
Advance in Heterologous Expression of Biomass-Degrading Auxiliary Activity 10 Family of Lytic Polysaccharide Monooxygenases
 ,
,
Abstract
:1. Introduction
2. Selection of Expression Host and Vector
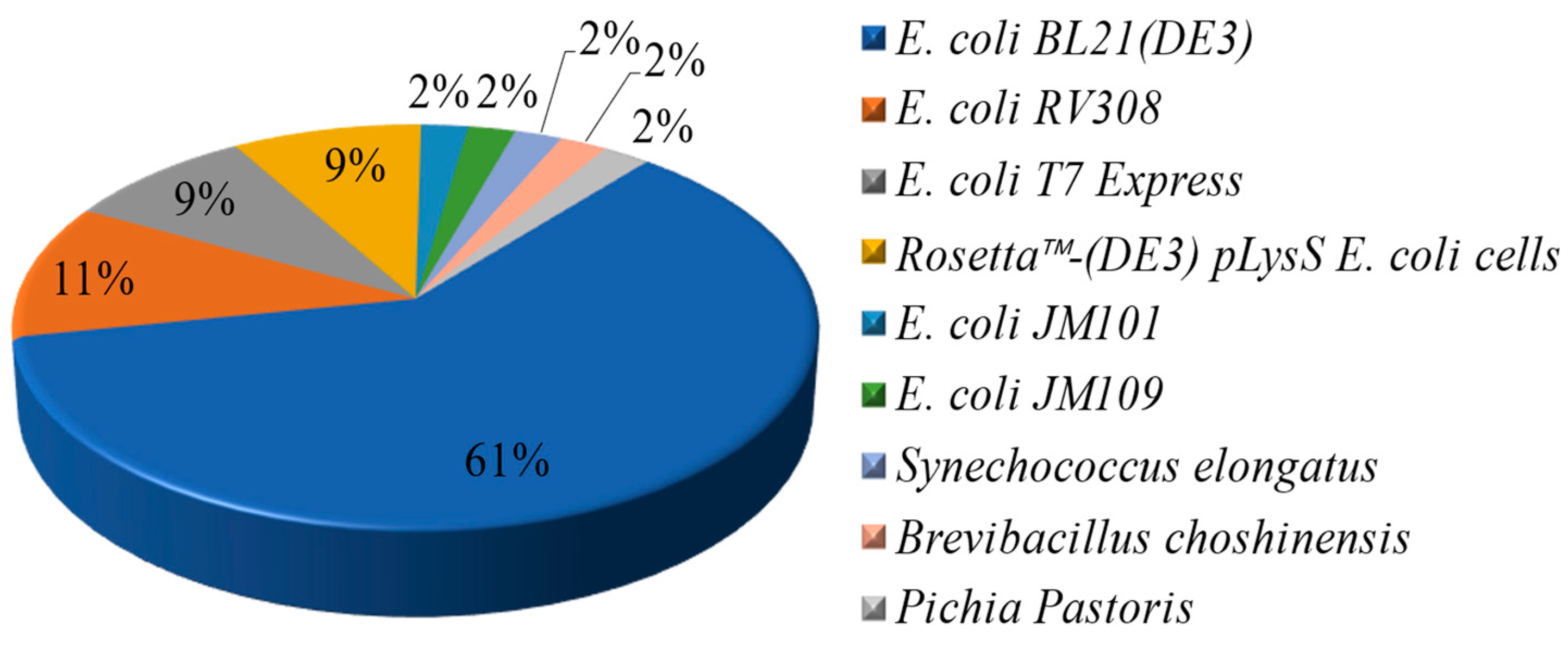
2.1. Expression of LPMOs in Escherichia coli (E. coli)
2.2. Expression of LPMOs in Other Host Microorganisms
3. Gene Design and N-Terminal Processing of LPMOs
3.1. Selection of Signal Peptides for Secretion
3.1.1. Selection of Signal Peptides in E. coli
3.1.2. Native Signal Peptides of LPMOs
3.1.3. Other Host-Specific Signal Peptides
3.2. Assistance of Chaperone Molecules and Selection of Protein Tags
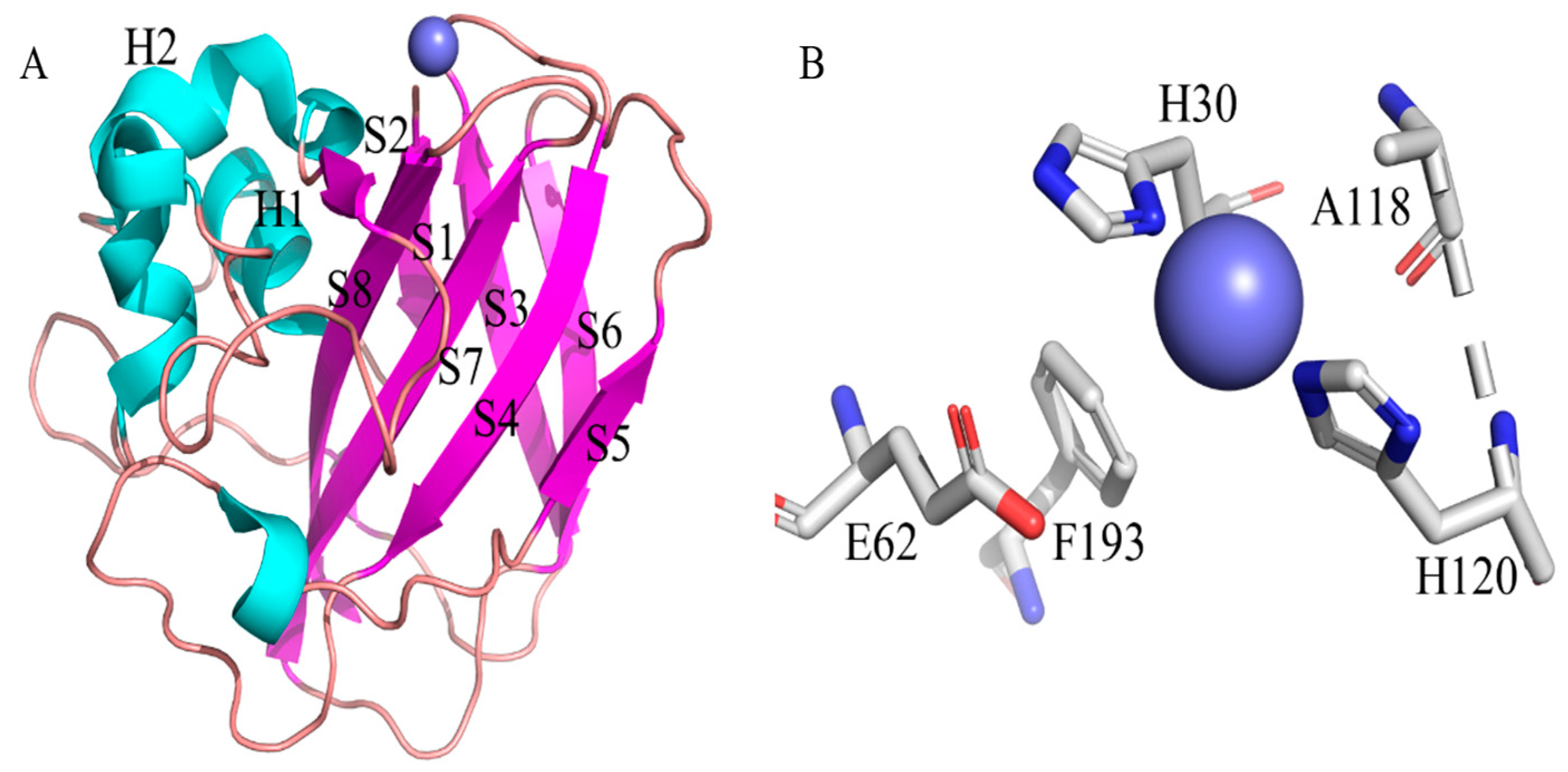
4. The Influence of Copper Ions on the Stability of LPMOs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Courtade, G.; Forsberg, Z.; Heggset, E.B.; Eijsink, V.G.H.; Aachmann, F.L. The carbohydrate-binding module and linker of a modular lytic polysaccharide monooxygenase promote localized cellulose oxidation. J. Biol. Chem. 2018, 293, 13006–13015. [Google Scholar] [CrossRef] [PubMed]
- Karkehabadi, S.; Hansson, H.; Kim, S.; Piens, K.; Mitchinson, C.; Sandgren, M. The First Structure of a Glycoside Hydrolase Family 61 Member, Cel61B from Hypocrea jecorina, at 1.6 angstrom Resolution. J. Mol. Biol. 2008, 383, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Vaaje-Kolstad, G.; Westereng, B.; Horn, S.J.; Liu, Z.L.; Zhai, H.; Sorlie, M.; Eijsink, V.G.H. An Oxidative Enzyme Boosting the Enzymatic Conversion of Recalcitrant Polysaccharides. Science 2010, 330, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, Z.; Vaaje-Kolstad, G.; Westereng, B.; Bunaes, A.C.; Stenstrom, Y.; MacKenzie, A.; Sorlie, M.; Horn, S.J.; Eijsink, V.G.H. Cleavage of cellulose by a CBM33 protein. Protein Sci. 2011, 20, 1479–1483. [Google Scholar] [CrossRef]
- Pierce, B.C.; Agger, J.W.; Zhang, Z.; Wichmann, J.; Meyer, A.S. A comparative study on the activity of fungal lytic polysaccharide monooxygenases for the depolymerization of cellulose in soybean spent flakes. Carbohydr. Res. 2017, 449, 85–94. [Google Scholar] [CrossRef]
- Levasseur, A.; Drula, E.; Lombard, V.; Coutinho, P.M.; Henrissat, B. Expansion of the enzymatic repertoire of the CAZy database to integrate auxiliary redox enzymes. Biotechnol. Biofuels 2013, 6, 41. [Google Scholar] [CrossRef]
- Gaber, Y.; Rashad, B.; Hussein, R.; Abdelgawad, M.; Ali, N.S.; Dishisha, T.; Varnai, A. Heterologous expression of lytic polysaccharide monooxygenases (LPMOs). Biotechnol. Adv. 2020, 43, 107583. [Google Scholar] [CrossRef]
- Correa, T.L.R.; Junior, A.T.; Wolf, L.D.; Buckeridge, M.S.; Dos Santos, L.V.; Murakami, M.T. An actinobacteria lytic polysaccharide monooxygenase acts on both cellulose and xylan to boost biomass saccharification. Biotechnol. Biofuels 2019, 12, 117. [Google Scholar] [CrossRef]
- Limsakul, P.; Phitsuwan, P.; Waeonukul, R.; Pason, P.; Tachaapaikoon, C.; Poomputsa, K.; Kosugi, A.; Sakka, M.; Sakka, K.; Ratanakhanokchai, K. A novel AA10 from Paenibacillus curdlanolyticus and its synergistic action on crystalline and complex polysaccharides. Appl. Microbiol. Biotechnol. 2020, 104, 7533–7550. [Google Scholar] [CrossRef]
- Li, J.; Solhi, L.; Goddard-Borger, E.D.; Mathieu, Y.; Wakarchuk, W.W.; Withers, S.G.; Brumer, H. Four cellulose-active lytic polysaccharide monooxygenases from Cellulomonas species. Biotechnol. Biofuels 2021, 14, 29. [Google Scholar] [CrossRef]
- Guo, X.; An, Y.; Lu, F.; Liu, F. Optimization of synergistic degradation of steam exploded corn straw by lytic polysaccharide monooxygenase R17L and cellulase. Ind. Crops Prod. 2022, 182, 114924. [Google Scholar] [CrossRef]
- Li, F.; Liu, Y.; Liu, Y.; Li, Y.; Yu, H. Heterologous expression and characterization of a novel lytic polysaccharide monooxygenase from Natrialbaceae archaeon and its application for chitin biodegradation. Bioresour. Technol. 2022, 354, 127174. [Google Scholar] [CrossRef]
- Chaplin, A.K.; Wilson, M.T.; Hough, M.A.; Svistunenko, D.A.; Hemsworth, G.R.; Walton, P.H.; Vijgenboom, E.; Worrall, J.A.R. Heterogeneity in the Histidine-brace Copper Coordination Sphere in Auxiliary Activity Family 10 (AA10) Lytic Polysaccharide Monooxygenases. J. Biol. Chem. 2016, 291, 12838–12850. [Google Scholar] [CrossRef]
- Forsberg, Z.; Bissaro, B.; Gullesen, J.; Dalhus, B.; Vaaje-Kolstad, G.; Eijsink, V.G.H. Structural determinants of bacterial lytic polysaccharide monooxygenase functionality. J. Biol. Chem. 2018, 293, 1397–1412. [Google Scholar] [CrossRef]
- Frommhagen, M.; Westphal, A.H.; van Berkel, W.J.H.; Kabel, M.A. Distinct Substrate Specificities and Electron-Donating Systems of Fungal Lytic Polysaccharide Monooxygenases. Front. Microbiol. 2018, 9, 1080. [Google Scholar] [CrossRef]
- Courtade, G.; Aachmann, F.L. Chitin-Active Lytic Polysaccharide Monooxygenases. Adv. Exp. Med. Biol. 2019, 1142, 115–129. [Google Scholar] [CrossRef]
- Sato, K.; Chiba, D.; Yoshida, S.; Takahashi, M.; Totani, K.; Shida, Y.; Ogasawara, W.; Nakagawa, Y.S. Functional analysis of a novel lytic polysaccharide monooxygenase from Streptomyces griseus on cellulose and chitin. Int. J. Biol. Macromol. 2020, 164, 2085–2091. [Google Scholar] [CrossRef]
- Gregory, R.C.; Hemsworth, G.R.; Turkenburg, J.P.; Hart, S.J.; Walton, P.H.; Davies, G.J. Activity, stability and 3-D structure of the Cu(ii) form of a chitin-active lytic polysaccharide monooxygenase from Bacillus amyloliquefaciens. Dalton Trans. 2016, 45, 16904–16912. [Google Scholar] [CrossRef]
- Courtade, G.; Le, S.B.; Saetrom, G.I.; Brautaset, T.; Aachmann, F.L. A novel expression system for lytic polysaccharide monooxygenases. Carbohydr. Res. 2017, 448, 212–219. [Google Scholar] [CrossRef]
- Courtade, G.; Balzer, S.; Forsberg, Z.; Vaaje-Kolstad, G.; Eijsink, V.G.; Aachmann, F.L. (1)H, (13)C, (15)N resonance assignment of the chitin-active lytic polysaccharide monooxygenase BlLPMO10A from Bacillus licheniformis. Biomol. NMR Assign. 2015, 9, 207–210. [Google Scholar] [CrossRef]
- Manjeet, K.; Purushotham, P.; Neeraja, C.; Podile, A.R. Bacterial chitin binding proteins show differential substrate binding and synergy with chitinases. Microbiol. Res. 2013, 168, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Manjeet, K.; Madhuprakash, J.; Mormann, M.; Moerschbacher, B.M.; Podile, A.R. A carbohydrate binding module-5 is essential for oxidative cleavage of chitin by a multi-modular lytic polysaccharide monooxygenase from Bacillus thuringiensis serovar kurstaki. Int. J. Biol. Macromol. 2019, 127, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, Y.; Cao, H.; Mou, G.; Yin, H. Expression and characterization of a lytic polysaccharide monooxygenase from Bacillus thuringiensis. Int. J. Biol. Macromol. 2015, 79, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, Z.; Nelson, C.E.; Dalhus, B.; Mekasha, S.; Loose, J.S.; Crouch, L.I.; Rohr, A.K.; Gardner, J.G.; Eijsink, V.G.; Vaaje-Kolstad, G. Structural and Functional Analysis of a Lytic Polysaccharide Monooxygenase Important for Efficient Utilization of Chitin in Cellvibrio japonicus. J. Biol. Chem. 2016, 291, 7300–7312. [Google Scholar] [CrossRef]
- Vaaje-Kolstad, G.; Bohle, L.A.; Gaseidnes, S.; Dalhus, B.; Bjoras, M.; Mathiesen, G.; Eijsink, V.G.H. Characterization of the Chitinolytic Machinery of Enterococcus faecalis V583 and High-Resolution Structure of Its Oxidative CBM33 Enzyme. J. Mol. Biol. 2012, 416, 239–254. [Google Scholar] [CrossRef]
- Mekasha, S.; Forsberg, Z.; Dalhus, B.; Bacik, J.P.; Choudhary, S.; Schmidt-Dannert, C.; Vaaje-Kolstad, G.; Eijsink, V.G. Structural and functional characterization of a small chitin-active lytic polysaccharide monooxygenase domain of a multi-modular chitinase from Jonesia denitrificans. FEBS Lett. 2016, 590, 34–42. [Google Scholar] [CrossRef]
- Christensen, I.A.; Eijsink, V.G.H.; Aachmann, F.L.; Courtade, G. (1)H, (13)C, (15)N resonance assignment of the apo form of the small, chitin-active lytic polysaccharide monooxygenase JdLPMO10A from Jonesia denitrificans. Biomol. NMR Assign. 2021, 15, 79–84. [Google Scholar] [CrossRef]
- Hamre, A.G.; Eide, K.B.; Wold, H.H.; Sorlie, M. Activation of enzymatic chitin degradation by a lytic polysaccharide monooxygenase. Carbohydr. Res. 2015, 407, 166–169. [Google Scholar] [CrossRef]
- Valenzuela, S.V.; Ferreres, G.; Margalef, G.; Pastor, F.I.J. Fast purification method of functional LPMOs from Streptomyces ambofaciens by affinity adsorption. Carbohydr. Res. 2017, 448, 205–211. [Google Scholar] [CrossRef]
- Forsberg, Z.; Mackenzie, A.K.; Sorlie, M.; Rohr, A.K.; Helland, R.; Arvai, A.S.; Vaaje-Kolstad, G.; Eijsink, V.G. Structural and functional characterization of a conserved pair of bacterial cellulose-oxidizing lytic polysaccharide monooxygenases. Proc. Natl. Acad. Sci. USA 2014, 111, 8446–8451. [Google Scholar] [CrossRef]
- Tanghe, M.; Danneels, B.; Last, M.; Beerens, K.; Stals, I.; Desmet, T. Disulfide bridges as essential elements for the thermostability of lytic polysaccharide monooxygenase LPMO10C from Streptomyces coelicolor. Protein Eng. Des. Sel. 2017, 30, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Courtade, G.; Forsberg, Z.; Vaaje-Kolstad, G.; Eijsink, V.G.H.; Aachmann, F.L. Chemical shift assignments for the apo-form of the catalytic domain, the linker region, and the carbohydrate-binding domain of the cellulose-active lytic polysaccharide monooxygenase ScLPMO10C. Biomol. NMR Assign. 2017, 11, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Kruer-Zerhusen, N.; Alahuhta, M.; Lunin, V.V.; Himmel, M.E.; Bomble, Y.J.; Wilson, D.B. Structure of a Thermobifida fusca lytic polysaccharide monooxygenase and mutagenesis of key residues. Biotechnol. Biofuels 2017, 10, 243. [Google Scholar] [CrossRef]
- Wong, E.; Vaaje-Kolstad, G.; Ghosh, A.; Hurtado-Guerrero, R.; Konarev, P.V.; Ibrahim, A.F.M.; Svergun, D.I.; Eijsink, V.G.H.; Chatterjee, N.S.; van Aalten, D.M.F. The Vibrio cholerae Colonization Factor GbpA Possesses a Modular Structure that Governs Binding to Different Host Surfaces. PLoS Pathog. 2012, 8, e1002373. [Google Scholar] [CrossRef] [PubMed]
- Paspaliari, D.K.; Loose, J.S.; Larsen, M.H.; Vaaje-Kolstad, G. Listeria monocytogenes has a functional chitinolytic system and an active lytic polysaccharide monooxygenase. FEBS J. 2015, 282, 921–936. [Google Scholar] [CrossRef]
- Mutahir, Z.; Mekasha, S.; Loose, J.S.M.; Abbas, F.; Vaaje-Kolstad, G.; Eijsink, V.G.H.; Forsberg, Z. Characterization and synergistic action of a tetra-modular lytic polysaccharide monooxygenase from Bacillus cereus. FEBS Lett. 2018, 592, 2562–2571. [Google Scholar] [CrossRef]
- Stepnov, A.A.; Forsberg, Z.; Sorlie, M.; Nguyen, G.S.; Wentzel, A.; Rohr, A.K.; Eijsink, V.G.H. Unraveling the roles of the reductant and free copper ions in LPMO kinetics. Biotechnol. Biofuels 2021, 14, 28. [Google Scholar] [CrossRef]
- Zhang, A.; Mo, X.; Zhou, N.; Wang, Y.; Wei, G.; Hao, Z.; Chen, K. Identification of Chitinolytic Enzymes in Chitinolyticbacter meiyuanensis and Mechanism of Efficiently Hydrolyzing Chitin to N-Acetyl Glucosamine. Front. Microbiol. 2020, 11, 572053. [Google Scholar] [CrossRef]
- Wang, D.; Li, J.; Wong, A.C.Y.; Aachmann, F.L.; Hsieh, Y.S.Y. A colorimetric assay to rapidly determine the activities of lytic polysaccharide monooxygenases. Biotechnol. Biofuels 2018, 11, 215. [Google Scholar] [CrossRef]
- Sunna, A.; Gibbs, M.D.; Chin, C.W.J.; Nelson, P.J.; Bergquist, P.L. A gene encoding a novel multidomain beta-1,4-mannanase from Caldibacillus cellulovorans and action of the recombinant enzyme on kraft pulp. Appl. Environ. Microbiol. 2000, 66, 664–670. [Google Scholar] [CrossRef]
- Mitsuhashi, W.; Saito, H.; Sato, M. Complete nucleotide sequence of the fusolin gene of an entomopoxvirus in the cupreous chafer, Anomala cuprea Hope (Coleoptera: Scarabaeidae). Insect Biochem. Mol. Biol. 1997, 27, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Ghatge, S.S.; Telke, A.A.; Waghmode, T.R.; Lee, Y.; Lee, K.W.; Oh, D.B.; Shin, H.D.; Kim, S.W. Multifunctional cellulolytic auxiliary activity protein HcAA10-2 from Hahella chejuensis enhances enzymatic hydrolysis of crystalline cellulose. Appl. Microbiol. Biotechnol. 2015, 99, 3041–3055. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.C.; Madupu, R.; Durkin, A.S.; Ekborg, N.A.; Pedamallu, C.S.; Hostetler, J.B.; Radune, D.; Toms, B.S.; Henrissat, B.; Coutinho, P.M.; et al. The Complete Genome of Teredinibacter turnerae T7901: An Intracellular Endosymbiont of Marine Wood-Boring Bivalves (Shipworms). PLoS ONE 2009, 4, e6085. [Google Scholar] [CrossRef]
- Fowler, C.A.; Sabbadin, F.; Ciano, L.; Hemsworth, G.R.; Elias, L.; Bruce, N.; McQueen-Mason, S.; Davies, G.J.; Walton, P.H. Discovery, activity and characterisation of an AA10 lytic polysaccharide oxygenase from the shipworm symbiont Teredinibacter turnerae. Biotechnol. Biofuels 2019, 12, 232. [Google Scholar] [CrossRef]
- Rodrigues, K.B.; Macedo, J.K.A.; Teixeira, T.; Barros, J.S.; Araujo, A.C.B.; Santos, F.P.; Quirino, B.F.; Brasil, B.; Salum, T.F.C.; Abdelnur, P.V.; et al. Recombinant expression of Thermobifida fusca E7 LPMO in Pichia pastoris and Escherichia coli and their functional characterization. Carbohydr. Res. 2017, 448, 175–181. [Google Scholar] [CrossRef]
- Russo, D.A.; Zedler, J.A.Z.; Wittmann, D.N.; Mollers, B.; Singh, R.K.; Batth, T.S.; van Oort, B.; Olsen, J.V.; Bjerrum, M.J.; Jensen, P.E. Expression and secretion of a lytic polysaccharide monooxygenase by a fast-growing cyanobacterium. Biotechnol. Biofuels 2019, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.J.; Yoon, S.H.; Kim, Y.W. Overproduction and characterization of a lytic polysaccharide monooxygenase in Bacillus subtilis using an assay based on ascorbate consumption. Enzym. Microb. Technol. 2016, 93–94, 150–156. [Google Scholar] [CrossRef]
- Nakagawa, Y.S.; Kudo, M.; Loose, J.S.; Ishikawa, T.; Totani, K.; Eijsink, V.G.; Vaaje-Kolstad, G. A small lytic polysaccharide monooxygenase from Streptomyces griseus targeting alpha- and beta-chitin. FEBS J. 2015, 282, 1065–1079. [Google Scholar] [CrossRef]
- Yadav, S.K.; Archana; Singh, R.; Singh, P.K.; Vasudev, P.G. Insecticidal fern protein Tma12 is possibly a lytic polysaccharide monooxygenase. Planta 2019, 249, 1987–1996. [Google Scholar] [CrossRef]
- Vaaje-Kolstad, G.; Houston, D.R.; Riemen, A.H.K.; Eijsink, V.G.H.; van Aalten, D.M.F. Crystal structure and binding properties of the Serratia marcescens chitin-binding protein CBP21. J. Biol. Chem. 2005, 280, 11313–11319. [Google Scholar] [CrossRef]
- Han, R.; Li, J.; Shin, H.-d.; Chen, R.R.; Du, G.; Liu, L.; Chen, J. Recent advances in discovery, heterologous expression, and molecular engineering of cyclodextrin glycosyltransferase for versatile applications. Biotechnol. Adv. 2014, 32, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Mekasha, S.; Tuveng, T.R.; Askarian, F.; Choudhary, S.; Schmidt-Dannert, C.; Niebisch, A.; Modregger, J.; Vaaje-Kolstad, G.; Eijsink, V.G.H. A trimodular bacterial enzyme combining hydrolytic activity with oxidative glycosidic bond cleavage efficiently degrades chitin. J. Biol. Chem. 2020, 295, 9134–9146. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, J.; Liu, X.; Pan, X.; Hou, J.; Ran, C.; Zhou, Z. Improving extracellular production of Serratia marcescens lytic polysaccharide monooxygenase CBP21 and Aeromonas veronii B565 chitinase Chi92 in Escherichia coli and their synergism. Amb. Express 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Haacke, A.; Fendrich, G.; Ramage, P.; Geiser, M. Chaperone over-expression in Escherichia coli: Apparent increased yields of soluble recombinant protein kinases are due mainly to soluble aggregates. Protein Expr. Purif. 2009, 64, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Watly, J.; Simonoysky, E.; Wieczorek, R.; Barbosa, N.; Miller, Y.; Kozlowski, H. Insight into the Coordination and the Binding Sites of Cu2+ by the Histidyl-6-Tag using Experimental and Computational Tools. Inorg. Chem. 2014, 53, 6675–6683. [Google Scholar] [CrossRef]
- Eijsink, V.G.H.; Petrovic, D.; Forsberg, Z.; Mekasha, S.; Rohr, A.K.; Varnai, A.; Bissaro, B.; Vaaje-Kolstad, G. On the functional characterization of lytic polysaccharide monooxygenases (LPMOs). Biotechnol. Biofuels 2019, 12, 58. [Google Scholar] [CrossRef]
- Ayala, J.C.; Pimienta, E.; Rodriguez, C.; Anne, J.; Vallin, C.; Milanes, M.T.; King-Batsios, E.; Huygen, K.; Van Mellaert, L. Use of Strep-tag II for rapid detection and purification of Mycobacterium tuberculosis recombinant antigens secreted by Streptomyces lividans. J. Microbiol. Methods 2013, 94, 192–198. [Google Scholar] [CrossRef]
- Sabbadin, F.; Hemsworth, G.R.; Ciano, L.; Henrissat, B.; Dupree, P.; Tryfona, T.; Marques, R.D.S.; Sweeney, S.T.; Besser, K.; Elias, L.; et al. An ancient family of lytic polysaccharide monooxygenases with roles in arthropod development and biomass digestion. Nat. Commun. 2018, 9, 756. [Google Scholar] [CrossRef]
- Frandsen, K.E.H.; Poulsen, J.C.N.; Tovborg, M.; Johansen, K.S.; Lo Leggio, L. Learning from oligosaccharide soaks of crystals of an AA13 lytic polysaccharide monooxygenase: Crystal packing, ligand binding and active-site disorder. Acta Crystallogr. Sect. D-Struct. Biol. 2017, 73, 64–76. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genebank No. | Resource Microorganisms | Expressing Host | Expressing Vector | Ref. |
|---|---|---|---|---|
| CBI42985.1 | Bacillus amyloliquefaciens DSM 7 | E. coli BL21(DE3) | pET SUMO | [18] |
| ATI74801.1 | Bacillus licheniformis ATCC 9789 | E. coli BL21(DE3) E. coli RV308 | pJB | [19,20] |
| AAU39477.1 | Bacillus licheniformis DSM 13 (ATCC 14580) | E. coli BL21(DE3) | pET-22b; pET-28a | [21] |
| AJP62637.1 | Bacillus thuringiensis serovar kurstaki ACCC 10066 | E. coli BL21(DE3) | pET-22b | [22,23] |
| ACE83992.1 | Cellvibrio japonicus Ueda107 | E. coli BL21(DE3); E. coli T7 Express; E. coli RV308 | pRSET B; pJB | [19,24] |
| ACE84760.1 | Cellvibrio japonicus Ueda107 | E. coli BL21(DE3) | pET-22b | [24] |
| AAO80225.1 | Enterococcus faecalis V583 | E. coli BL21(DE3) | pRSET B | [25] |
| ACV09037.1 | Jonesia denitrificans DSM 20603 | E. coli T7 Express; E. coli RV308 | pRSET; pUCBB-eGFP | [19,26,27] |
| - | Jonesia denitrificans | Escherichia coli | - | [26] |
| AAU88202.1 | Serratia marcescens BJL200 | E. coli T7 Express; E. coli RV308 | pUC57; pJB | [14,19,28] |
| CAJ89556.1 | Streptomyces ambofaciens ATCC 23877 | E. coli BL21(DE3) | pET-11a | [29] |
| CAJ90160.1 | pET-11a | [29] | ||
| CAB61160.1 | Streptomyces coelicolor A3(2) | E. coli BL21(DE3) | pRSET B | [30] |
| CAB61600.1 | Streptomyces coelicolor A3(2) | E. coli BL21(DE3); E. coli T7 Express; E. coli RV308; E. coli XL10 gold | pETite N-His SUMO T7; pCXP34; pRSET B; pJB; pWW0 | [1,19,30,31,32] |
| EOY47895.1 | Streptomyces lividans 1326 | E. coli BL21(DE3) | pET-26b | [13] |
| AAZ55700.1 | Thermobifida fusca YX | E. coli BL21(DE3) | pET-26b; pET-27b; pRSET B | [30,33] |
| AAF96709.1 | Vibrio cholerae O1 biovar El Tor str. N16961 | E. coli BL21(DE3) | pET-22b | [34] |
| ABA49030.1 | Burkholderia pseudomallei 1710b | E. coli BL21(DE3) | - | - |
| AEO07443.1 | Listeria monocytogenes 10403S | E. coli BL21(DE3) | pET46 Ek⁄LIC | [35] |
| ADL45185.1 | Micromonospora aurantiaca ATCC 27029 | E. coli BL21(DE3); E. coli T7 Express; E. coli RV308 | pRSET B; pUC57; pJB | [14,19] |
| - | Micromonospora aurantiaca ATCC 27029 | E. coli BL21(DE3) | pRSET B | [14] |
| D9T1F0 | E. coli BL21(DE3) | pRSET B | [14] | |
| - | E. coli BL21(DE3) | pRSET B | [14] | |
| BAG17028.1 | Streptomyces griseus subsp. griseus NBRC 13350 | E. coli BL21(DE3) | pNCMO2 | [17] |
| AAP09751.1 | Bacillus cereus ATCC 14579 | E. coli BL21(DE3) | pRSET B | [36] |
| MT882343 | Streptomyces griseolus | E. coli BL21(DE3) | pET-26b | [37] |
| AIY28331.1. | Paenibacillus curdlanolyticus B-6 | E. coli BL21(DE3) | pET-28a | [9] |
| AAO80225.1 | Enterococcus faecalis V583 | E. coli BL21(DE3) | pRSETB | [25] |
| WP_099398589.1 | Chitinolyticbacter meiyuanensis SYBC-H1 | E. coli BL21(DE3) | pET-28a | [38] |
| WP_039915213.1 | Cellvibrio mixtus | E. coli BL21(DE3) | pET-26b | [39] |
| AEE44415.1 | Cellulomonas fimi strain ATCC 484 | -Rosetta™-(DE3) pLysS E. coli cells | pET-29b | [10] |
| ADG73094.1 | Cellulomonas flavigena DSM 20109 | -Rosetta™-(DE3) pLysS E. coli cells | pET-29b | [10] |
| ADG73091.1 | Cellulomonas flavigena DSM 20109 | -Rosetta™-(DE3) pLysS E. coli cells | pET-29b | [10] |
| ADG73234.1 | Cellulomonas flavigena DSM 20109 | -Rosetta™-(DE3) pLysS E. coli cells | pET-29b | [10] |
| AAF22274.1 | Caldibacillu cellulovorans | E. coli JM101 | pJLA602 | [40] |
| BAA25629.1 | Anomala cuprea entomopoxvirus | E. coli JM109 | pBluescript II KS | [41] |
| ABC27701.1 | Hahella chejuensis KCTC 2396 | Escherichia coli | pET-28a | [42] |
| CAE14645.1 | Photorhabdus laumondii subsp. laumondii TTO1 | Escherichia coli | pBluescript SK | [43] |
| ACR14100.1 | Teredinibacter turnerae T7901 | Escherichia coli | pET-22b | [43,44] |
| ADW01716.1 | Streptomyces pratensis ATCC 33331 | Escherichia coli | - | - |
| AAZ55306.1 | Thermobifida fusca YX | E. coli BL21(DE3); Pichia Pastoris; Synechococcus elongatus UTEX 2973 | pET-21a; pPICZαA ;pET-26b; pRSET B; pDF-trc | [30,33,45,46] |
| ADP32663 | Bacillus atrophaeus 1942 | Bacillus subtilis LKS87 | pUBRTA | [47] |
| BAG23684.1 | Streptomyces griseus subsp. griseus NBRC 13350 | Brevibacillus choshinensis | pNCMO2 | [48] |
| - | Kitasatospora papulosa (DSM41643) | Escherichia coli | pET-22b | [8] |
| AFR32946.1 | Tectaria macrodonta | - | - | [49] |
| Expressing Host | Signal Peptide | Ref. |
|---|---|---|
| E. coli | PelB | [8,33,37,44] |
| OmpA | [52] | |
| SmAA10A | [14] | |
| Native | [14,27,29,30] | |
| SacB, pelB, TorA, WompA, OmpASIL2, LM-SEA, LSEA-mut, Exyl, gIII, STII, XCs, and CBHI | [53] | |
| Synechococcus elongatus | Native, TorA | [46] |
| Pichia pastoris | α-mating factor | [45] |
| Brevibacillus choshinensis | Native | [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zhou, Z.; Lou, T.; Xiang, R.; Zhang, D.; Wang, D.; Wang, S. Advance in Heterologous Expression of Biomass-Degrading Auxiliary Activity 10 Family of Lytic Polysaccharide Monooxygenases. Fermentation 2023, 9, 795. https://doi.org/10.3390/fermentation9090795
Zhang H, Zhou Z, Lou T, Xiang R, Zhang D, Wang D, Wang S. Advance in Heterologous Expression of Biomass-Degrading Auxiliary Activity 10 Family of Lytic Polysaccharide Monooxygenases. Fermentation. 2023; 9(9):795. https://doi.org/10.3390/fermentation9090795
Chicago/Turabian StyleZhang, Hongyu, Zixuan Zhou, Tingting Lou, Rong Xiang, Deguang Zhang, Danyun Wang, and Suying Wang. 2023. "Advance in Heterologous Expression of Biomass-Degrading Auxiliary Activity 10 Family of Lytic Polysaccharide Monooxygenases" Fermentation 9, no. 9: 795. https://doi.org/10.3390/fermentation9090795