorganisms; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1994; pp. 1–18. ISBN 978-3-527-61581-0. [Google Scholar]
- Glick, B.R. The enhancement of plant growth by freeliving bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Probanza, A.; Lucas, J.A.; Acero, N.; Gutierrez Mañero, F.J. The influence of native rhizobacteria on european alder (Alnus glutinosa L. Gaertn.) growth. Plant Soil 1996, 182, 59–66. [Google Scholar] [CrossRef]
- Ioannou, N. Integrating soil solarization with grafting on resistant rootstocks for management of soil-borne pathogens of eggplant. J. Hortic. Sci. Biotechnol. 2001, 76, 396–401. [Google Scholar] [CrossRef]
- Ozturk, B.; Ozer, H. Effects of grafting and green manure treatments on postharvest quality of tomatoes. J. Soil Sci. Plant Nutr. 2019, 19, 780–792. [Google Scholar] [CrossRef]
- Joshi, M.; Leskovar, D.; Djidonou, D.; Jifon, J.; Avila, C.; Masabni, J.; Crosby, K. Production Systems and Growing Environments Had Stronger Effects than Grafting on the Nutritional Quality of Tomato. ACS Food Sci. Technol. 2021, 1, 1399–1411. [Google Scholar] [CrossRef]
- Khan, E.M.; Kakava, E.M.A.; Chachalis, D.; Goulas, C. Effect of grafting on growth and yield of tomato (Lycopersicon esculentum Mill.) in greenhouse and open field. J. Appl. Hortic. 2006, 8, 3–7. [Google Scholar]
- Gisbert, C.; Sánchez-Torres, P.; Raigón, M.D.; Nuez, F. Phytophthora capsici resistance evaluation in pepper hybrids: Agronomic performance and fruit quality of pepper grafted plants. J. Food Agric. Environ. 2010, 8, 116–121. [Google Scholar]
- Gisbert, C.; Prohens, J.; Raigón, M.D.; Stommel, J.R.; Nuez, F. Eggplant relatives as sources of variation for developing new rootstocks: Effects of grafting on eggplant yield and fruit apparent quality and composition. Sci. Hortic. 2011, 128, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Tesi, R. Orticoltura Mediterranea Sostenibile; Pàtron Editore: Bologna, Italy, 2010. [Google Scholar]
- Keswani, C.; Prakash, O.; Bharti, N.; Vílchez, J.I.; Sansinenea, E.; Lally, R.D.; Borriss, R.; Singh, S.P.; Gupta, V.K.; Fraceto, L.F.; et al. Re-addressing the biosafety issues of plant growth promoting rhizobacteria. Sci. Total Environ. 2019, 690, 841–852. [Google Scholar] [CrossRef]
- Fogg, D.N.; Wilkinson, N.T. The colorimetric determination of phosphorus. Analist 1958, 83, 406–414. [Google Scholar] [CrossRef]
- Morand, P.; Gullo, J.L. Mineralisation des tissus vegetaux en vue du dosage de P, Ca, Mg, Na, K. Ann. Agron. 1970, 21, 229–236. [Google Scholar]
- Stommel, J.R.; Whitaker, B.D. Phenolic acid content and composition of eggplant fruit in a germplasm core subset. J. Am. Soc. Hortic. Sci. 2003, 128, 704–710. [Google Scholar] [CrossRef]
- Mennella, G.; Lo Scalzo, R.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Acciarri, N.; De Almeida, A.E.; Rotino, G.L. Chemical and bioactive quality traits during fruit ripening in eggplant (S. melongena L.) and allied species. J. Agric. Food Chem. 2012, 60, 11821–11831. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas Putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; Consentino, B.B.; D’Anna, F.; Rouphael, Y. Rootstock and Arbuscular Mycorrhiza Combinatorial Effects on Eggplant Crop Performance and Fruit Quality under Greenhouse Conditions. Agronomy 2020, 10, 693. [Google Scholar] [CrossRef]
- Parewa, H.P.; Yadav, J.; Rakshit, A.; Meena, V.S.; Karthikeyan, N. Plant growth promoting rhizobacteria enhance growth and nutrient uptake of crops. Agric. Sustain. Dev. 2014, 2, 101–116. [Google Scholar]
- Bhattacharyya, P.N.; Jha, D.K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Palmer, J.W.; Harker, F.R.; Tustin, D.S.; Johnston, J. Fruit dry matter concentration: A new quality metric for apples. J. Sci. Food Agric. 2010, 90, 2586–2594. [Google Scholar] [CrossRef]
- Ordookhani, K.; Zare, M. Effect of Pseudomonas, Azotobacter and arbuscular mycorrhiza fungi on lycopene, antioxidant activity and total soluble solid in tomato (Lycopersicon esculentum F1 Hybrid, Delba). Adv. Environ. Biol. 2011, 5, 1290–1294. [Google Scholar]
- Hungria, M.; Campo, R.J.; Souza, E.M.; Pedrosa, F.O. Inoculation with Selected Strains of Azospirillum brasilense and A. lipoferum Improves Yields of Maize and Wheat in Brazil. Plant Soil 2010, 331, 413–425. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Lee, I.J. Gibberellins producing Bacillus methylotrophicus KE2 supports plant growth and enhances nutritional metabolites and food values of lettuce. Plant Physiol. Biochem. 2016, 109, 181–189. [Google Scholar] [CrossRef]
- De Santi Ferrara, F.I.; Oliveira, Z.M.; Gonzales, H.H.S.; Floh, E.I.S.; Barbosa, H.R. Endophytic and Rhizospheric Enterobacteria Isolated from Sugar Cane Have Different Potentials for Producing Plant Growth-Promoting Substances. Plant Soil 2012, 353, 409–417. [Google Scholar] [CrossRef]
- Zeffa, D.M.; Perini, L.J.; Silva, M.B.; de Sousa, N.V.; Scapim, C.A.; de Oliveira, A.L.M.; Júnior, A.T.D.; Gonçalves, L.S.A. Azospirillum brasilense Promotes Increases in Growth and Nitrogen Use Efficiency of Maize Genotypes. PLoS ONE 2019, 14, e0215332. [Google Scholar] [CrossRef] [Green Version]
- Hollman, P.C.; Geelen, A.; Kromhout, D. Dietary flavonol intake may lower stroke risk in men and women. J. Nutr. 2010, 140, 600–604. [Google Scholar] [CrossRef] [Green Version]
- Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cohn, J.S.; Harvey, I.; Le Cornu, K.A.; Ryder, J.J.; Hall, W.L.; Cassidy, A. Flavonoids, flavonoid-rich foods, and cardiovascular risk: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2008, 88, 38–50. [Google Scholar] [CrossRef]
- Oztekin, G.B.; Tuzel, Y.; Tuzel, I.H. Does mycorrhiza improve salinity tolerance in grafted plants? Sci. Hortic. 2013, 149, 55–60. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef]
- Cappellari, L.d.R.; Chiappero, J.; Santoro, M.V.; Giordano, W.; Banchio, E. Inducing Phenolic Production and Volatile Organic Compounds Emission by Inoculating Mentha piperita with Plant Growth-Promoting Rhizobacteria. Sci. Hortic. 2017, 220, 193–198. [Google Scholar] [CrossRef]
- Cassidy, A. Berry anthocyanin intake and cardiovascular health. Mol. Aspects Med. 2018, 61, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C. Plant response to plant growth-promoting rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awad, M.A.; de Jager, A.; van der Plas, L.H.W.; van der Krol, A.R. Flavonoid and chlorogenic acid changes in skin of ‘Elstar’ and ‘Jonagold’ apples during development and ripening. Sci. Hortic. 2001, 90, 69–83. [Google Scholar] [CrossRef]
- Badar, M.; Mhmood, K.; Hassan, I.; Ahmed, M.; Ahmad, I.; Ahmad, N.; Hasan, M. Plant growth promoting bacteria (PGPB) enhance growth and yield of strawberry cultivars. Appl. Ecol. Environ. Res. 2022, 20, 2187–2203. [Google Scholar] [CrossRef]
- Lingua, G.; Bona, E.; Manassero, P.; Marsano, F.; Todeschini, V.; Cantamessa, S.; Copetta, A.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting Pseudomonads increases anthocyanin concentration in strawberry fruits (Fragaria x ananassa var. Selva) in conditions of reduced fertilization. Int. J. Mol. Sci. 2013, 14, 16207–16225. [Google Scholar] [CrossRef]
- Mystkowska, I. Reduction of glycoalkaloids in potato under the influence of biostimulators. Appl. Ecol. Environ. Res. 2019, 17, 3567–3574. [Google Scholar] [CrossRef]
- Papathanasiou, F.; Mitchell, S.H.; Watson, S.; Harvey, B.M.R. Effect of environmental stress during tuber development on accumulation of glycoalkaloids in potato (Solanum tuberosum L.). J. Sci. Food Agric. 1999, 79, 1183–1189. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Total Yield (kg Plant−1) | Marketable Yield (kg Plant−1) | Number of Marketable Fruits (no. Plant−1) | Mean Mass of Marketable Fruits (g Fruit−1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Type of plant (T) | Gloria not grafted | 3.8 | c | 3.5 | c | 9.8 | b | 355.3 | b |
| Gloria self-grafted | 4.1 | c | 3.8 | c | 10.7 | b | 355.9 | b | |
| Gloria/S. torvum | 4.8 | a | 4.6 | a | 12.5 | a | 365.9 | a | |
| Gloria/S. aethiopicum | 4.4 | b | 4.2 | b | 11.9 | a | 349.0 | b | |
| Gloria/S. macrocarpon | 1.7 | d | 1.4 | d | 5.1 | c | 275.0 | c | |
| Biostimulant (B) | Control | 3.7 | b | 3.3 | b | 9.6 | b | 336.0 | b |
| Inoculated | 3.9 | a | 3.6 | a | 10.3 | a | 344.4 | a | |
| Significance | |||||||||
| T | *** | *** | *** | *** | |||||
| B | * | *** | ** | *** | |||||
| T × B | NS | NS | NS | NS | |||||
| Treatments | Fruit Dry Matter (%) | Firmness (N) | SSC (°Brix) | ||||
|---|---|---|---|---|---|---|---|
| Type of plant (T) | Gloria not grafted | 7.1 | a | −46.8 | a | 4.6 | c |
| Gloria self-grafted | 7.1 | a | −46.6 | a | 4.3 | d | |
| Gloria/S. torvum | 7.1 | a | −46.6 | a | 4.1 | e | |
| Gloria/S. aethiopicum | 7.1 | a | −46.8 | a | 4.9 | b | |
| Gloria/S. macrocarpon | 7.2 | a | −46.7 | a | 5.2 | a | |
| Biostimulant (B) | Control | 7.0 | b | −46.0 | a | 4.5 | b |
| Inoculated | 7.2 | a | −47.4 | b | 4.7 | a | |
| Significance | |||||||
| T | NS | NS | *** | ||||
| B | * | *** | *** | ||||
| T × B | NS | NS | NS | ||||
| Treatments | P (mg 100 g−1 dw) | K (mg 100 g−1 dw) | Ca (mg 100 g−1 dw) | Mg (mg 100 g−1 dw) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Type of plant (T) | Gloria not grafted | 543.3 | a | 339.7 | b | 108.7 | a | 19.1 | b |
| Gloria self-grafted | 544.0 | a | 340.0 | b | 109.3 | a | 18.8 | b | |
| Gloria/S. torvum | 541.7 | a | 343.6 | b | 109.0 | a | 14.5 | c | |
| Gloria/S. aethiopicum | 522.9 | b | 301.9 | c | 98.8 | c | 20.6 | a | |
| Gloria/S. macrocarpon | 499.9 | c | 348.2 | a | 106.3 | b | 19.2 | b | |
| Biostimulant (B) | Control | 530.6 | a | 334.2 | a | 106.3 | a | 17.4 | b |
| Inoculated | 530.1 | a | 335.1 | a | 106.5 | a | 19.5 | a | |
| Significance | |||||||||
| T | *** | *** | *** | *** | |||||
| B | NS | NS | NS | *** | |||||
| T × B | NS | NS | NS | NS | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Consentino, B.B.; Sabatino, L.; Vultaggio, L.; Rotino, G.L.; La Placa, G.G.; D’Anna, F.; Leto, C.; Iacuzzi, N.; De Pasquale, C. Grafting Eggplant Onto Underutilized Solanum Species and Biostimulatory Action of Azospirillum brasilense Modulate Growth, Yield, NUE and Nutritional and Functional Traits. Horticulturae 2022, 8, 722. https://doi.org/10.3390/horticulturae8080722
Consentino BB, Sabatino L, Vultaggio L, Rotino GL, La Placa GG, D’Anna F, Leto C, Iacuzzi N, De Pasquale C. Grafting Eggplant Onto Underutilized Solanum Species and Biostimulatory Action of Azospirillum brasilense Modulate Growth, Yield, NUE and Nutritional and Functional Traits. Horticulturae. 2022; 8(8):722. https://doi.org/10.3390/horticulturae8080722
Chicago/Turabian StyleConsentino, Beppe Benedetto, Leo Sabatino, Lorena Vultaggio, Giuseppe Leonardo Rotino, Gaetano Giuseppe La Placa, Fabio D’Anna, Claudio Leto, Nicolò Iacuzzi, and Claudio De Pasquale. 2022. "Grafting Eggplant Onto Underutilized Solanum Species and Biostimulatory Action of Azospirillum brasilense Modulate Growth, Yield, NUE and Nutritional and Functional Traits" Horticulturae 8, no. 8: 722. https://doi.org/10.3390/horticulturae8080722