
Dynamic Changes of Fruit Physiological Quality and Sugar Components during Fruit Growth and Development of Actinidia eriantha

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Measurement of Fruit Physiological Quality
2.3. Determination of Sugar Components
2.4. Determination of Enzymes Activities Related to Sugar Metabolism
2.5. Expression Analysis of Genes Related to Sugar Metabolism
2.6. Data Analysis
3. Results
3.1. Dynamic Changes of Appearance Quality-Related Indexes of ‘Ganlv 1’ during Fruit Development
3.2. Dynamic Changes in Dry Matter and Soluble Solid Content in A. eriantha ‘Ganlv 1’ during Fruit Development
3.3. Determination of Titratable Acid and AsA Contents in ‘Ganlv 1’ during Fruit Development
3.4. Change of Soluble Sugar Components in ‘Ganlv 1’ during Fruit Development
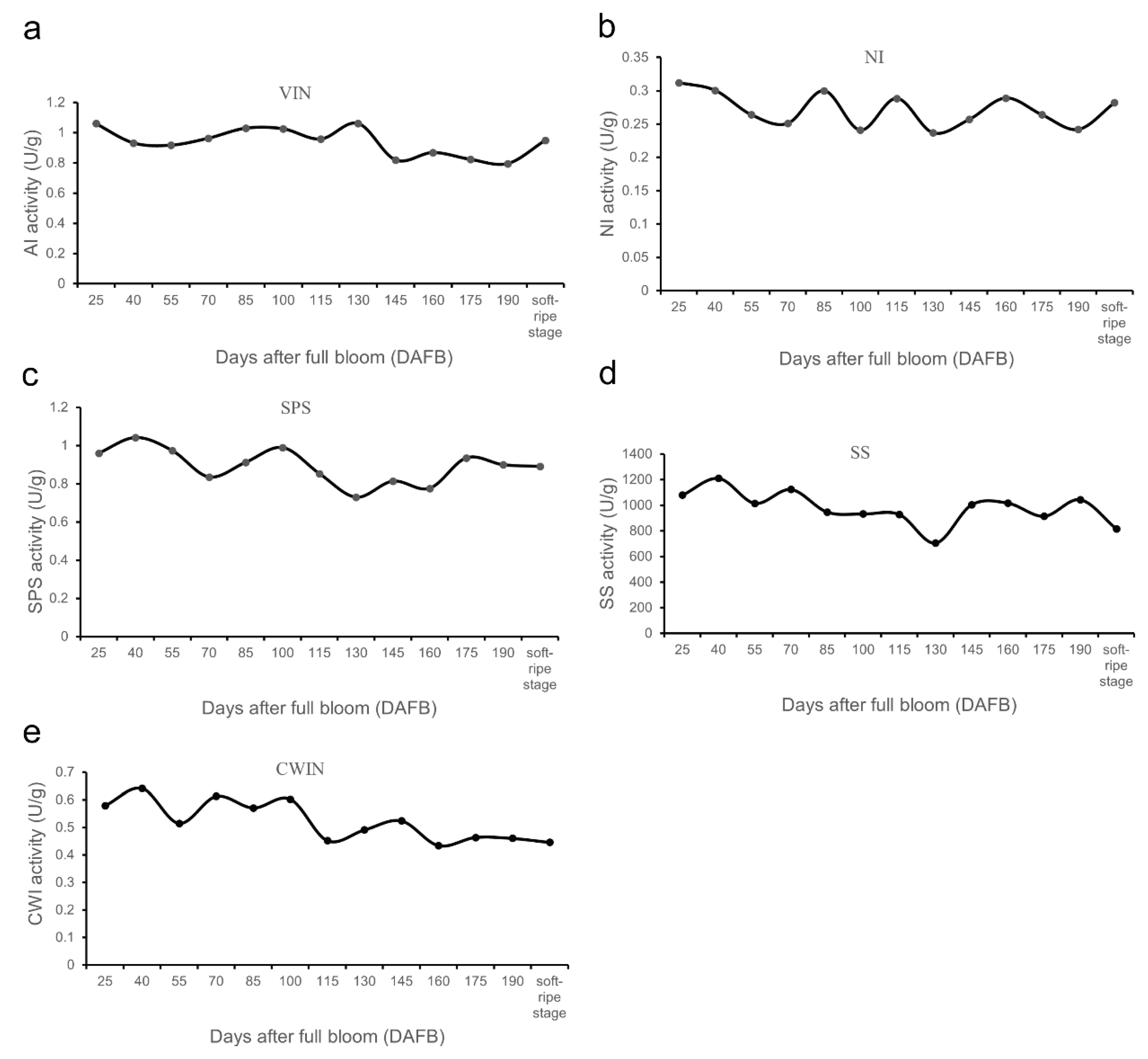
3.5. Changes of Soluble Sugar-Related Metabolic Enzyme Activities during Fruit Development
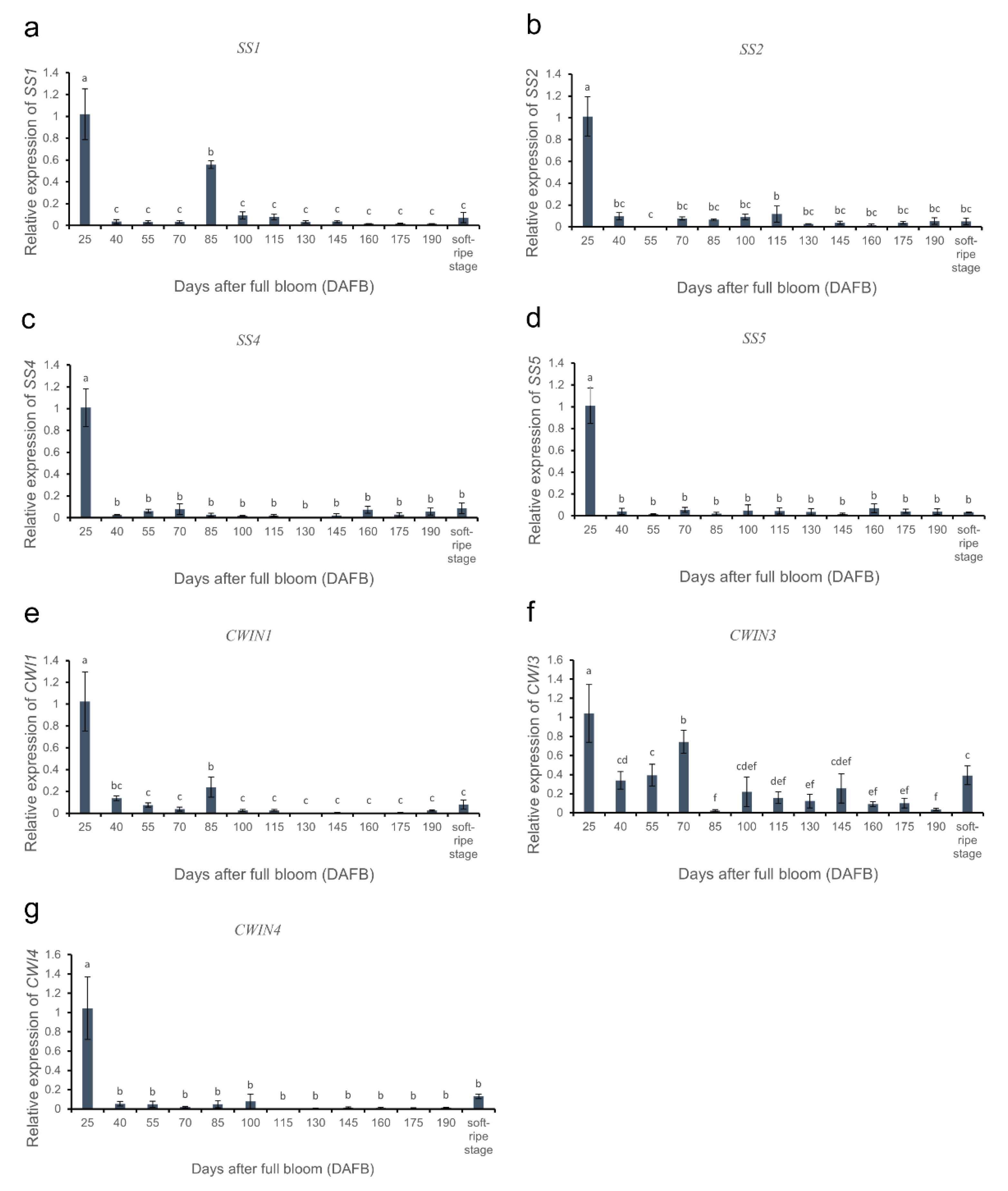
3.6. Gene Expression Analysis of Soluble Sugar Metabolism-Related Enzymes during Fruit Development
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, H.W. Actinidia Germplasm Resources in China; China Forestry Publishing House: Beijing, China, 2013. [Google Scholar]
- Xie, M.; Wu, Y.J.; Jiang, G.H.; Zhang, Q.C.; Zhang, H.Q.; Peng, S.J.; Liu, K.M. A new big fruit Actinidia eriantha Benth. cultivar ‘White’. Acta Hortic. Sin. 2008, 35, 1555. [Google Scholar]
- Xu, X.B.; Huang, C.H.; Qu, X.Y.; Chen, M.; Zhong, M.; Lang, B.B.; Chen, C.J.; Xie, M.; Zhang, W.B. A new easy peeling Actinidia eriantha cultivar ‘Ganmi 6’. Acta Hortic. Sin. 2015, 42, 2539–2540. [Google Scholar]
- Xu, Y.H.; Song, Q.Q.; Hu, B.; Li, S.G.; Zheng, X.L.; Jiang, T.J. Harvest maturity affects quality and storability of Actinidia eriantha cv. White fruit. J. Nucl. Agric. Sci. 2020, 34, 0521–0531. [Google Scholar]
- Qu, X.Y.; Lang, B.B.; Zhong, M.; Zhu, B.; Tao, J.J.; Huang, C.H.; Xu, X.B. Principal component analysis and comprehensive evaluation of fruit quality of Actinidia eriantha. Chin. Agric. Sci. Bull. 2016, 32, 92–96. [Google Scholar]
- Lang, B.B.; Zhu, B.; Xie, M.; Zhang, W.B.; Seyrek, U.A.; Huang, C.H.; Xu, X.B. Variation and probability grading of the main quantitative characteristics of wild Actinidia eriantha germplasm resources. J. Fruit Sci. 2016, 33, 8–15. [Google Scholar]
- Wang, Y.Z.; Pan, Z.M. Comprehensive evaluation of 22 Actinidia eriantha germplasm resources based on principal components analysis. Acta Agric. Zhejiangensis 2021, 33, 825–830. [Google Scholar]
- Zhang, H.Q.; Xie, M.; Xiao, J.P.; Zhou, L.Q.; Song, G.H. Characterization of fruit development of the diploid kiwifruit, Actinidia eriantha ‘White’. J. Fruit Sci. 2015, 32, 238–246. [Google Scholar]
- Chen, C.J.; Tao, J.J.; Qu, X.Y.; Huang, C.H.; Xu, X.B. Dynamic variation in sugar, acid, and AsA contents of ‘Gnami 6’ kiwifruit (Actinidia eriantha Benth) fruits. Agric. Sci. Technol. 2015, 16, 2589–2591, 2595. [Google Scholar]
- Liu, C.H.; Qiu, G.L.; Liu, Z.B.; Yang, Y.; Zhuang, Q.G.; Zhang, Q. Study on the changes of physicochemical properties of A. eriantha during the fruit development. J. Sichuan Univ. (Nat. Sci. Ed.) 2019, 56, 951–956. [Google Scholar]
- Zhang, H.Q.; Xie, M.; Zhang, C.; Yang, L.Q.; Zhang, Z.; Xiao, J.P.; Zhou, L.Q. Difference in starch accumulation and characterization of sugar metabolism during fruit development of kiwifruit. Sci. Agric. Sin. 2014, 47, 3453–3464. [Google Scholar]
- Zheng, Q.M.; Tang, Z.; Xu, Q.; Deng, X.X. Isolation, phylogenetic relationship and expression profiling of sugar transporter genes in sweet orange (Citrus sinensis). Plant Cell Tissue Organ Cult. 2014, 119, 609–624. [Google Scholar] [CrossRef]
- Lee, P.R.; Tan, R.M.; Yu, B.; Curran, P.; Liu, S.Q. Sugars, organic acids, and phenolic acids of exotic seasonable tropical fruits. Nutr. Food Sci. 2013, 43, 267–276. [Google Scholar] [CrossRef]
- Zhao, J.H.; Li, H.X.; Xi, W.P.; An, W.; Niu, L.L.; Cao, Y.L.; Wang, H.F.; Wang, Y.J.; Yin, Y. Changes in sugars and organic acids in wolfberry (Lycium barbarum L.) fruit during development and maturation. Food Chem. 2015, 173, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.N.; Yan, L.Y.; Zhang, B.; Yang, S.B.; Zhao, Z.Y. A study on sugar and organic acid components in different apple cultivars. J. Fruit Sci. 2021, 38, 1877–1889. [Google Scholar]
- Zhang, G.J. Study on Sugars and Acids Inheritance and the Expression Pattern of β-Glucosidase Genes (VvBGs) during Grape Berry Ripening; China Agricultural University: Beijing, China, 2013. [Google Scholar]
- Huang, Q. Studies on Characterization of Organic Acids Metabolism and Expression of Relative Genes during Fruit Development of Relative Genes during Fruit Development of ‘Ganmi 6’; Jiangxi Agricultural University: Nanchang, China, 2019. [Google Scholar]
- Cao, J.K.; Jiang, W.B.; Zhao, Y.M. Experiment Guidance of Postharvest Physiology and Biochemistry of Fruits and Vegetables; Chinese Light Industry Press: Beijing, China, 2007. [Google Scholar]
- Yue, J.Y.; Liu, J.C.; Tang, W.; Wu, Y.Q.; Tang, X.F.; Li, W.; Yang, Y.; Wang, L.H.; Huang, S.X.; Fang, C.; et al. Kiwifruit Genome Database (KGD): A comprehensive resource for kiwifruit genomics. Hortic. Res. 2020, 7, 117. [Google Scholar] [CrossRef]
- Wang, X.X.; Chen, Y.; Jiang, S.; Xu, F.; Wang, H.F.; Wei, Y.Y.; Shao, X.F. PpINH1, an invertase inhibitor, interacts with vacuolar invertase PpVIN2 in regulating the chilling tolerance of peach fruit. Hortic. Res. 2020, 7, 168. [Google Scholar] [CrossRef]
- Chen, M.Y.; Zhang, P.; Zhao, T.T.; Han, F.; Liu, X.L.; Zhong, C.H. Relationship between harvest indices and fruit quality traits in Actinidia chinensis ‘Jintao’. Plant Sci. J. 2019, 37, 621–627. [Google Scholar]
- Nardozza, S.; Gamble, J.; Axten, L.G.; Wohlers, M.W.; Clearwater, M.J.; Feng, J.Q.; Harker, F.R. Dry matter content and fruit size affect flavour and texture of novel Actinidia deliciosa. J. Sci. Food Agric. 2011, 91, 742–748. [Google Scholar] [CrossRef]
- Wu, Y.J.; Xie, M.; Zhang, Q.C.; Jiang, G.H.; Zhang, H.Q.; Long, Q.J.; Han, W.J.; Chen, J.W.; Shong, G.H. Characteristics of ‘White’: A new easy-peel cultivar of Actinidia eriantha. N. Z. J. Crop Hortic. Sci. 2009, 37, 369–373. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.L.; Richardson, A.C.; Costa, G.; Marsh, H.; MacRae, E.A.; Clearwater, M.J. Variation in carbon content and size in developing fruit of Actinidia deliciosa genotypes. Funct. Plant Biol. 2010, 37, 545–554. [Google Scholar] [CrossRef]
- Qi, W.Y.; Zhou, C.H.; Song, L.J.; Zhong, Y.; Zheng, X.L. Study on sugar metabolism of Actinidia eriantha Benth ‘White’ during storage. J. Fruit Sci. 2016, 33, 744–751. [Google Scholar]
- Richardson, A.C.; Boldingh, H.L.; McAtee, P.A.; Gunaseelan, K.; Luo, Z.W.; Atkinson, R.G.; David, K.M.; Burdon, J.N.; Schaffer, R.J. Fruit development of the diploid kiwifruit, Actinidia chinensis ‘Hort16A’. BMC Plant Biol. 2011, 11, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moscatello, S.; Famiani, F.; Proietti, S.; Farinelli, D.; Battistelli, A. Sucrose synthase dominates carbohydrate metabolism and relative growth rate in growing kiwifruit (Actinidia deliciosa, cv Hayward). Sci. Hortic. 2011, 128, 197–205. [Google Scholar] [CrossRef]
- MacRae, E.; Quick, W.P.; Benker, C.; Stitt, M. Carbohydrate metabolism during postharvest ripening in kiwifruit. Planta 1992, 188, 314–323. [Google Scholar] [CrossRef]
- Gomez, M.; Lajolo, F.; Cordenunsi, B. Evolution of soluble sugars during ripening of papaya fruit and its relation to sweet taste. J. Food Sci. 2010, 67, 442–447. [Google Scholar] [CrossRef]
- Basson, C.E.; Groenewald, J.H.; Kossmann, J.; Cronjé, C.; Bauer, R. Sugar and acid-related quality attributes and enzyme activities in strawberry fruits: Invertase is the main sucrose hydrolysing enzyme. Food Chem. 2010, 121, 1156–1162. [Google Scholar] [CrossRef]
- Zhang, C.H.; Yu, M.L.; Ma, R.J.; Shen, Z.J.; Zhang, B.B.; Korir, N.K. Structure, expression profile, and evolution of the sucrose synthase gene family in peach (Prunus persica). Acta Physiol. Plant. 2015, 37, 81. [Google Scholar] [CrossRef]
- Zhu, X.D.; Wang, M.Q.; Li, X.P.; Jiu, S.T.; Wang, C.; Fang, J.G. Genome-wide analysis of the sucrose synthase gene family in grape (Vitis vinifera): Structure, evolution, and expression Profiles. Genes 2017, 8, 111. [Google Scholar] [CrossRef]
- Lv, J.H.; Wang, Y.Z.; Cheng, R.; Wang, G.M.; Zhang, S.L.; Wu, J.; Zhang, H.P. Genome-wide identification and expression analysis of sucrose synthase (SUS) and sucrose phosphate synthase (SPS) gene families in pear. Acta Hortic. Sin. 2018, 45, 421–435. [Google Scholar]
- Tong, X.L.; Wang, Z.Y.; Ma, B.Q.; Zhang, C.X.; Zhu, L.C.; Ma, F.W.; Li, M.J. Structure and expression analysis of the sucrose synthase gene family in apple. J. Integr. Agric. 2018, 17, 847–856. [Google Scholar] [CrossRef] [Green Version]
- He, X.X.; Wei, Y.Y.; Kou, J.Y.; Xu, F.; Chen, Z.H.; Shao, X.F. PpVIN2, an acid invertase gene family member, is sensitive to chilling temperature and affects sucrose metabolism in postharvest peach fruit. Plant Growth Regul. 2018, 86, 169–180. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Y.; Yang, Y.; Yan, Y.Q. Identification of Sucrose Synthase Gene Family in Kiwifruit and Their Expression during Fruit Development. Mol. Plant Breed. 2021. Available online: http://kns.cnki.net/kcms/detail/46.1068.S.20210422.1703.010.html (accessed on 17 April 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| CWIN1 | GCCGAAAGGCTACATCAGTCA | TCACTGCACATAAGCACCACATAC |
| CWIN3 | CAAGTCCAAAACCTAGCCGTG | CAAGCAGTGGTGGGGTCTCT |
| CWIN4 | GGCTAACCTTGAAGAGTACACACC | AGCATCAGAGCACATGAGAACC |
| VIN3 | TTCGGCAGAGAAATGGGCT | GTCATAGAATGTCTTTGATGCGTAG |
| VIN5 | CAAGTCCAAAACCTAGCCGTG | CAAGCAGTGGTGGGGTCTCT |
| NIN1 | ACTTTATCGGTAATGTCGGTCCT | CGGGAGTTGCCAATGACG |
| NIN2 | TGCCGAGAGCCGTTTACTG | AGCATCTTCGCCACCAAATAG |
| NIN3 | GGAATGTCAGCCCTGCGAG | TCAGGAGTTGCCAAAGATGCTA |
| SPS1 | AAGCGGGGACACTGACTACG | GAAAGAGGGTAGGTTCTGTTGGC |
| SPS2 | TTGTCTGAAGGAGAGAAGGGAG | ATTGGAAAAGTTACGCTGGAA |
| SPS5 | CAAAGCCGAGATGAAGAAGATG | CCTCCACCACTGTCAACCCA |
| SS1 | GCATTGCTGATACGAAGGG | CGACTATGATTTCCGCTGGT |
| SS2 | GGGAAAACGGGTTAGAGCAG | AAACACCACGAAGAGCAGGG |
| SS4 | TCAGAGATATTCCAGGCACCG | TCAGAGATATTCCAGGCACCG |
| SS5 | CAAGAATCATCGCAGACGGA | GAGTGAGGGCAAGAAGTGTAAGC |
| Actin | GCTCCACCTGAGAGGAAATAC | CGAAATCCACATCTGTTGAAAG |
| Stages (DAFB) | Vertical Diameter (mm) | Transverse Diameter (mm) | Lateral Diameter (mm) | Single Fruit Weight (g) | Fruit Shape Index |
|---|---|---|---|---|---|
| 25 | 28.77 ± 2.34 i | 15.14 ± 2.9 h | 14.7 ± 2.42 g | 2.320 ± 0.841 j | 1.9 ± 0.18 c |
| 40 | 40.17 ± 3.58 h | 17.3 ± 1.09 g | 16.53 ± 0.65 f | 6.597 ± 1.158 i | 2.32 ± 0.28 a |
| 55 | 41.56 ± 2.65 gh | 19.05 ± 1.56 f | 18.3 ± 1.54 e | 8.538 ± 1.362 h | 2.18 ± 0.13 b |
| 70 | 41.27 ± 2.82 gh | 22.62 ± 3.14 e | 21.52 ± 2.47 d | 10.349 ± 1.362 g | 1.82 ± 0.22 cdef |
| 85 | 42.22 ± 3.48 g | 24.2 ± 3.54 cd | 22.76 ± 3.40 cd | 11.580 ± 2.252 g | 1.74 ± 0.34 fg |
| 100 | 42.59 ± 3.44 fg | 24.82 ± 1.20 bcd | 23.81 ± 1.39 abc | 13.066 ± 2.695 f | 1.72 ± 1.60 g |
| 115 | 44.11 ± 2.71 ef | 25.26 ± 1.69 bc | 24.62 ± 2.52 ab | 15.809 ± 2.223 e | 1.75 ± 0.17 efg |
| 130 | 44.62 ± 4.31 de | 24.91 ± 2.27 bcd | 23.43 ± 1.71 bc | 16.555 ± 2.976 de | 1.79 ± 0.24 defg |
| 145 | 46.46 ± 4.46 cd | 24.94 ± 2.56 bcd | 23.82 ± 1.90 abc | 17.560 ± 3.901 bcd | 1.86 ± 0.29 cde |
| 160 | 47.84 ± 2.84 bc | 26.04 ± 2.52 ab | 24.07 ± 1.37 abc | 18.132 ± 1.196 bc | 1.84 ± 0.26 cdef |
| 175 | 48.5 ± 2.01 b | 25.46 ± 2.32 bc | 23.27 ± 2.32 bc | 18.363 ± 2.832 b | 1.91 ± 0.17 cd |
| 190 | 51.24 ± 4.91 a | 27.18 ± 1.94 a | 25.16 ± 3.29 a | 21.982 ± 5.741 a | 1.89 ± 0.13 cd |
| Soft-ripening stage | 49.05 ± 3.20 b | 25.8 ± 2.29 bc | 23.14 ± 2.11 c | 19.695 ± 2.702 ab | 1.93 ± 0.13 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, J.; Wu, M.; Jiao, X.; Chen, S.; Jia, D.; Xu, X.; Huang, C. Dynamic Changes of Fruit Physiological Quality and Sugar Components during Fruit Growth and Development of Actinidia eriantha. Horticulturae 2022, 8, 529. https://doi.org/10.3390/horticulturae8060529
Tao J, Wu M, Jiao X, Chen S, Jia D, Xu X, Huang C. Dynamic Changes of Fruit Physiological Quality and Sugar Components during Fruit Growth and Development of Actinidia eriantha. Horticulturae. 2022; 8(6):529. https://doi.org/10.3390/horticulturae8060529
Chicago/Turabian StyleTao, Junjie, Mengting Wu, Xudong Jiao, Shuangshuang Chen, Dongfeng Jia, Xiaobiao Xu, and Chunhui Huang. 2022. "Dynamic Changes of Fruit Physiological Quality and Sugar Components during Fruit Growth and Development of Actinidia eriantha" Horticulturae 8, no. 6: 529. https://doi.org/10.3390/horticulturae8060529