
Comparative Metabolomic and Transcriptomic Analysis Reveals That Variations in Flavonoids Determine the Colors of Different Rambutan Cultivars
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Sampling
2.2. Physiological Index Measurement
2.3. Metabolomics Analysis
2.4. RNA Extraction and Sequencing
2.5. cDNA Synthesis and Quantitative Real-Time PCR (Q-PCR)
2.6. Weighted Gene Co-Expression Network Analysis (WGCNA)
2.7. Statistical Analysis
3. Results and Analysis
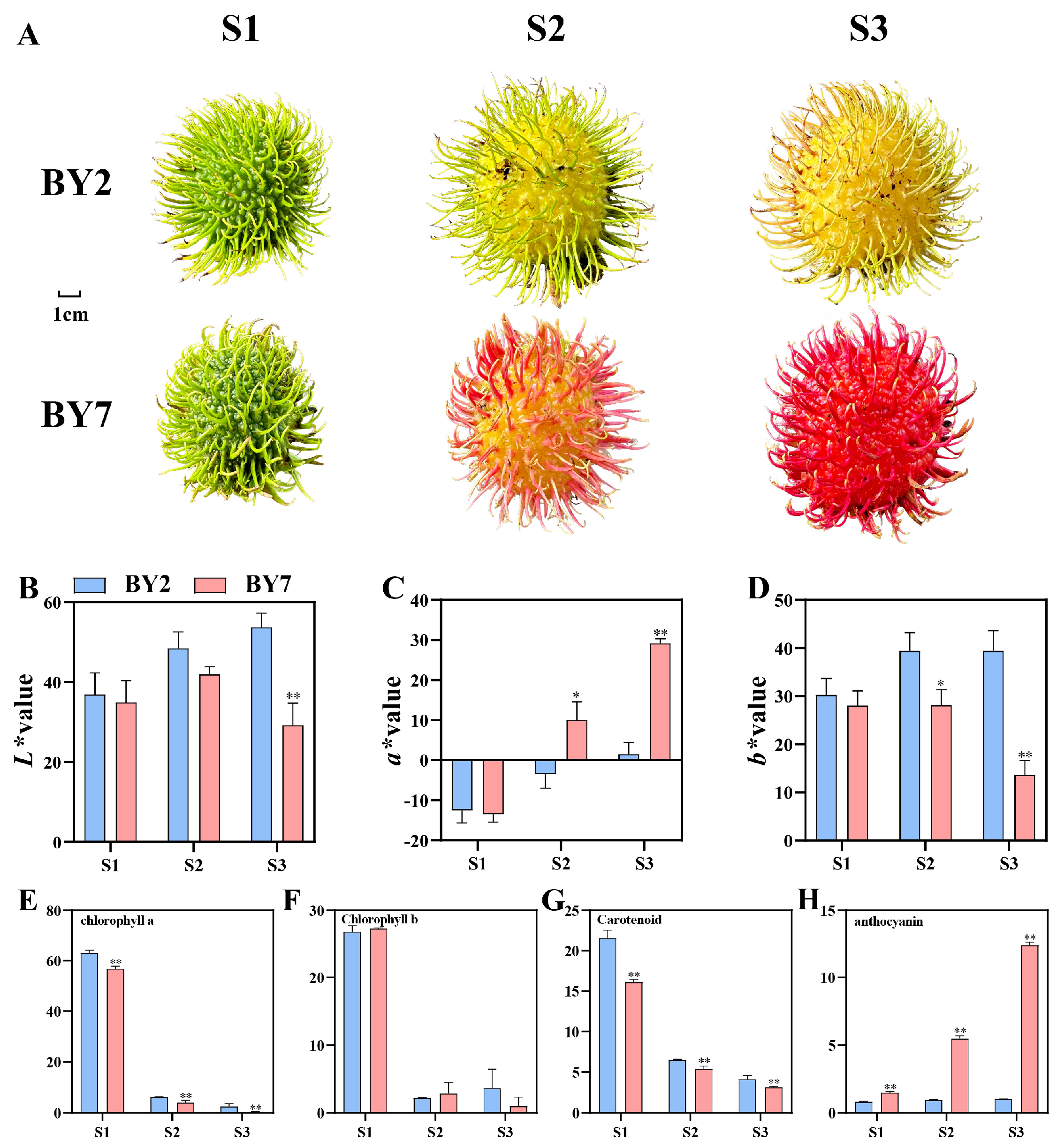
3.1. Basic Analysis of Two Differently Colored Rambutan Cultivars
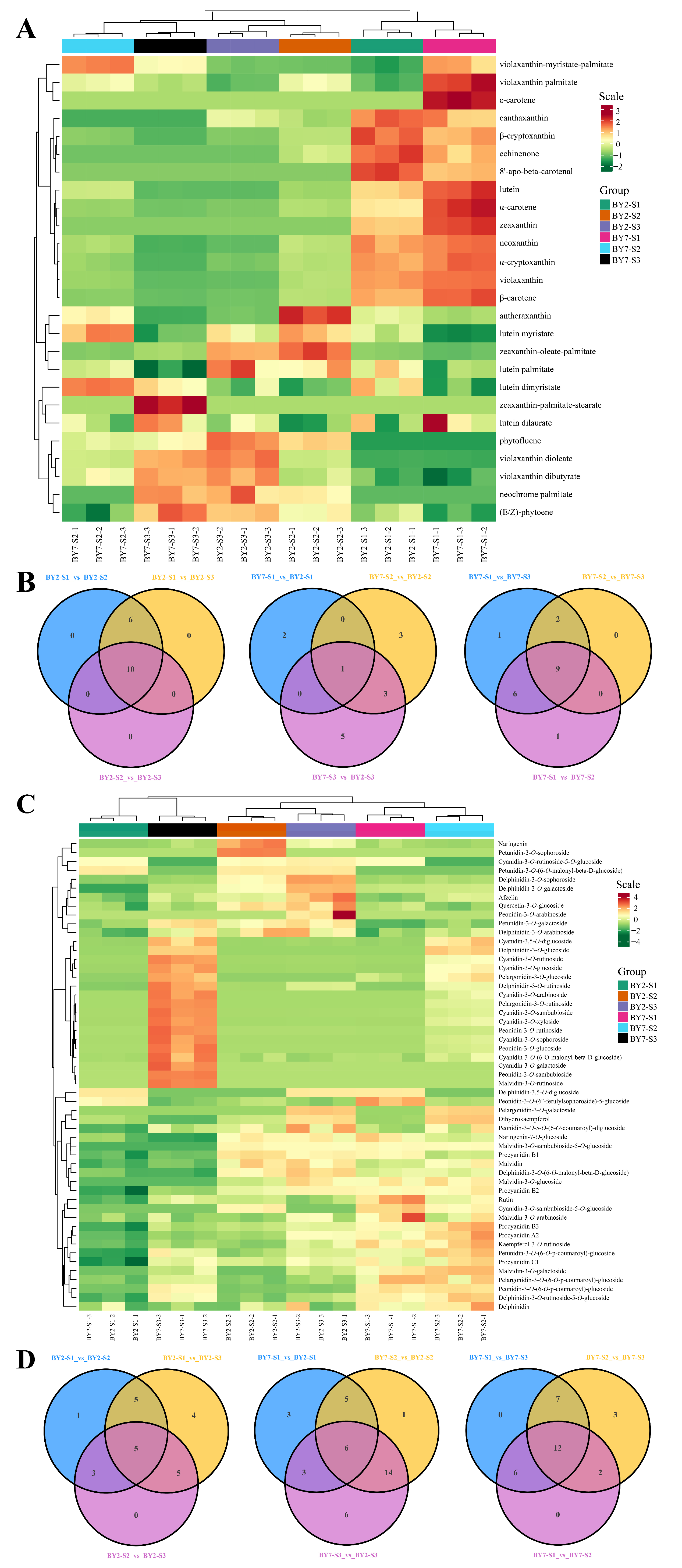
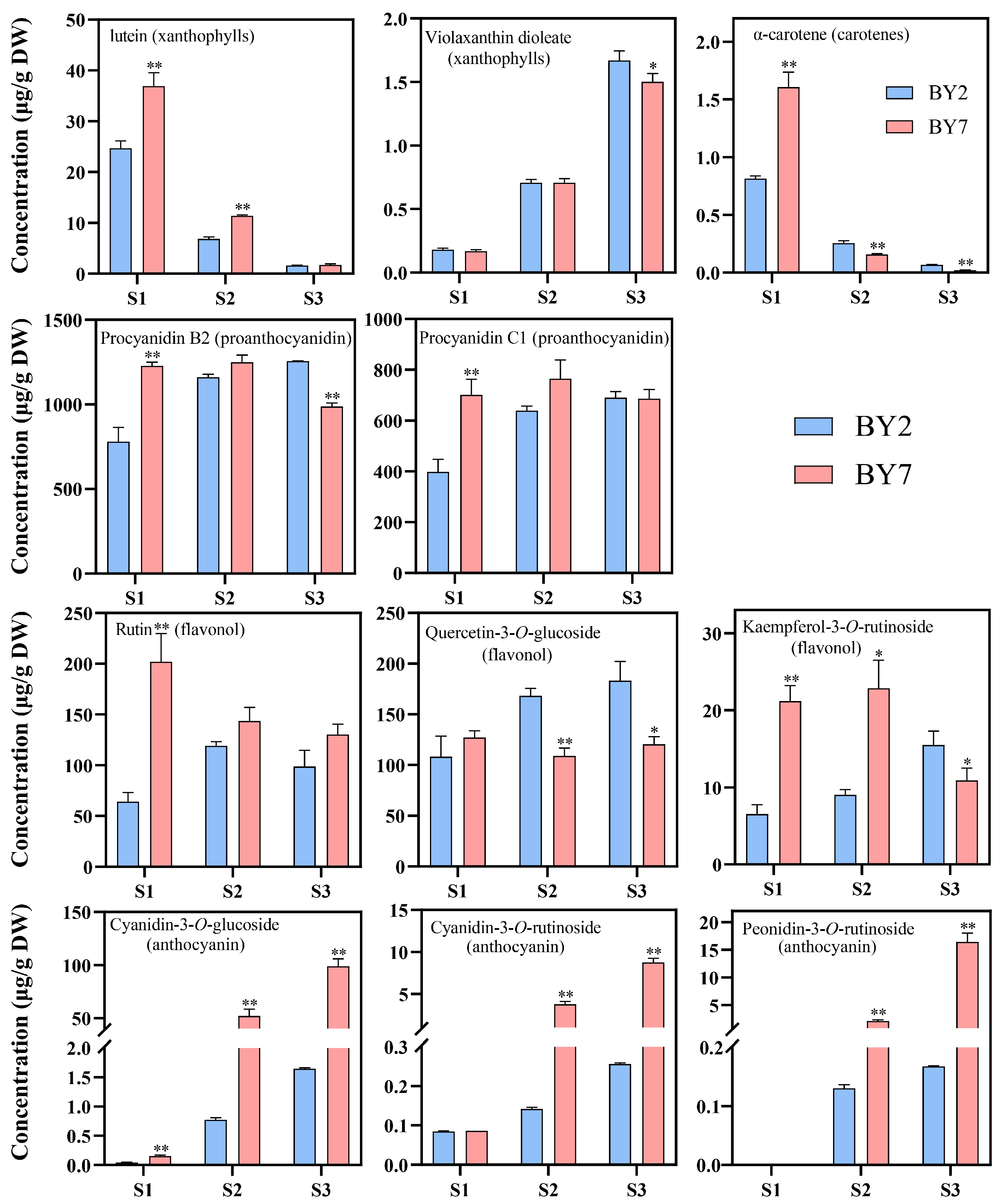
3.2. Carotenoid Accumulation in ‘BY7’ and ‘BY2’
3.3. Flavonoids Accumulation in the ‘BY7’ and ‘BY2’ Rambutan Cultivars
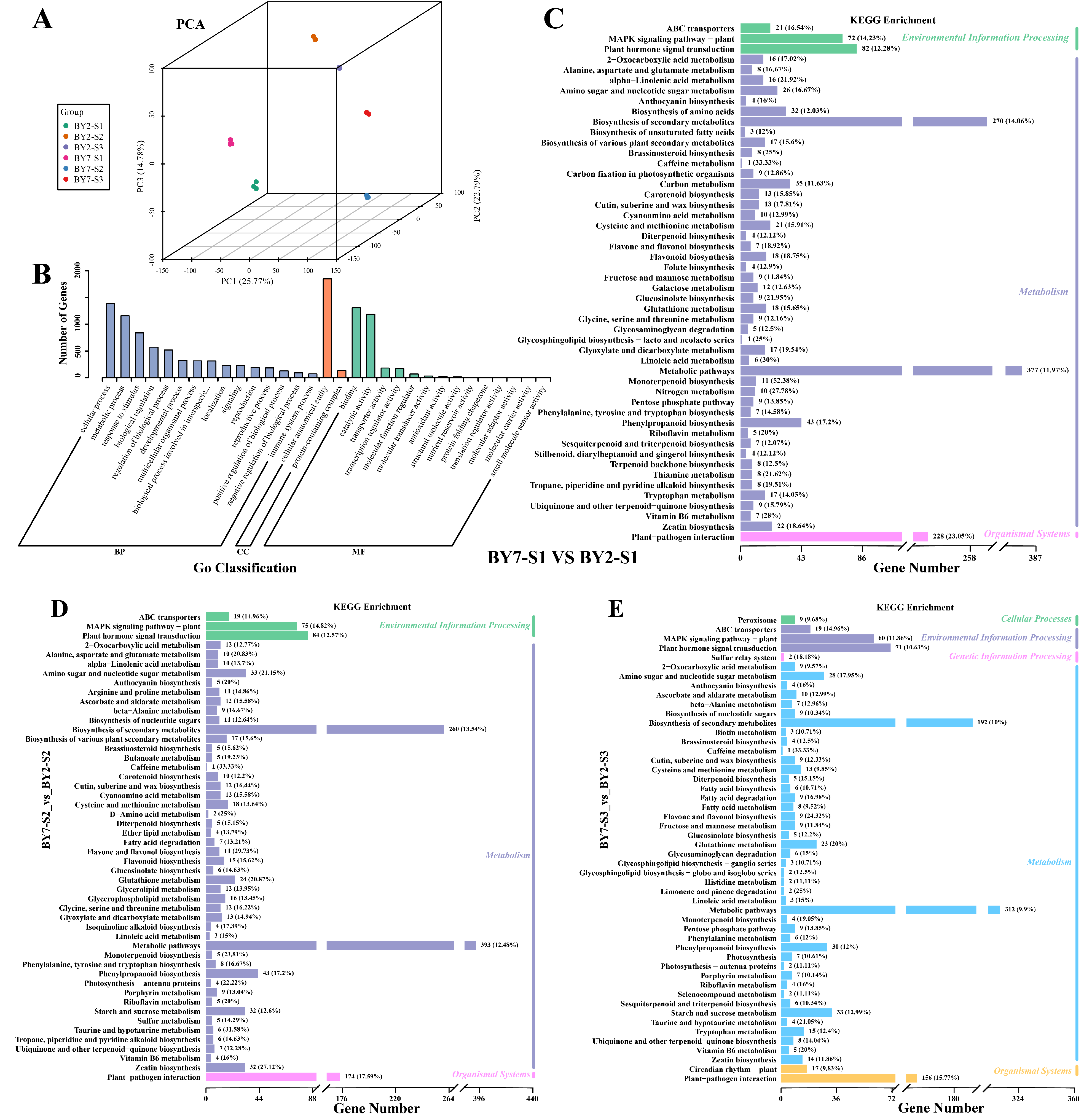
3.4. RNA Seq
3.5. Dynamics of DEG Expression
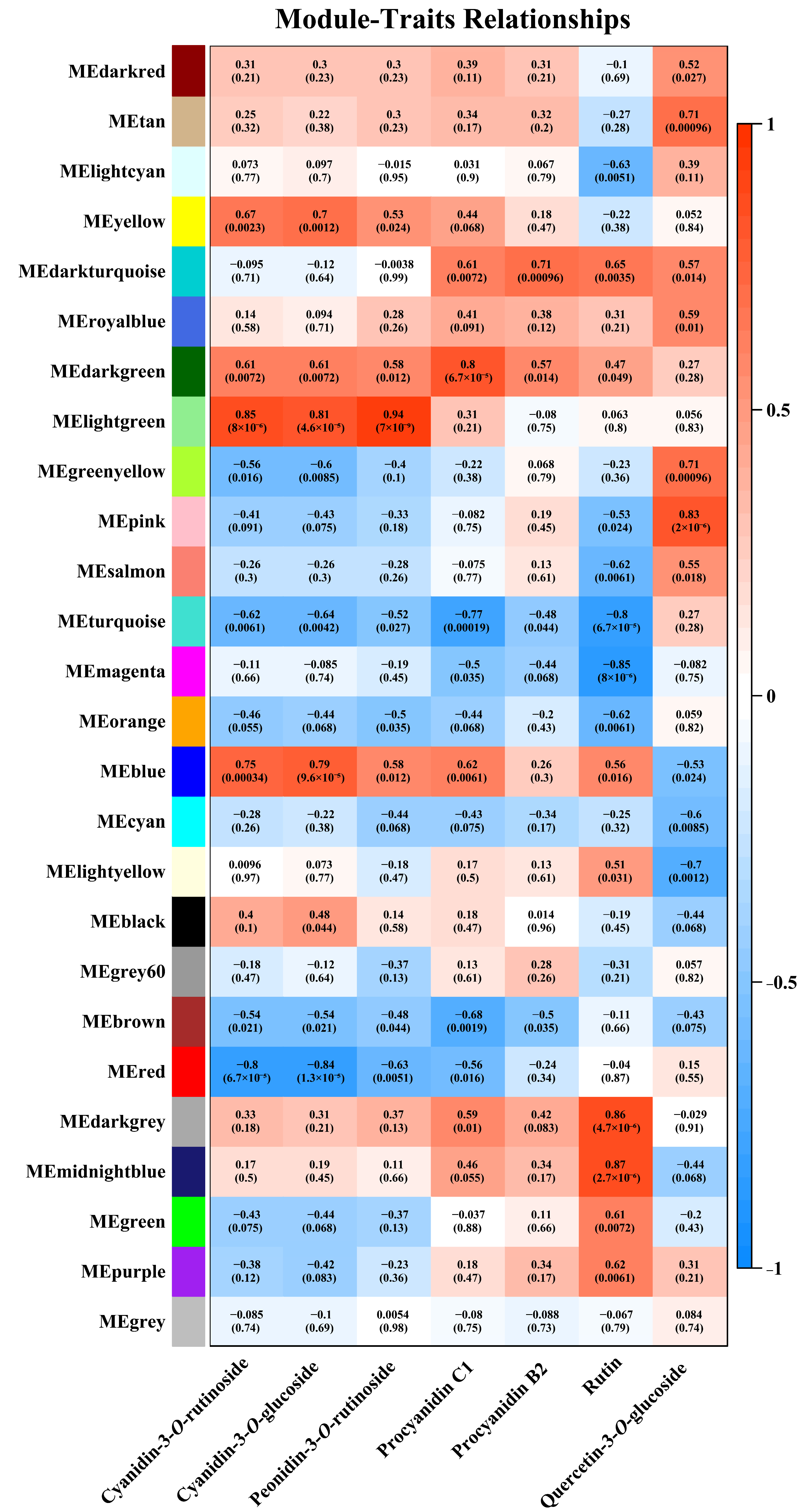
3.6. Flavonoid-Related DEGs Identified via WGCNA
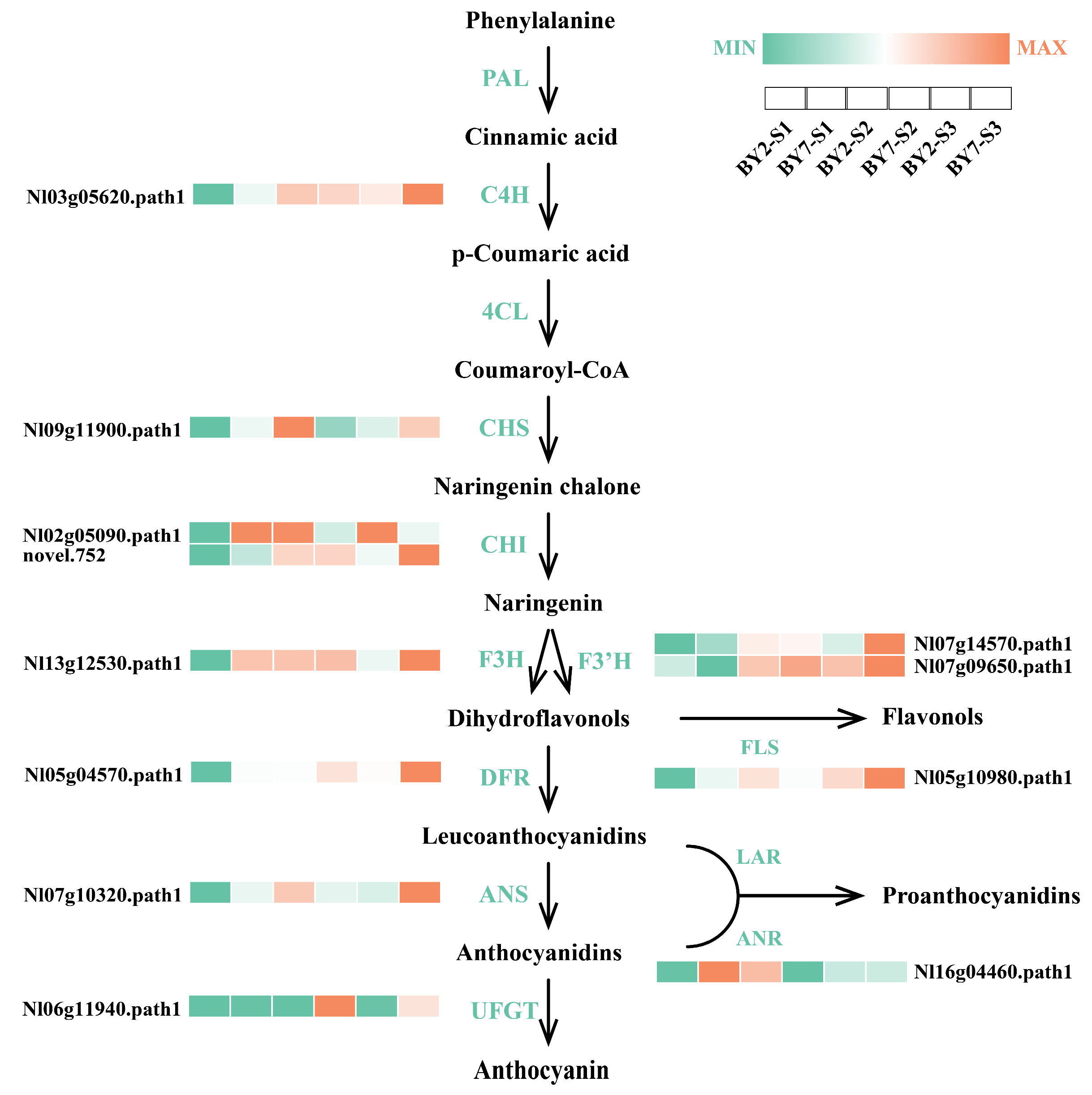
3.7. Structural Genes Associated with Flavonoid Biosynthesis
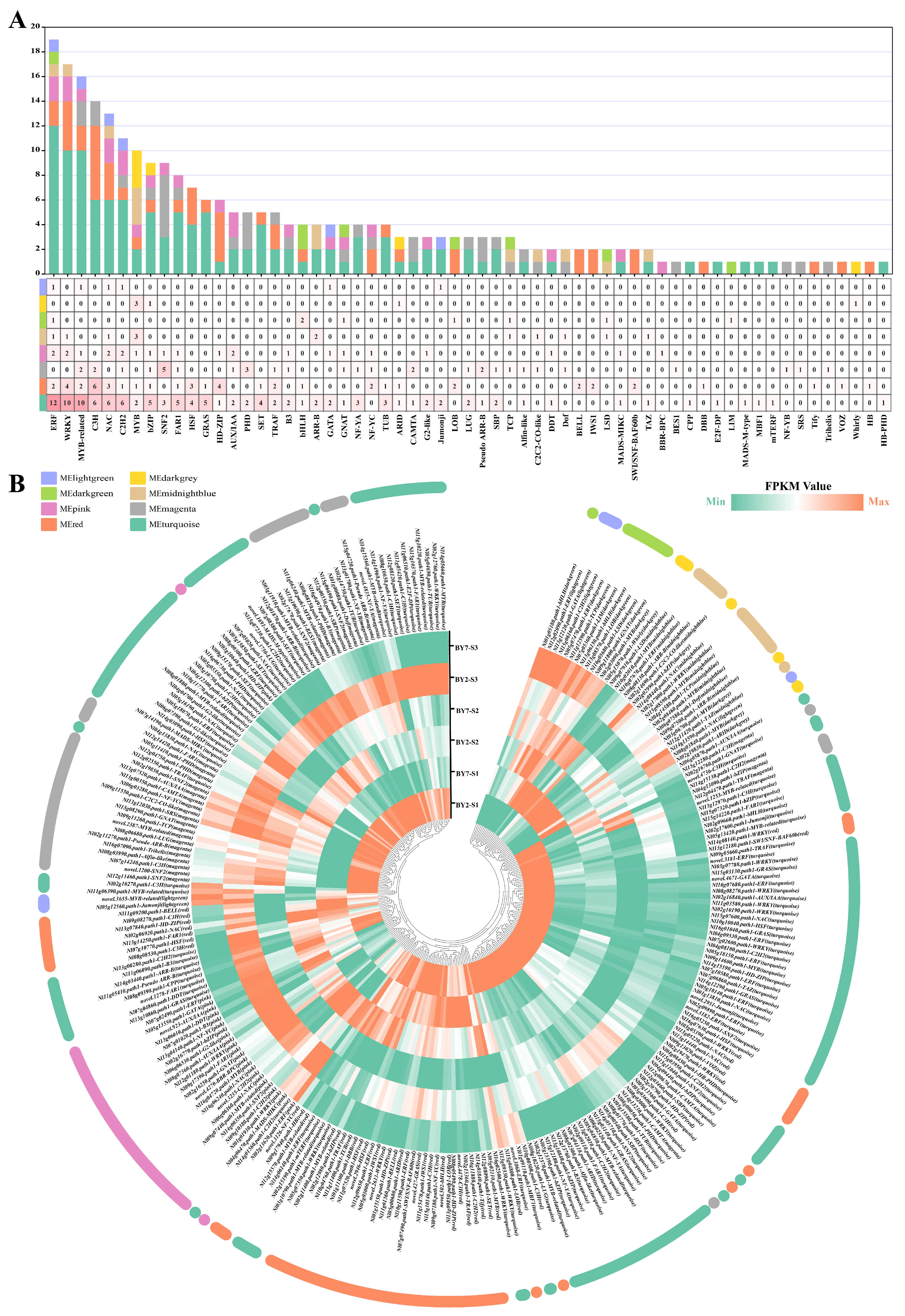
3.8. Regulatory Genes
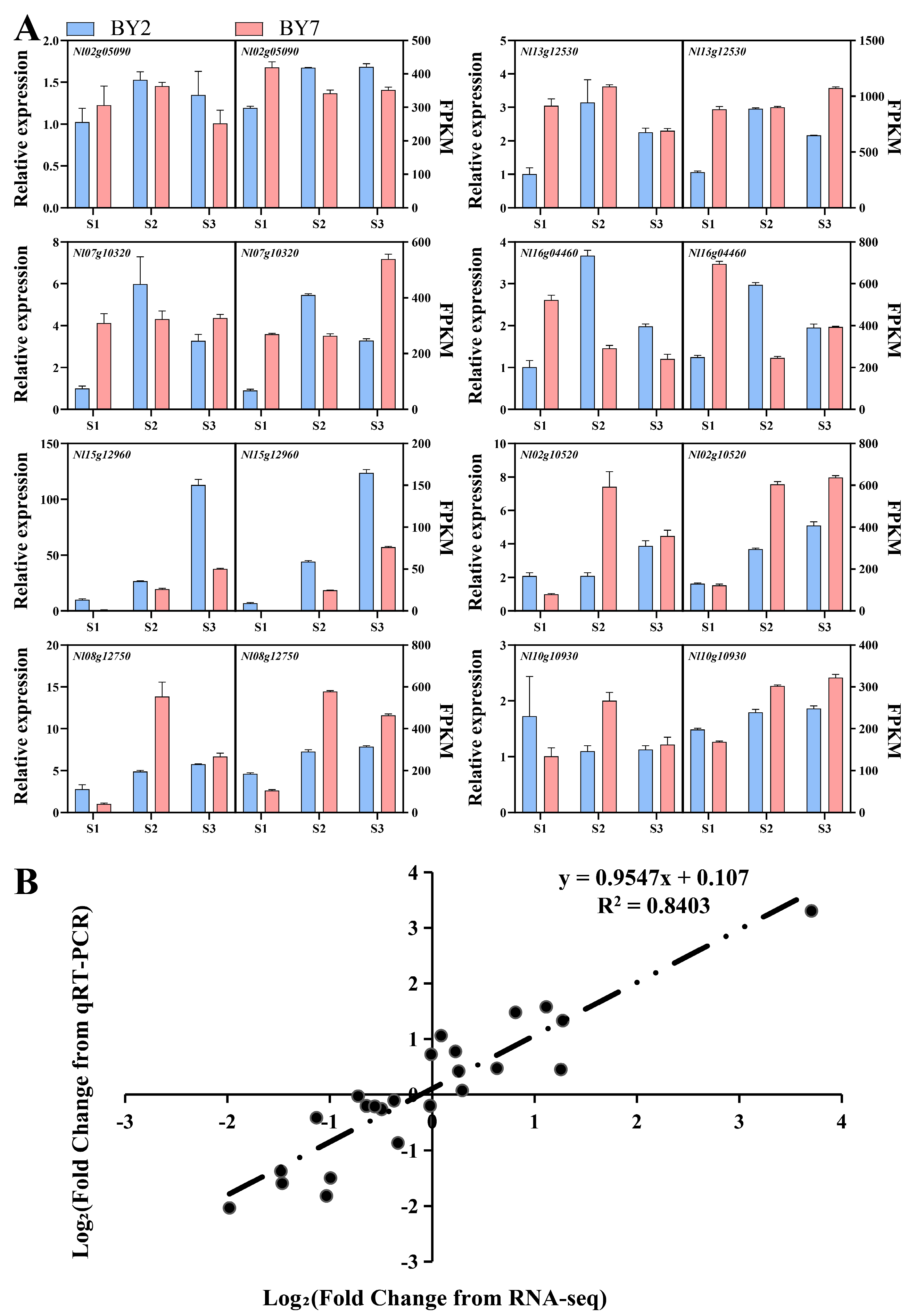
3.9. qPCR Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid Metabolism and Regulation in Horticultural Crops. Hort. Res. 2015, 2, 15036. [Google Scholar] [CrossRef]
- Cuthill, I.C.; Allen, W.L.; Arbuckle, K.; Caspers, B.; Chaplin, G.; Hauber, M.E.; Hill, G.E.; Jablonski, N.G.; Jiggins, C.D.; Kelber, A.; et al. The Biology of Color. Science 2017, 357, eaan0221. [Google Scholar] [CrossRef]
- Shi, B.; Wu, H.; Zheng, B.; Qian, M.; Gao, A.; Zhou, K. Analysis of Light-Independent Anthocyanin Accumulation in Mango (Mangifera indica L.). Horticulturae 2021, 7, 423. [Google Scholar] [CrossRef]
- Yue, P.; Jiang, Z.; Sun, Q.; Wei, R.; Yin, Y.; Xie, Z.; Larkin, R.M.; Ye, J.; Chai, L.; Deng, X. Jasmonate Activates a CsMPK6-CsMYC2 Module That Regulates the Expression of β-Citraurin Biosynthetic Genes and Fruit Coloration in Orange (Citrus sinensis). Plant Cell 2023, 35, 1167–1185. [Google Scholar] [CrossRef]
- Ni, J.; Zhao, Y.; Tao, R.; Yin, L.; Gao, L.; Strid, Å.; Qian, M.; Li, J.; Li, Y.; Shen, J.; et al. Ethylene Mediates the Branching of the Jasmonate-Induced Flavonoid Biosynthesis Pathway by Suppressing Anthocyanin Biosynthesis in Red Chinese Pear Fruits. Plant Biotechnol. J. 2020, 18, 1223–1240. [Google Scholar] [CrossRef]
- Chen, J.; Xie, F.; Cui, Y.; Chen, C.; Lu, W.; Hu, X.; Hua, Q.; Zhao, J.; Wu, Z.; Gao, D.; et al. A Chromosome-Scale Genome Sequence of Pitaya (Hylocereus undatus) Provides Novel Insights into the Genome Evolution and Regulation of Betalain Biosynthesis. Hort. Res. 2021, 8, 164. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in Flavonoid Research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Schweiggert, R.M.; Carle, R. Carotenoid Deposition in Plant and Animal Foods and Its Impact on Bioavailability. Crit. Rev. Food Sci. Nutr. 2017, 57, 1807–1830. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Ortega, E.; Hornero-Méndez, D. Carotenoids in Cereals: An Ancient Resource with Present and Future Applications. Phytochem. Rev. 2015, 14, 873–890. [Google Scholar] [CrossRef]
- Sarma, A.D.; Sharma, R. Anthocyanin-DNA Copigmentation Complex: Mutual Protection against Oxidative Damage. Phytochemistry 1999, 52, 1313–1318. [Google Scholar] [CrossRef]
- Lampila, P.; van Lieshout, M.; Gremmen, B.; Lähteenmäki, L. Consumer Attitudes towards Enhanced Flavonoid Content in Fruit. Food Res. Int. 2009, 42, 122–129. [Google Scholar] [CrossRef]
- Fang, T.; Chen, J.; Lin, Q.; Zhong, Y.; Duan, Y.; Bi, J. Phenolic Profiling Reveals the Metabolite Basis of Flesh Colour and Fresh-Cut Browning in Apple Fruit. Int. J. Food Sci. Technol. 2022, 57, 2257–2266. [Google Scholar] [CrossRef]
- Moise, A.R.; Al-Babili, S.; Wurtzel, E.T. Mechanistic Aspects of Carotenoid Biosynthesis. Chem. Rev. 2014, 114, 164–193. [Google Scholar] [CrossRef]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The Flavonoid Biosynthetic Pathway in Arabidopsis: Structural and Genetic Diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef]
- Giuliano, G. Plant Carotenoids: Genomics Meets Multi-Gene Engineering. Curr. Opin. Plant Biol. 2014, 19, 111–117. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB Transcription Factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Ye, J.; Hu, T.; Yang, C.; Li, H.; Yang, M.; Ijaz, R.; Ye, Z.; Zhang, Y. Transcriptome Profiling of Tomato Fruit Development Reveals Transcription Factors Associated with Ascorbic Acid, Carotenoid and Flavonoid Biosynthesis. PLoS ONE 2015, 10, e0130885. [Google Scholar] [CrossRef]
- Sirisompong, W.; Jirapakkul, W.; Klinkesorn, U. Response Surface Optimization and Characteristics of Rambutan (Nephelium lappaceum L.) Kernel Fat by Hexane Extraction. LWT Food Sci. Technol. 2011, 44, 1946–1951. [Google Scholar] [CrossRef]
- Chai, K.F.; Mohd Adzahan, N.; Karim, R.; Rukayadi, Y.; Mohd Ghazali, H. Selected Physicochemical Properties of Registered Clones and Wild Types Rambutan (Nephelium lappaceum L.) Fruits and Their Potentials in Food Products. Sains Malays. 2018, 47, 1483–1490. [Google Scholar] [CrossRef]
- O’Hare, T.J. Postharvest Physiology and Storage of Rambutan. Postharvest Biol. Technol. 1995, 6, 189–199. [Google Scholar] [CrossRef]
- Chai, K.F.; Mohd Adzahan, N.; Karim, R.; Rukayadi, Y.; Mohd Ghazali, H. Physicochemical Properties of Rambutan (Nephelium lappaceum L.) Seed during Natural Fermentation of the Whole Peeled Fruit. Int. Food Res. J. 2020, 27, 397–407. [Google Scholar]
- Lee, P.; Tan, R.; Yu, B.; Curran, P.; Liu, S. Sugars, Organic Acids, and Phenolic Acids of Exotic Seasonable Tropical Fruits. Food Sci. Nutr. 2013, 43, 267–276. [Google Scholar] [CrossRef]
- Deng, H.; Wu, G.; Zhang, R.; Yin, Q.; Xu, B.; Zhou, L.; Chen, Z. Comparative Nutritional and Metabolic Analysis Reveals the Taste Variations during Yellow Rambutan Fruit Maturation. Food Chem. X 2023, 17, 100580. [Google Scholar] [CrossRef]
- Ding, R.; Che, X.; Shen, Z.; Zhang, Y. Metabolome and Transcriptome Profiling Provide Insights into Green Apple Peel Reveals Light- and UV-B-Responsive Pathway in Anthocyanins Accumulation. BMC Plant Biol. 2021, 21, 351. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.-Y.; Dillies, M.-A. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a Toolkit for Biologists Integrating Various HTS-Data Handling Tools with a User-Friendly Interface. bioRxiv 2018. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants in Plants: Location and Functional Significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red Colouration in Apple Fruit Is Due to the Activity of the MYB Transcription Factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef]
- Ban, Y.; Kondo, S.; Ubi, B.E.; Honda, C.; Bessho, H.; Moriguchi, T. UDP-Sugar Biosynthetic Pathway: Contribution to Cyanidin 3-Galactoside Biosynthesis in Apple Skin. Planta 2009, 230, 871–881. [Google Scholar] [CrossRef]
- Yi, D.; Zhang, H.; Lai, B.; Liu, L.; Pan, X.; Ma, Z.; Wang, Y.; Xie, J.; Shi, S.; Wei, Y. Integrative Analysis of the Coloring Mechanism of Red Longan Pericarp through Metabolome and Transcriptome Analyses. J. Agric. Food Chem. 2021, 69, 1806–1815. [Google Scholar] [CrossRef]
- Aida, R.; Yoshida, K.; Kondo, T.; Kishimoto, S.; Shibata, M. Copigmentation Gives Bluer Flowers on Transgenic Torenia Plants with the Antisense Dihydroflavonol-4-Reductase Gene. Plant Sci. 2000, 160, 49–56. [Google Scholar] [CrossRef]
- Zhou, X.-W.; Fan, Z.-Q.; Chen, Y.; Zhu, Y.-L.; Li, J.-Y.; Yin, H.-F. Functional Analyses of a Flavonol Synthase–like Gene from Camellia Nitidissima Reveal Its Roles in Flavonoid Metabolism during Floral Pigmentation. J. Biosci. 2013, 38, 593–604. [Google Scholar] [CrossRef]
- Kang, Y.; Li, Y.; Zhang, T.; Wang, P.; Liu, W.; Zhang, Z.; Yu, W.; Wang, J.; Wang, J.; Zhou, Y. Integrated Metabolome, Full-Length Sequencing, and Transcriptome Analyses Unveil the Molecular Mechanisms of Color Formation of the Canary Yellow and Red Bracts of Bougainvillea × Buttiana ‘Chitra’. Plant J. 2023, 116, 1441–1461. [Google Scholar] [CrossRef]
- Yuan, Y.-W.; Rebocho, A.B.; Sagawa, J.M.; Stanley, L.E.; Bradshaw, H.D. Competition between Anthocyanin and Flavonol Biosynthesis Produces Spatial Pattern Variation of Floral Pigments between Mimulus Species. Proc. Natl. Acad. Sci. USA 2016, 113, 2448–2453. [Google Scholar] [CrossRef]
- Shen, J.; Zou, Z.; Zhang, X.; Zhou, L.; Wang, Y.; Fang, W.; Zhu, X. Metabolic Analyses Reveal Different Mechanisms of Leaf Color Change in Two Purple-Leaf Tea Plant (Camellia sinensis L.) Cultivars. Hort. Res. 2018, 5, 7. [Google Scholar] [CrossRef]
- Kanzaki, S.; Ichihi, A.; Tanaka, Y.; Fujishige, S.; Koeda, S.; Shimizu, K. The R2R3-MYB Transcription Factor MiMYB1 Regulates Light Dependent Red Coloration of ‘Irwin’ Mango Fruit Skin. Sci. Hortic. 2020, 272, 109567. [Google Scholar] [CrossRef]
- Zhu, W.; Wu, H.; Yang, C.; Shi, B.; Zheng, B.; Ma, X.; Zhou, K.; Qian, M. Postharvest Light-Induced Flavonoids Accumulation in Mango (Mangifera indica L.) Peel Is Associated with the up-Regulation of Flavonoids-Related and Light Signal Pathway Genes. Front. Plant Sci. 2023, 14, 1136281. [Google Scholar] [CrossRef]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 Synergistically Specify the Expression of BANYULS and Proanthocyanidin Biosynthesis in Arabidopsis thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin Synthesis and Expression of Genes Encoding Leucoanthocyanidin Reductase and Anthocyanidin Reductase in Developing Grape Berries and Grapevine Leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef]
- Han, Y.; Vimolmangkang, S.; Soria-Guerra, R.E.; Korban, S.S. Introduction of Apple ANR Genes into Tobacco Inhibits Expression of Both CHI and DFR Genes in Flowers, Leading to Loss of Anthocyanin. J. Exp. Bot. 2012, 63, 2437–2447. [Google Scholar] [CrossRef]
- Ji, X.; Li, Z.; Zhang, M.; Lang, S.; Song, X. ChMYB1-ChbHLH42-ChTTG1 Module Regulates Abscisic Acid-Induced Anthocyanin Biosynthesis in Cerasus humilis. Hortic Plant J. 2023, 10, 51–65. [Google Scholar] [CrossRef]
- Heppel, S.C.; Jaffé, F.W.; Takos, A.M.; Schellmann, S.; Rausch, T.; Walker, A.R.; Bogs, J. Identification of Key Amino Acids for the Evolution of Promoter Target Specificity of Anthocyanin and Proanthocyanidin Regulating MYB Factors. Plant Mol. 2013, 82, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Ming, M.; Allan, A.C.; Gu, C.; Li, L.; Wu, X.; Wang, R.; Chang, Y.; Qi, K.; Zhang, S.; et al. Map-Based Cloning of the Pear Gene MYB114 Identifies an Interaction with Other Transcription Factors to Coordinately Regulate Fruit Anthocyanin Biosynthesis. Plant J. 2017, 92, 437–451. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Zhang, R.; Jiang, S.; Wang, H.; Ming, F. The MYB Transcription Factor RcMYB1 Plays a Central Role in Rose Anthocyanin Biosynthesis. Hort. Res. 2023, 10, uhad080. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Yue, M.; Liu, Y.; Zhang, N.; Lin, Y.; Zhang, Y.; Wang, Y.; Li, M.; Luo, Y.; Zhang, Y.; et al. A Novel R2R3-MYB Transcription Factor FaMYB5 Positively Regulates Anthocyanin and Proanthocyanidin Biosynthesis in Cultivated Strawberries (Fragaria × Ananassa). Plant Biotechnol. J. 2023, 21, 1140–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xu, H.; Jiang, S.; Zhang, Z.; Lu, N.; Qiu, H.; Qu, C.; Wang, Y.; Wu, S.; Chen, X. MYB12 and MYB22 Play Essential Roles in Proanthocyanidin and Flavonol Synthesis in Red-Fleshed Apple (Malus sieversii f. Niedzwetzkyana). Plant J. 2017, 90, 276–292. [Google Scholar] [CrossRef] [PubMed]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis Transcription Factor MYB12 Is a Flavonol-Specific Regulator of Phenylpropanoid Biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Yu, W.; Gao, Y.; Ni, J.; Yin, L.; Zhang, X.; Li, H.; Wang, D.; Bai, S.; Teng, Y. Light-Induced Basic/Helix-Loop-Helix64 Enhances Anthocyanin Biosynthesis and Undergoes CONSTITUTIVELY PHOTOMORPHOGENIC1-Mediated Degradation in Pear. Plant Physiol. 2020, 184, 1684–1701. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Premathilake, A.T.; Gao, Y.; Yu, W.; Tao, R.; Teng, Y.; Bai, S. Ethylene-Activated PpERF105 Induces the Expression of the Repressor-Type R2R3-MYB Gene PpMYB140 to Inhibit Anthocyanin Biosynthesis in Red Pear Fruit. Plant J. 2021, 105, 167–181. [Google Scholar] [CrossRef]
- Rajput, R.; Naik, J.; Stracke, R.; Pandey, A. Interplay between R2R3 MYB-Type Activators and Repressors Regulates Proanthocyanidin Biosynthesis in Banana (Musa acuminata). New Phytol. 2022, 236, 1108–1127. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Zhu, M.; Chen, S.; Li, C.; Zhang, Q.; Gao, L.; Liu, X.; Shen, S.; Fu, F.; Xu, X.; et al. BnbHLH92a Negatively Regulates Anthocyanin and Proanthocyanidin Biosynthesis in Brassica napus. Crop J. 2023, 11, 374–385. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, H.; Wang, N.; Jiang, S.; Fang, H.; Zhang, Z.; Yang, G.; Wang, Y.; Su, M.; Xu, L.; et al. The Ethylene Response Factor MdERF1B Regulates Anthocyanin and Proanthocyanidin Biosynthesis in Apple. Plant Mol. Biol. 2018, 98, 205–218. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, X.; Gong, Q.; Cao, J.; Shen, W.; Yin, X.; Grierson, D.; Zhang, B.; Xu, C.; Li, X.; et al. Three AP2/ERF Family Members Modulate Flavonoid Synthesis by Regulating Type IV Chalcone Isomerase in Citrus. Plant Biotechnol. J. 2021, 19, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wang, S.; Yu, W.; Liao, Y.; Pan, C.; Zhang, M.; Tao, R.; Wei, J.; Gao, Y.; Wang, D.; et al. The Ethylene-Responsive Transcription Factor PpERF9 Represses PpRAP2.4 and PpMYB114 via Histone Deacetylation to Inhibit Anthocyanin Biosynthesis in Pear. Plant Cell 2023, 35, 2271–2292. [Google Scholar] [CrossRef]
- Li, C.; Wu, J.; Hu, K.-D.; Wei, S.-W.; Sun, H.-Y.; Hu, L.-Y.; Han, Z.; Yao, G.-F.; Zhang, H. PyWRKY26 and PybHLH3 Cotargeted the PyMYB114 Promoter to Regulate Anthocyanin Biosynthesis and Transport in Red-Skinned Pears. Hort. Res. 2020, 7, 37. [Google Scholar] [CrossRef]
- Grunewald, W.; De Smet, I.; Lewis, D.R.; Löfke, C.; Jansen, L.; Goeminne, G.; Vanden Bossche, R.; Karimi, M.; De Rybel, B.; Vanholme, B.; et al. Transcription Factor WRKY23 Assists Auxin Distribution Patterns during Arabidopsis Root Development through Local Control on Flavonol Biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 1554–1559. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Fang, J.; Yin, W.; Yan, X.; Tu, M.; Liu, H.; Zhang, Z.; Li, Z.; Gao, M.; et al. VqWRKY56 Interacts with VqbZIPC22 in Grapevine to Promote Proanthocyanidin Biosynthesis and Increase Resistance to Powdery Mildew. New Phytol. 2023, 237, 1856–1875. [Google Scholar] [CrossRef]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-Related Genes of the Kyoho Grape (Vitis labruscana) Regulate Anthocyanin Biosynthesis. Planta 2002, 215, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Lee, H. MYB-Related Transcription Factors Function as Regulators of the Circadian Clock and Anthocyanin Biosynthesis in Arabidopsis. Plant Signal. Behav. 2016, 11, e1139278. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Jiang, S.; Zhang, T.; Xu, H.; Fang, H.; Zhang, J.; Su, M.; Wang, Y.; Zhang, Z.; Wang, N.; et al. Apple NAC Transcription Factor MdNAC52 Regulates Biosynthesis of Anthocyanin and Proanthocyanidin through MdMYB9 and MdMYB11. Plant Sci. 2019, 289, 110286. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Pei, J.; Yan, X.; Cui, X.; Tsuruta, M.; Liu, Y.; Lian, C. A Poplar B-Box Protein PtrBBX23 Modulates the Accumulation of Anthocyanins and Proanthocyanidins in Response to High Light. Plant Cell Environ. 2021, 44, 3015–3033. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Clean Base (G) | Error Rate (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| BY2-S1-1 | 48,214,978 | 46,952,488 | 7.04 | 0.03 | 97.1 | 92.16 | 45.45 |
| BY2-S1-2 | 45,776,308 | 44,553,686 | 6.68 | 0.03 | 97.05 | 92.06 | 45.38 |
| BY2-S1-3 | 47,594,682 | 46,284,140 | 6.94 | 0.03 | 96.89 | 91.76 | 45.39 |
| BY2-S2-1 | 50,542,420 | 49,255,750 | 7.39 | 0.03 | 96.68 | 91.3 | 45.59 |
| BY2-S2-2 | 53,658,274 | 52,363,548 | 7.85 | 0.03 | 96.82 | 91.59 | 45.6 |
| BY2-S2-3 | 55,737,910 | 54,482,616 | 8.17 | 0.03 | 96.84 | 91.63 | 45.55 |
| BY2-S3-1 | 45,602,194 | 44,623,996 | 6.69 | 0.03 | 96.97 | 91.82 | 45.38 |
| BY2-S3-2 | 45,403,508 | 44,327,470 | 6.65 | 0.03 | 97.43 | 92.73 | 45.4 |
| BY2-S3-3 | 45,686,916 | 44,736,964 | 6.71 | 0.03 | 97.24 | 92.34 | 45.48 |
| BY7-S1-1 | 60,699,674 | 58,961,646 | 8.84 | 0.03 | 96.98 | 91.94 | 45.88 |
| BY7-S1-2 | 74,150,954 | 71,883,730 | 10.78 | 0.03 | 97.21 | 92.44 | 45.83 |
| BY7-S1-3 | 63,322,594 | 61,633,524 | 9.25 | 0.03 | 96.99 | 91.94 | 45.88 |
| BY7-S2-1 | 67,488,394 | 65,868,242 | 9.88 | 0.03 | 97.18 | 92.35 | 45.51 |
| BY7-S2-2 | 58,550,090 | 57,328,064 | 8.6 | 0.03 | 97.11 | 92.1 | 45.6 |
| BY7-S2-3 | 56,373,226 | 55,146,590 | 8.27 | 0.03 | 97.25 | 92.41 | 45.53 |
| BY7-S3-1 | 59,593,582 | 58,351,826 | 8.75 | 0.03 | 97.21 | 92.33 | 45.43 |
| BY7-S3-2 | 47,865,312 | 46,842,628 | 7.03 | 0.03 | 97.22 | 92.42 | 45.42 |
| BY7-S3-3 | 62,324,624 | 60,877,868 | 9.13 | 0.03 | 97.14 | 92.2 | 45.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhu, W.; Yang, C.; Li, M.; Feng, S.; Tang, L.; Yang, C.; Cui, Z. Comparative Metabolomic and Transcriptomic Analysis Reveals That Variations in Flavonoids Determine the Colors of Different Rambutan Cultivars. Horticulturae 2024, 10, 263. https://doi.org/10.3390/horticulturae10030263
Wang J, Zhu W, Yang C, Li M, Feng S, Tang L, Yang C, Cui Z. Comparative Metabolomic and Transcriptomic Analysis Reveals That Variations in Flavonoids Determine the Colors of Different Rambutan Cultivars. Horticulturae. 2024; 10(3):263. https://doi.org/10.3390/horticulturae10030263
Chicago/Turabian StyleWang, Jiaqi, Wencan Zhu, Chengkun Yang, Maofu Li, Shun Feng, Lizhu Tang, Chengchao Yang, and Zhifu Cui. 2024. "Comparative Metabolomic and Transcriptomic Analysis Reveals That Variations in Flavonoids Determine the Colors of Different Rambutan Cultivars" Horticulturae 10, no. 3: 263. https://doi.org/10.3390/horticulturae10030263