
Effect of Inoculation with Lacticaseibacillus casei and Staphylococcus carnosus on the Quality of Squid (Dosidicus gigas) Surimi Sausage
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Fermented Squid Surimi Sausages
2.3. Determination of pH
2.4. Microbiological Analysis
2.5. DNA Extraction and Sequencing
2.5.1. DNA Extraction, PCR, and Libraries Construction
2.5.2. Sequencing Data Processing and Analysis
2.6. Determination of Biogenic Amines
2.7. Texture Profile Analyses, and Rupture and Color Tests
2.8. Appearance Photograph
2.9. Statistical Analysis
3. Results and Discussion
3.1. Effect of Inoculation on pH of the Squid Surimi Sausages
3.2. LAB and Staphylococcus Counts during Fermentation
3.3. Effect of Inoculation on the Bacterial Community of the Squid Surimi Sausages
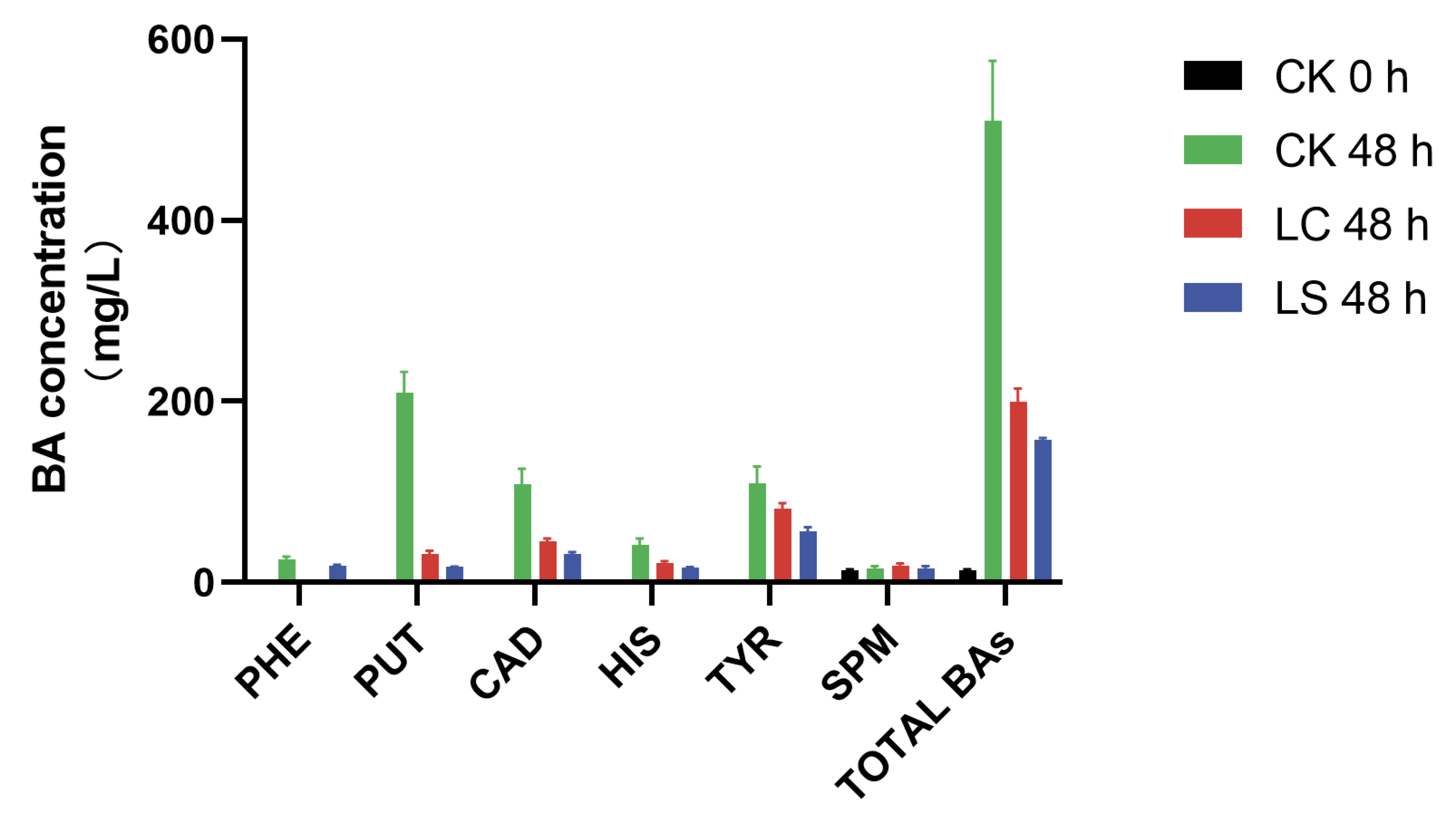
3.4. Effect of Inoculation on the BAs Accumulation of Squid Surimi Sausages
3.5. Effect of Inoculation on the Texture, Rupture Strength, and Color of Squid Surimi Sausages
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munekata, P.E.S.; Pateiro, M.; Tomasevic, I.; Domínguez, R.; Barretto, A.C.d.S.; Santos, E.M.; Lorenzo, J.M. Functional fermented meat products with probiotics—A review. J. Appl. Microbiol. 2021, 133, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Baruah, R.; Ray, M.; Halami, P.M. Preventive and therapeutic aspects of fermented foods. J. Appl. Microbiol. 2022, 132, 3476–3489. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ji, L.; Zhang, J.; Zhao, Z.; Zhang, R.; Bai, T.; Hou, B.; Zhang, Y.; Liu, D.; Wang, W.; et al. A Review: Microbial Diversity and Function of Fermented Meat Products in China. Front. Microbiol. 2021, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Potes, M.E.; Elias, M. Role of Starter Cultures on the Safety of Fermented Meat Products. Front. Microbiol. 2019, 10, 853. [Google Scholar] [CrossRef]
- Dos Santos Cruxen, C.E.; Funck, G.D.; Haubert, L.; da Silva Dannenberg, G.; de Lima Marques, J.; Chaves, F.C.; da Silva, W.P.; Fiorentini, Â.M. Selection of native bacterial starter culture in the production of fermented meat sausages: Application potential, safety aspects, and emerging technologies. Food Res. Int. 2019, 122, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Lu, H. Study on Myofibrils and Myofibrils Gel Properties of Jumbo squid (Dosidicus gigas). Ph.D. Thesis, Zhejiang Gongshang University, Hangzhou, China, 2009. (In Chinese). [Google Scholar]
- Gómez-Guillén, M.C.; Montero, P.; Solas, M.T.; Borderías, A.J. Thermally Induced Aggregation of Giant Squid (Dosidicus gigas) Mantle Proteins. Physicochemical Contribution of Added Ingredients. J. Agric. Food Chem. 1998, 46, 3440–3446. [Google Scholar] [CrossRef]
- Yang, R.; Xu, A.; Chen, Y.; Sun, N.; Zhang, J.; Jia, R.; Huang, T.; Yang, W. Effect of laver powder on textual, rheological properties and water distribution of squid (Dosidicus gigas) surimi gel. J. Texture Stud. 2020, 51, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Deng, S.; Lv, G.; Li, M.; Bao, H.; Gao, Y.; Jia, R. Improvement of Gel Quality of Squid (Dosidicus gigas) Meat by Using Sodium Gluconate, Sodium Citrate, and Sodium Tartrate. Foods 2022, 11, 173. [Google Scholar] [CrossRef]
- Li, D.-Y.; Tan, Z.-F.; Liu, Z.-Q.; Wu, C.; Liu, H.-L.; Guo, C.; Zhou, D.-Y. Effect of hydroxyl radical induced oxidation on the physicochemical and gelling properties of shrimp myofibrillar protein and its mechanism. Food Chem. 2021, 351, 129344. [Google Scholar] [CrossRef]
- Hu, Y.; Xia, W.; Ge, C. Effect of mixed starter cultures fermentation on the characteristics of silver carp sausages. World J. Microbiol. Biotechnol. 2007, 23, 1021–1031. [Google Scholar] [CrossRef]
- Sidira, M.; Kandylis, P.; Kanellaki, M.; Kourkoutas, Y. Effect of immobilized Lactobacillus casei on the evolution of flavor compounds in probiotic dry-fermented sausages during ripening. Meat Sci. 2015, 100, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Szymański, P.; Łaszkiewicz, B.; Siekierko, U.; Kołożyn-Krajewska, D. Effects of the Use of Staphylococcus carnosus in the Curing Process of Meat with a Reduced Amount of Sodium Nitrite on Colour, Residue Nitrite and Nitrate, Content of Nitrosyl Pigments, and Microbiological and the Sensory Quality of Cooked Meat Product. J. Food Qual. 2020, 8, 2020. [Google Scholar] [CrossRef]
- Yang, W.; Chen, S.; Yu, S. Effects of Different Soaking Processes and Additives on the Acid Removal Efficiency of Squid (Dosidicus gigas). Jiangxi Fish. Sci. Technol. 2019, 5, 45–47. (In Chinese) [Google Scholar]
- Dongni, W. Study on Improvement Gel Properties of Squid (Illex argentinus) Surimi. Master’s Thesis, Dalian Polytechnic University, Dalian, China, 2016. (In Chinese). [Google Scholar]
- Ren, H.; Deng, Y.; Wang, X. Effect of a compound starter cultures inoculation on bacterial profile and biogenic amine accumulation in Chinese Sichuan sausages. Food Sci. Hum. Wellness 2022, 11, 341–348. [Google Scholar] [CrossRef]
- Hu, Y.; Xia, W.; Ge, C. Characterization of Fermented Silver Carp Sausages Inoculated with Mixed Starter Culture. LWT Food Sci. Technol. 2008, 41, 730–738. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Tasić, T.; Ikonić, P.; Mandić, A.; Jokanović, M.; Tomović, V.; Savatić, S.; Petrović, L. Biogenic amines content in traditional dry fermented sausage Petrovská klobása as possible indicator of good manufacturing practice. Food Control. 2012, 23, 107–112. [Google Scholar] [CrossRef]
- Ranran, L. Study on the Dynamic Changes and Control of Biogenic Amines in Low-Sodium Bacon. Master’s Thesis, Southwest University, Chongqing, China, 2020. (In Chinese). [Google Scholar]
- Wang, D.; Zhao, L.; Su, R.; Jin, Y. Effects of different starter culture combinations on microbial counts and physico-chemical properties in dry fermented mutton sausages. Food Sci. Nutr. 2019, 7, 1957–1968. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Regenstein, J.M.; Yu, D.; Yang, F.; Jiang, Q. Correlations between microbiota succession and flavor formation during fermentation of Chinese low-salt fermented common carp (Cyprinus carpio L.) inoculated with mixed starter cultures. Food Microbiol. 2020, 90, 103487. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Gómez, M.; Purriños, L.; Fonseca, S.; Rodriguez, J.M.L. Effect of commercial starter cultures on volatile compound profile and sensory characteristics of dry-cured foal sausage. J. Sci. Food Agric. 2016, 96, 1194–1201. [Google Scholar] [CrossRef]
- Wang, D.; Hu, G.; Wang, H.; Wang, L.; Zhang, Y.; Zou, Y.; Zhao, L.; Liu, F.; Jin, Y. Effect of Mixed Starters on Proteolysis and Formation of Biogenic Amines in Dry Fermented Mutton Sausages. Foods 2021, 10, 2939. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Y.; Sun, J.; Pan, P.; Liu, Y.; Tian, T. Effects of starter culture inoculation on microbial community diversity and food safety of Chinese Cantonese sausages by high-throughput sequencing. J. Food Sci. Technol. 2020, 58, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Dias, I.; Laranjo, M.; Potes, M.E.; Agulheiro-Santos, A.C.; Ricardo-Rodrigues, S.; Fraqueza, M.J.; Oliveira, M.; Elias, M. Staphylococcus spp. and Lactobacillus sakei Starters with High Level of Inoculation and an Extended Fermentation Step Improve Safety of Fermented Sausages. Fermentation 2022, 8, 49. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, X.; Ding, Y.; Ke, Z.; Zhou, X.; Zhang, J. Diversity and succession of the microbial community and its correlation with lipid oxidation in dry-cured black carp (Mylopharyngodon piceus) during storage. Food Microbiol. 2021, 98, 103686. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Lu, F.; Qiu, M.; Ding, Y.; Zhou, X. Dynamics and Diversity of Microbial Community Succession of Surimi During Fermentation with Next-Generation Sequencing. J. Food Saf. 2015, 36, 308–316. [Google Scholar] [CrossRef]
- Luo, M.; Chen, L.; Liu, H.; Jiang, J.; Lai, C.; Zhou, C.; Hong, P.; Gao, P. Analysis of changes in the microbial community structure and physicochemical properties during the fermentation of sand crab juice. FEMS Microbiol. Lett. 2021, 368, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhang, C.; Gu, Z.; Li, C.; Fang, Z.; Zeng, Z.; Zhang, Z.; Hu, B.; Chen, H.; Wu, W.; et al. Effect of microbial fermentation on the sensory characteristics and chemical compositions of Chinese sweet tea (Lithocarpus litseifolius (Hance) Chun. Food Biosci. 2022, 46, 101567. [Google Scholar] [CrossRef]
- Li, C.; Zhao, Y.; Wang, Y.; Li, L.; Yang, X.; Chen, S.; Zhao, Y.; Zhou, W. Microbial community changes induced by Pediococcus pentosaceus improve the physicochemical properties and safety in fermented tilapia sausage. Food Res. Int. 2021, 147, 110476. [Google Scholar] [CrossRef]
- Wang, J.; Hou, J.; Zhang, X.; Hu, J.; Yu, Z.; Zhu, Y. Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum. Foods 2022, 11, 736. [Google Scholar]
- Xiao, Y.; Liu, Y.; Chen, C.; Xie, T.; Li, P. Effect of Lactobacillus plantarum and Staphylococcus xylosus on flavour development and bacterial communities in Chinese dry fermented sausages. Food Res. Int. 2020, 135, 109247. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. A narrative review on biogenic amines in fermented fish and meat products. J. Food Sci. Technol. 2021, 58, 1623–1639. [Google Scholar] [CrossRef]
- Dias, I.; Laranjo, M.; Potes, M.E.; Agulheiro-Santos, A.C.; Ricardo-Rodrigues, S.; Fialho, A.R.; Véstia, J.; Fraqueza, M.J.; Oliveira, M.; Elias, M. Co-Inoculation with Staphylococcus equorum and Lactobacillus sakei Reduces Vasoactive Biogenic Amines in Traditional Dry-Cured Sausages. Int. J. Environ. Res. Public Health 2021, 18, 7100. [Google Scholar] [CrossRef] [PubMed]
- Dias, I.; Laranjo, M.; Potes, M.E.; Agulheiro-Santos, A.C.; Ricardo-Rodrigues, S.; Fialho, A.R.; Véstia, J.; Fraqueza, M.J.; Oliveira, M.; Elias, M. Autochthonous Starter Cultures Are Able to Reduce Biogenic Amines in a Traditional Portuguese Smoked Fermented Sausage. Microorganisms 2020, 8, 686. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xia, W.; Yang, F.; Kim, J.M.; Nie, X. Effect of fermentation temperature on the microbial and physicochemical properties of silver carp sausages inoculated with Pediococcus pentosaceus. Food Chem. 2010, 118, 512–518. [Google Scholar] [CrossRef]
- Hu, Y.; Xia, W.; Liu, X. Changes in biogenic amines in fermented silver carp sausages inoculated with mixed starter cultures. Food Chem. 2007, 104, 188–195. [Google Scholar] [CrossRef]
- Sun, Y.; Hua, Q.; Tian, X.; Xu, Y.; Gao, P.; Xia, W. Effect of starter cultures and spices on physicochemical properties and microbial communities of fermented fish (Suanyu) after fermentation and storage. Food Res. Int. 2022, 159, 111631. [Google Scholar] [CrossRef] [PubMed]
- Szczesniak, A.S. Texture is a sensory property. Food Qual. Prefer. 2002, 13, 215–225. [Google Scholar] [CrossRef]
- González-Fernández, C.; Santos, E.M.; Rovira, J.; Jaime, I. The effect of sugar concentration and starter culture on instrumental and sensory textural properties of chorizo-Spanish dry-cured sausage. Meat Sci. 2006, 74, 467–475. [Google Scholar] [CrossRef]
- Xu, J.; Chang, J.; Kong, B.; Xia, X.; Liu, Q. Effects of Water Addition on the Quality Characteristics of Starch-Meat Sausage. Sci. Technol. Food Ind. 2022, 43, 23–30. (In Chinese) [Google Scholar]
- Liu, Y.; Sun, Q.; Pan, Y.; Wei, S.; Xia, Q.; Liu, S.; Ji, H.; Deng, C.; Hao, J. Investigation on the correlation between changes in water and texture properties during the processing of surimi from golden pompano (Trachinotus ovatus). J. Food Sci. 2021, 86, 376–384. [Google Scholar] [CrossRef]
- Akan, S.; Özdestan-Ocak, Ö. Influence of grape seed extract on suppression of biogenic amine accumulation, chemical and color traits of wet tarhana during fermentation. Food Biosci. 2021, 42, 101065. [Google Scholar] [CrossRef]
- Wang, D.; Cheng, F.; Wang, Y.; Han, J.; Gao, F.; Tian, J.; Zhang, K.; Jin, Y. The Changes Occurring in Proteins during Processing and Storage of Fermented Meat Products and Their Regulation by Lactic Acid Bacteria. Foods 2022, 11, 2427. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Chao1 | Shannon Index | Simpson Index | Good’s Coverage |
|---|---|---|---|---|
| CK1 | 144 | 3.971 | 0.851 | 1 |
| CK2 | 160 | 3.458 | 0.76 | 1 |
| CK3 | 141 | 3.657 | 0.83 | 1 |
| CK4 | 161 | 4.045 | 0.869 | 1 |
| LC1 | 89 | 1.617 | 0.395 | 1 |
| LC2 | 76 | 1.522 | 0.383 | 1 |
| LC3 | 84 | 1.787 | 0.435 | 1 |
| LC4 | 97 | 1.811 | 0.439 | 1 |
| LS1 | 58 | 1.308 | 0.327 | 1 |
| LS2 | 80 | 1.421 | 0.343 | 1 |
| LS3 | 71 | 1.496 | 0.364 | 1 |
| LS4 | 62 | 1.523 | 0.371 | 1 |
| CK | LS | |
|---|---|---|
| TPA test | ||
| Hardness (g) | 1893.086 ± 194.019 a | 4219.882 ± 269.484 b |
| Springiness | 0.948 ± 0.039 a | 0.849 ± 0.019 b |
| Cohesivenes | 0.799 ± 0.007 a | 0.686 ± 0.040 b |
| Chewiness (g) | 1432.319 ± 139.941 a | 2457.859 ± 248.508 b |
| Rupture test | ||
| Rupture strength (g) | 178.071 ± 16.030 a | 355.209 ± 82.139 b |
| Rupture hardness (g) | 186.528 ± 17.234 a | 371.631 ± 92.166 b |
| Color test | ||
| L | 49.475 ± 0.289 a | 55.320 ± 1.726 b |
| a | 11.883 ± 0.493 a | 15.003 ± 0.408 b |
| b | 19.488 ± 0.330 a | 23.752 ± 0.618 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, H.; Weng, P.; Wu, Z. Effect of Inoculation with Lacticaseibacillus casei and Staphylococcus carnosus on the Quality of Squid (Dosidicus gigas) Surimi Sausage. Fermentation 2023, 9, 794. https://doi.org/10.3390/fermentation9090794
Mu H, Weng P, Wu Z. Effect of Inoculation with Lacticaseibacillus casei and Staphylococcus carnosus on the Quality of Squid (Dosidicus gigas) Surimi Sausage. Fermentation. 2023; 9(9):794. https://doi.org/10.3390/fermentation9090794
Chicago/Turabian StyleMu, Hongliang, Peifang Weng, and Zufang Wu. 2023. "Effect of Inoculation with Lacticaseibacillus casei and Staphylococcus carnosus on the Quality of Squid (Dosidicus gigas) Surimi Sausage" Fermentation 9, no. 9: 794. https://doi.org/10.3390/fermentation9090794