
Identification of a Strain Degrading Ammonia Nitrogen, Optimization of Ammonia Nitrogen Degradation Conditions, and Gene Expression of Key Degrading Enzyme Nitrite Reductase
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Source of the Wastewater Sample
2.2. The Main Culture Medium
2.3. Strains, Plasmids, Reagent, Medium, and Culture Condition
2.4. Identification of Strain
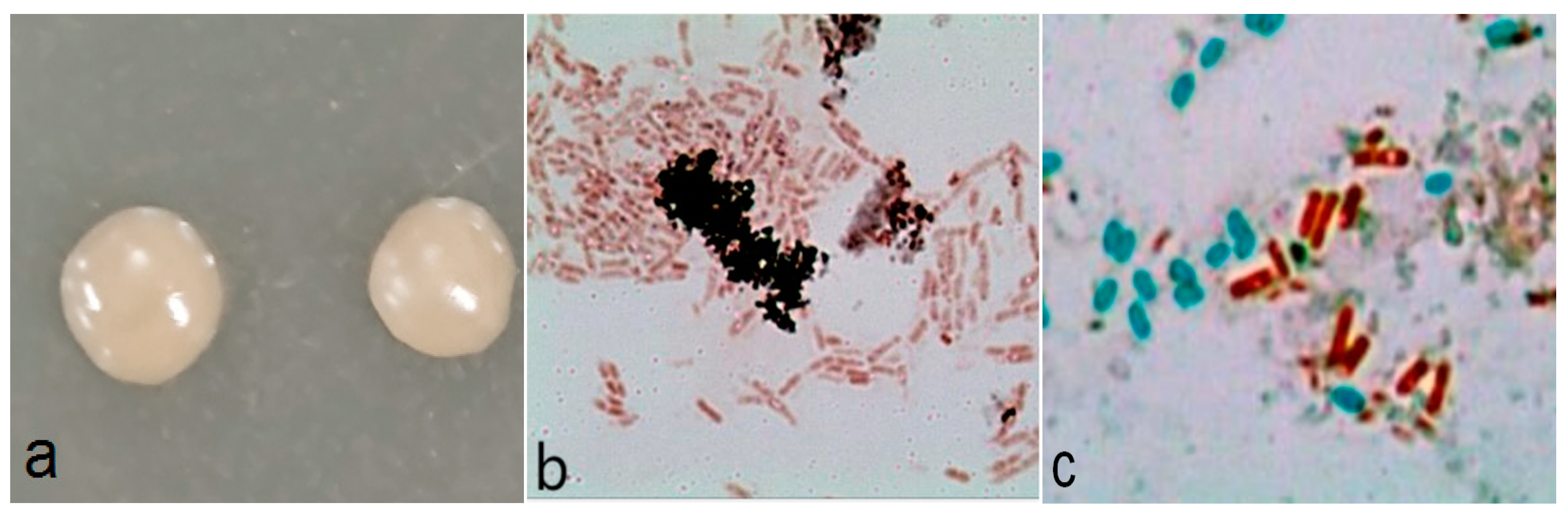
2.4.1. Morphology
2.4.2. Physio-Biochemical Characteristics
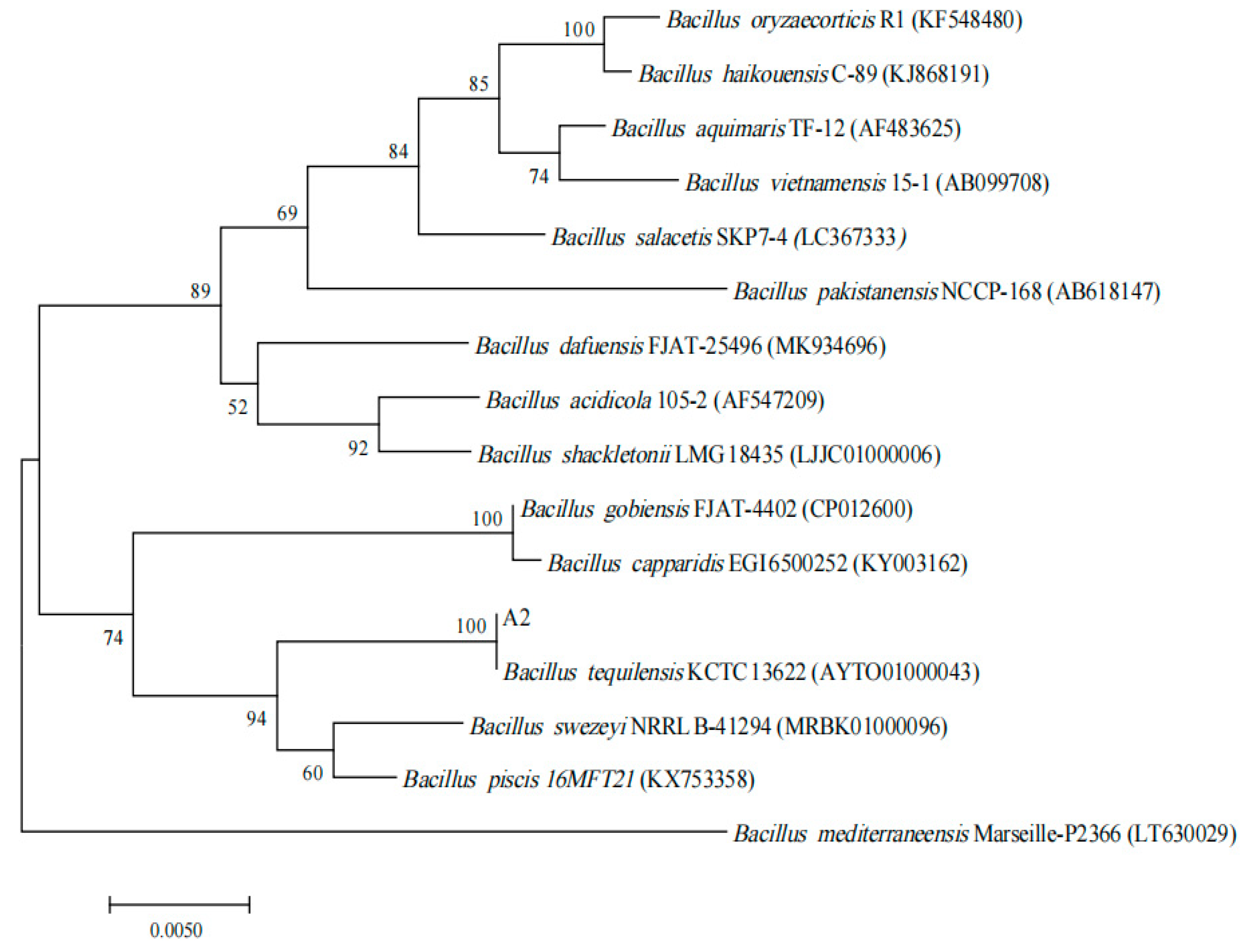
2.4.3. Molecular Identification
2.4.4. In Silico (DDH) Analysis
2.4.5. Fatty Acid Analysis
2.5. Ammonia Nitrogen Degradation Experiment
2.6. Analytical Methods
2.7. Sequence Analysis and Homology Modeling of NiR-A2
2.8. Expression of NiR-A2
2.9. Purification and Enzymatic Properties of the Recombinant Enzyme
3. Results and Discussion
3.1. Identification of A2
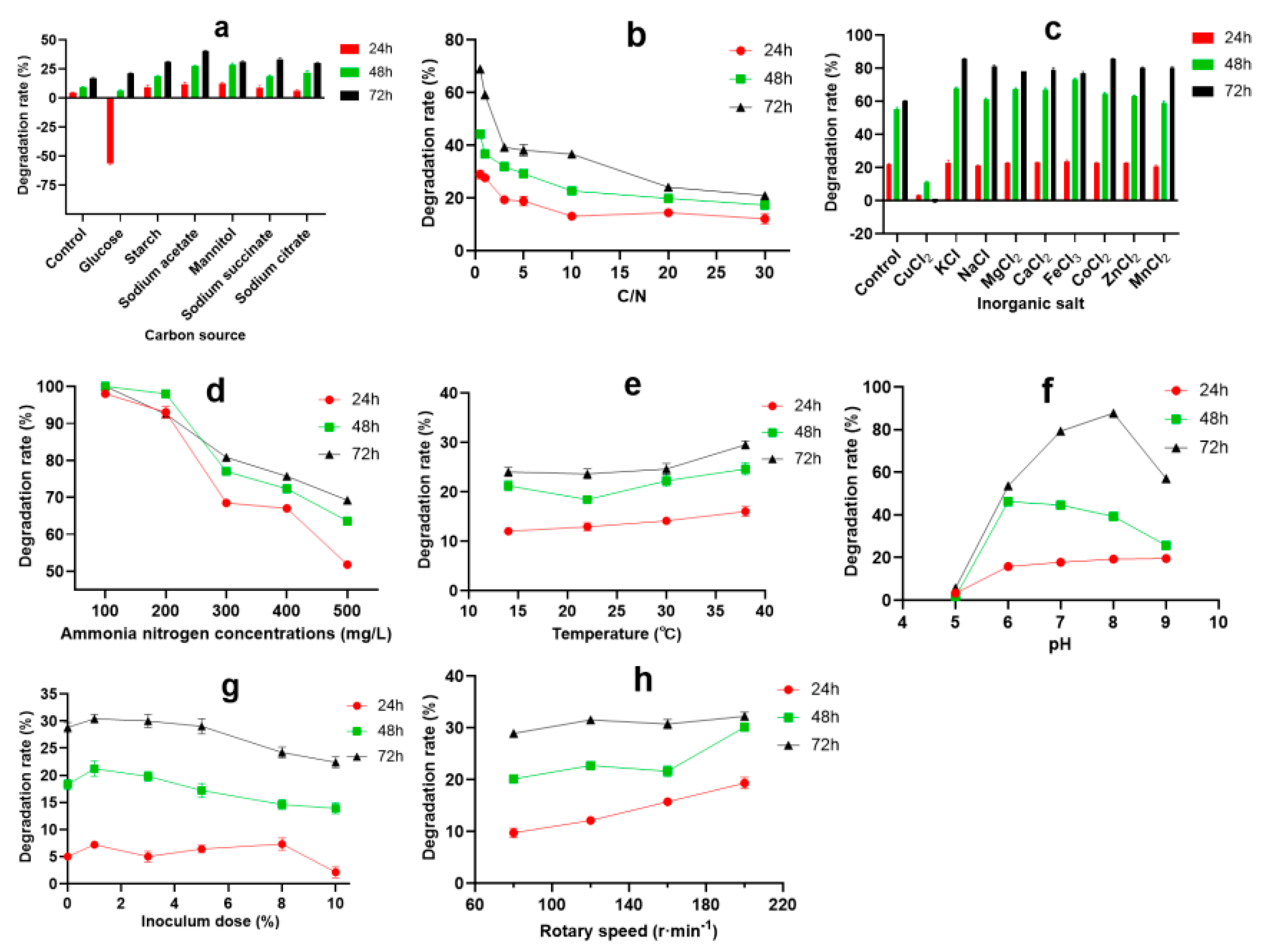
3.2. Optimization of Ammonia Nitrogen Degradation Condition
3.3. Sequence Analysis, Homology Modeling, Expression and Enzyme Characterization of NiR-A2
3.3.1. Sequence Analysis of NiR-A2
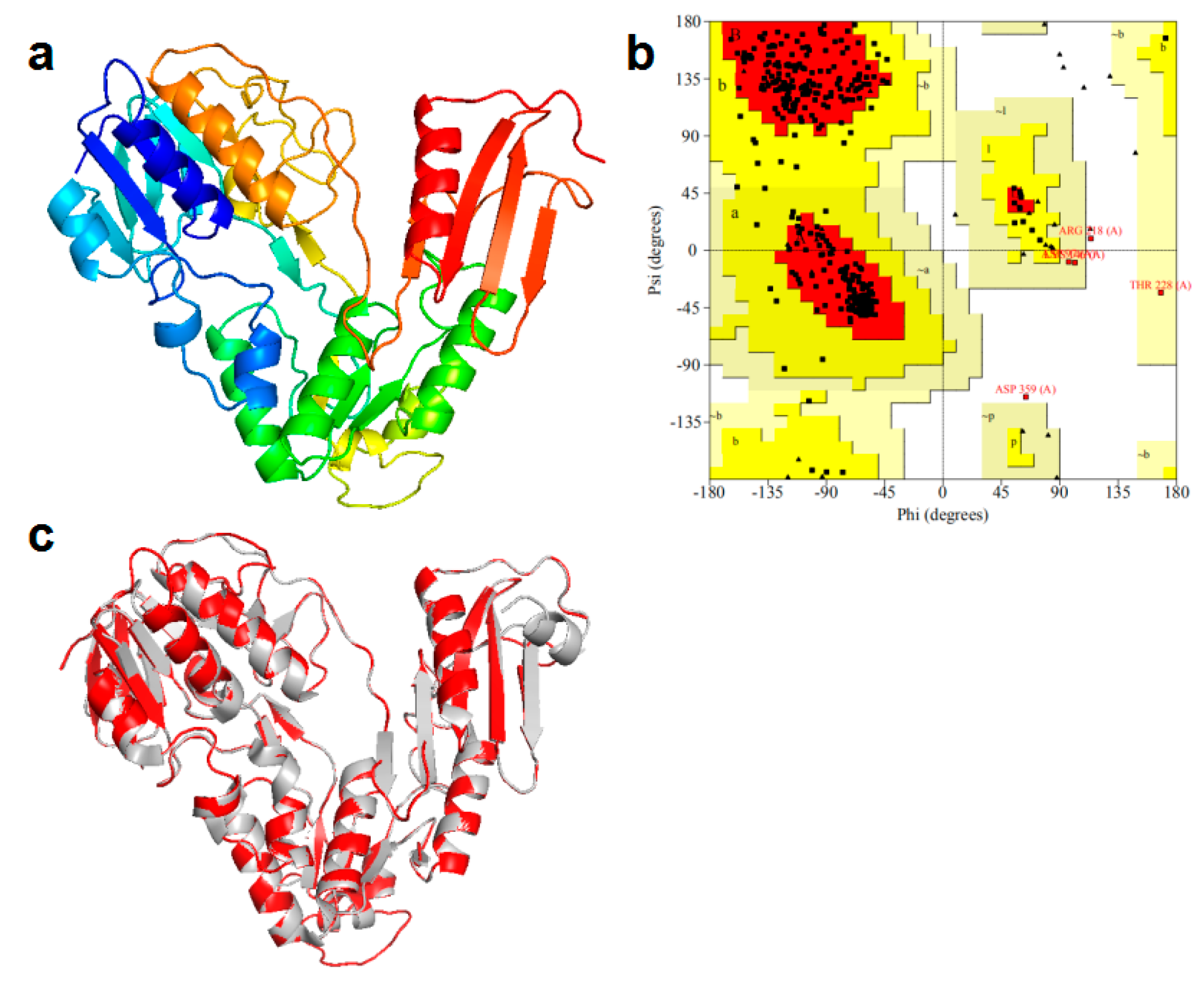
3.3.2. Homology Modeling for NH2-Terminal Amino Acid Sequence of NiR-A2
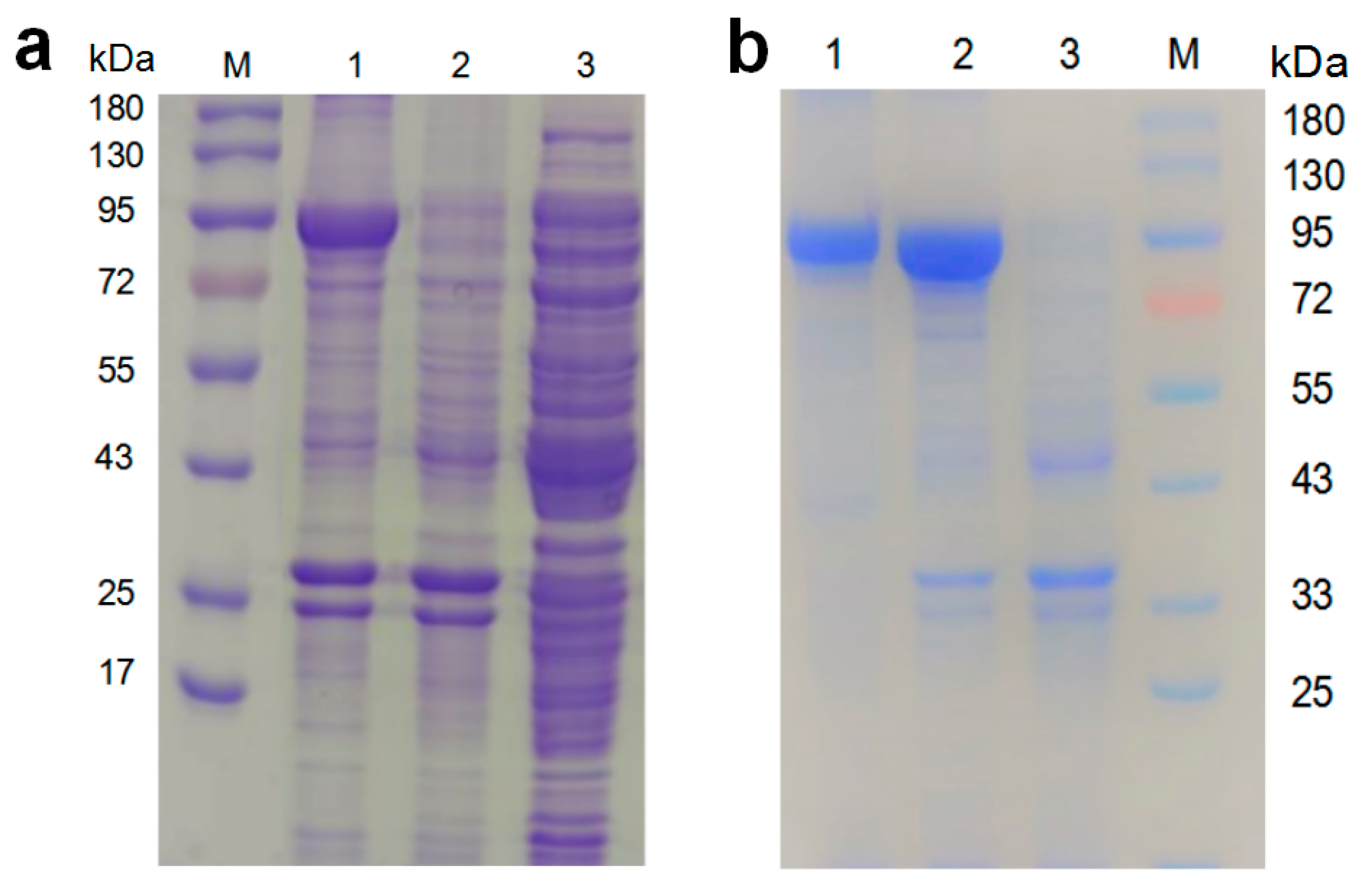
3.3.3. Expression and Purification of NiR-A2
3.3.4. The Biochemical Characterization of Recombinant NiR-A2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salim, H.; Patterson, P.; Ricke, S.; Kim, W. Enhancement of microbial nitrification to reduce ammonia emission from poultry manure: A review. World’s Poult. Sci. J. 2014, 70, 839–856. [Google Scholar] [CrossRef]
- Chen, P.; Li, J.; Li, Q.X.; Wang, Y.; Li, S.; Ren, T.; Wang, L. Simultaneous heterotrophic nitrification and aerobic denitrification by bacterium Rhodococcus sp. CPZ24. Bioresour. Technol. 2012, 116, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Sommer, S.; Hutchings, N. Ammonia emission from field applied manure and its reduction—Invited paper. Eur. J. Agron. 2001, 15, 1–15. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, M.; Qu, J.; Zhang, Y.; Liu, H. Characterization and Mechanism Analysis of Tylosin Biodegradation and Simultaneous Ammonia Nitrogen Removal with Strain Klebsiella pneumoniae TN-1. Bioresour. Technol. 2021, 336, 125342. [Google Scholar] [CrossRef]
- Yun, L.; Yu, Z.; Li, Y.; Luo, P.; Jiang, X.; Tian, Y.; Ding, X. Ammonia Nitrogen and Nitrite Removal by a Heterotrophic Sphingomonas Sp. Strain LPN080 and Its Potential Application in Aquaculture. Aquaculture 2019, 500, 477–484. [Google Scholar] [CrossRef]
- Yi, M.; Wang, H.; Ma, X.; Wang, C.; Wang, M.; Liu, Z.; Lu, M.; Cao, J.; Ke, X. Efficient Nitrogen Removal of a Novel Pseudomonas chengduensis Strain BF6 Mainly through Assimilation in the Recirculating Aquaculture Systems. Bioresour. Technol. 2023, 379, 129036. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xiao, X.; Zhao, Y.; Tu, B.; Zhang, Y. Screening of Efficient Ammonia–Nitrogen Degrading Bacteria and Its Application in Livestock Wastewater. Biomass Convers. Biorefin. 2022, 16, 1–9. [Google Scholar] [CrossRef]
- Zhang, Q.-L.; Liu, Y.; Ai, G.-M.; Miao, L.-L.; Zheng, H.-Y.; Liu, Z.-P. The Characteristics of a Novel Heterotrophic Nitrification-Aerobic Denitrification Bacterium, Bacillus methylotrophicus Strain L7. Bioresour. Technol. 2012, 108, 35–44. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The Evolution and Future of Earth’s Nitrogen Cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef]
- Fukuda, Y.; Tamada, T.; Takami, H.; Suzuki, S.; Inoue, T.; Nojiri, M. Cloning, Expression, Purification, Crystallization and Preliminary X-Ray Crystallographic Study of GK0767, the Copper-Containing Nitrite Reductase from Geobacillus kaustophilus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 692–695. [Google Scholar] [CrossRef]
- Gao, H.; Li, C.; Ramesh, B.; Hu, N. Cloning, Purification and Characterization of Novel Cu-Containing Nitrite Reductase from the Bacillus firmus GY-49. World J. Microbiol. Biotechnol. 2018, 34, 10. [Google Scholar] [CrossRef]
- Huang, Y.; Liang, M.; Zhao, S.; Chen, S.; Liu, J.; Liu, D.; Lu, Y. Isolation, Expression, and Biochemical Characterization: Nitrite Reductase from Bacillus cereus LJ01. RSC Adv. 2020, 10, 37871–37882. [Google Scholar] [CrossRef]
- Wang, Q.; Fu, W.; Lu, R.; Pan, C.; Yi, G.; Zhang, X.; Rao, Z. Characterization of Bacillus subtilis Ab03 for Efficient Ammonia Nitrogen Removal. Syst. Microbiol. Biomanuf. 2022, 2, 580–588. [Google Scholar] [CrossRef]
- Yu, C.-H.; Wang, Y.; Guo, T.; Shen, W.-X.; Gu, M.-X. Isolation and Identification of Ammonia Nitrogen Degradation Strains from Industrial Wastewater. ENG 2012, 04, 790–793. [Google Scholar] [CrossRef]
- Zhang, F.; Xie, F.; Zhou, K.; Zhang, Y.; Zhao, Q.; Song, Z.; Cui, H. Nitrogen Removal Performance of Novel Isolated Bacillus Sp. Capable of Simultaneous Heterotrophic Nitrification and Aerobic Denitrification. Appl. Biochem. Biotechnol. 2022, 194, 3196–3211. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Xie, H.; Yang, E.; Shen, X.; Dai, P.; Zhang, J. Nutrient Removal and Microbial Mechanisms in Constructed Wetland Microcosms Treating High Nitrate/Nitrite Polluted River Water. RSC Adv. 2016, 6, 70848–70854. [Google Scholar] [CrossRef]
- John, E.M.; Krishnapriya, K.; Sankar, T.V. Treatment of Ammonia and Nitrite in Aquaculture Wastewater by an Assembled Bacterial Consortium. Aquaculture 2020, 526, 735390. [Google Scholar] [CrossRef]
- Song, Z.-F.; An, J.; Fu, G.-H.; Yang, X.-L. Isolation and Characterization of an Aerobic Denitrifying Bacillus Sp. YX-6 from Shrimp Culture Ponds. Aquaculture 2011, 319, 188–193. [Google Scholar] [CrossRef]
- Chun, B.H.; Han, D.M.; Kim, K.H.; Jeong, S.E.; Park, D.; Jeon, C.O. Genomic and Metabolic Features of Tetragenococcus halophilus as Revealed by Pan-Genome and Transcriptome Analyses. Food Microbiol. 2019, 83, 36–47. [Google Scholar] [CrossRef]
- Knepp, Z.J.; Ghaner, A.; Root, K.T. Purification and Refolding Protocol for Cold-Active Recombinant Esterase Aa SGNH1 from Aphanizomenon flos-aquae Expressed as Insoluble Inclusion Bodies. Prep. Biochem. Biotechnol. 2022, 52, 394–403. [Google Scholar] [CrossRef]
- Lipničanová, S.; Chmelová, D.; Godány, A.; Ondrejovič, M.; Miertuš, S. Purification of Viral Neuraminidase from Inclusion Bodies Produced by Recombinant Escherichia coli. J. Biotechnol. 2020, 316, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Wintzingerode, F.; Rainey, F.A.; Kroppenstedt, R.M.; Stackebrandt, E. Identification of Environmental Strains of Bacillus Mycoides by Fatty Acid Analysis and Species-Specific 16S RDNA Oligonucleotide Probe. FEMS Microbiol. Ecol. 2006, 24, 201–209. [Google Scholar] [CrossRef]
- Gatti, J.J.; Dobeck, J.M.; Smith, C.; White, R.R.; Socransky, S.S.; Skobe, Z. Bacteria of Asymptomatic Periradicular Endodontic Lesions Identified by DNA-DNA Hybridization. Dent. Traumatol. 2000, 16, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, H.; Lebre, P.; Blom, J.; Cowan, D.; De Maayer, P. Phylogenomic Re-Assessment of the Thermophilic Genus Geobacillus. Syst. Appl. Microbiol. 2016, 39, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, P.; Fry, F.S.; Curtis, S.K.; Al-Khaldi, S.F.; Mossoba, M.M.; Yurawecz, M.P.; Dunkel, V.C. Use of Fatty Acid Profiles to Identify Food-Borne Bacterial Pathogens and Aerobic Endospore-Forming Bacilli. J. Agric. Food Chem. 2005, 53, 3735–3742. [Google Scholar] [CrossRef]
- Yang, X.; Wang, S.; Zhou, L. Effect of Carbon Source, C/N Ratio, Nitrate and Dissolved Oxygen Concentration on Nitrite and Ammonium Production from Denitrification Process by Pseudomonas stutzeri D6. Bioresour. Technol. 2012, 104, 65–72. [Google Scholar] [CrossRef]
- Joo, H.-S.; Hirai, M.; Shoda, M. Characteristics of Ammonium Removal by Heterotrophic Nitrification-Aerobic Denitrification by Alcaligenes faecalis No. 4. J. Biosci. Bioeng. 2005, 100, 184–191. [Google Scholar] [CrossRef]
- Seenivasagan, R.; Kasimani, R.; Babalola, O.O.; Karthika, A.; Rajakumar, S.; Ayyasamy, P.M. Effect of Various Carbon Source, Temperature and PH on Nitrate Reduction Efficiency in Mineral Salt Medium Enriched with Bacillus weinstephnisis (DS45). Groundw. Sustain. Dev. 2017, 5, 21–27. [Google Scholar] [CrossRef]
- Chen, M.; Wang, W.; Feng, Y.; Zhu, X.; Zhou, H.; Tan, Z.; Li, X. Impact Resistance of Different Factors on Ammonia Removal by Heterotrophic Nitrification–Aerobic Denitrification Bacterium Aeromonas Sp. HN-02. Bioresour. Technol. 2014, 167, 456–461. [Google Scholar] [CrossRef]
- Yang, X.-P.; Wang, S.-M.; Zhang, D.-W.; Zhou, L.-X. Isolation and Nitrogen Removal Characteristics of an Aerobic Heterotrophic Nitrifying–Denitrifying Bacterium, Bacillus subtilis A1. Bioresour. Technol. 2011, 102, 854–862. [Google Scholar] [CrossRef]
- Wang, Y.; Zou, Y.-L.; Chen, H.; Lv, Y.-K. Nitrate Removal Performances of a New Aerobic Denitrifier, Acinetobacter haemolyticus ZYL, Isolated from Domestic Wastewater. Bioprocess Biosyst. Eng. 2021, 44, 391–401. [Google Scholar] [CrossRef]
- Mobarry, B.K.; Wagner, M.; Urbain, V.; Rittmann, B.E.; Stahl, D.A. Phylogenetic Probes for Analyzing Abundance and Spatial Organization of Nitrifying Bacteria. Appl. Environ. Microbiol. 1996, 62, 2156–2162. [Google Scholar] [CrossRef]
- Zumft, W.G. The Biological Role of Nitric Oxide in Bacteria. Arch. Microbiol. 1993, 160, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Fan, X.; Li, J.; Wang, X.; Yuan, Z. Isolation and Identification of Naphthalene Degrading Bacteria and Their Degradation Characteristics under Rainwater Environment in Heavily Polluted Areas. J. Environ. Sci. Health Part A 2021, 56, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Izumi, A.; Schnell, R.; Schneider, G. Crystal Structure of NirD, the Small Subunit of the Nitrite Reductase NirbD from Mycobacterium Tuberculosis at 2.0 Å Resolution: NirD from Mycobacterium tuberculosis. Proteins 2012, 80, 2799–2803. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Zhang, D.; Wang, D.; Zhi, Y.; Zhou, P. Heterologous Expression and Biochemical Characterization of Assimilatory Nitrate and Nitrite Reductase Reveals Adaption and Potential of Bacillus megaterium NCT-2 in Secondary Salinization Soil. Int. J. Biol. Macromol. 2017, 101, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, H.; Ibici, H.N.; Erdoğan, E.M.; Türedi, Z.; Ergenekon, P.; Özkan, M. Nitrite is reduced by nitrite reductase NirB without small subunit NirD in Escherichia coli. J. Biosci. Bioeng. 2022, 134, 393–398. [Google Scholar] [CrossRef]
- Ward, D.E.; Donnelly, C.J.; Mullendore, M.E.; van der Oost, J.; de Vos, W.M.; Iii, E.J.C. The NADH Oxidase from Pyrococcus furiosus: Implications for the Protection of Anaerobic Hyperthermophiles against Oxidative Stress. Eur. J. Biochem. 2001, 268, 5816–5823. [Google Scholar] [CrossRef]
- Hirano, J.; Miyamoto, K.; Ohta, H. Purification and Characterization of Thermostable H2O2-Forming NADH Oxidase from 2-Phenylethanol-Assimilating Brevibacterium Sp. KU1309. Appl. Microbiol. Biotechnol. 2008, 80, 71. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Nishikawa, K.; Shomura, Y.; Kawasaki, S.; Niimura, Y.; Higuchi, Y. Crystallization and Preliminary X-Ray Analysis of NADH: Rubredoxin Oxidoreductase from Clostridium acetobutylicum. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Colandene, J.D.; Garrett, R.H. Functional Dissection and Site-Directed Mutagenesis of the Structural Gene for NAD(P)H-Nitrite Reductase in Neurospora crassa. J. Biol. Chem. 1996, 271, 24096–24104. [Google Scholar] [CrossRef] [PubMed]
- García-Lugo, P.; González, C.; Perdomo, G.; Brito, N.; Ávila, J.; de la Rosa, J.M.; Siverio, J.M. Cloning, Sequencing, and Expression OfH.a.YNR1 AndH.a.YNI1, Encoding Nitrate and Nitrite Reductases in the Yeast Hansenula anomala. Yeast 2000, 16, 1099–1105. [Google Scholar] [CrossRef]
- Olmo-Mira, M.F.; Cabello, P.; Pino, C.; Martínez-Luque, M.; Richardson, D.J.; Castillo, F.; Roldán, M.D.; Moreno-Vivián, C. Expression and Characterization of the Assimilatory NADH-Nitrite Reductase from the Phototrophic Bacterium Rhodobacter capsulatus E1F1. Arch. Microbiol. 2006, 186, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Hirai, T.; Arai, H.; Ishii, M.; Igarashi, Y. Purification, Characterization, and Gene Cloning of Thermophilic Cytochrome Cd1 Nitrite Reductase from Hydrogenobacter thermophilus TK-6. J. Biosci. Bioeng. 2006, 101, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Hunter, W.J. Purification and Characterization of Soybean Nodule Nitrite Reductase. Physiol. Plant. 1984, 60, 467–472. [Google Scholar] [CrossRef]
- Ichiki, H.; Tanaka, Y.; Mochizuki, K.; Yoshimatsu, K.; Sakurai, T.; Fujiwara, T. Purification, Characterization, and Genetic Analysis of Cu-Containing Dissimilatory Nitrite Reductase from a Denitrifying Halophilic Archaeon, Haloarcula marismortui. J. Bacteriol. 2001, 183, 4149–4156. [Google Scholar] [CrossRef]
- Lawton, T.J.; Bowen, K.E.; Sayavedra-Soto, L.A.; Arp, D.J.; Rosenzweig, A.C. Characterization of a Nitrite Reductase Involved in Nitrifier Denitrification. J. Biol. Chem. 2013, 288, 25575–25583. [Google Scholar] [CrossRef]
- Song, Q.; Wang, B.; Zhao, F.; Han, Y.; Zhou, Z. Expression, Characterization and Molecular Docking of the Assimilatory NaDH-Nitrite Reductase from Acidovorax wautersii QZ-4. Biochem. Eng. J. 2020, 159, 107589. [Google Scholar] [CrossRef]
- Suzuki, S.; Kataoka, K.; Yamaguchi, K. Metal Coordination and Mechanism of Multicopper Nitrite Reductase. Acc. Chem. Res. 2000, 33, 728–735. [Google Scholar] [CrossRef]
- Zhang, W.; Tian, G.; Feng, S.; Wong, J.H.; Zhao, Y.; Chen, X.; Wang, H.; Ng, T.B. Boletus Edulis Nitrite Reductase Reduces Nitrite Content of Pickles and Mitigates Intoxication in Nitrite-Intoxicated Mice. Sci. Rep. 2015, 5, 14907. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, Y.; Zhou, S.; Kim, S.-W.; Fushinobu, S.; Maruyama, J.; Kitamoto, K.; Wakagi, T.; Shoun, H. A Eukaryotic Copper-Containing Nitrite Reductase Derived from a NirK Homolog Gene of Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2010, 74, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; MacGregor, B.; Teske, A. Identification, Expression and Activity of Candidate Nitrite Reductases From Orange Beggiatoaceae, Guaymas Basin. Front. Microbiol. 2019, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Yang, X.-Y.; Xu, Q.; Cui, H.-L. Characterization of a Novel Cu-Containing Dissimilatory Nitrite Reductase from the Haloarchaeon Halorussus Sp. YCN54. Extremophiles 2020, 24, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, F.; Pistorius, A.; Farkas, D.; De Grip, W.; Hansson, Ö.; Sjölin, L.; Neutze, R. PH Dependence of Copper Geometry, Reduction Potential, and Nitrite Affinity in Nitrite Reductase. J. Biol. Chem. 2007, 282, 6347–6355. [Google Scholar] [CrossRef]
- Matsuoka, M.; Kumar, A.; Muddassar, M.; Matsuyama, A.; Yoshida, M.; Zhang, K.Y.J. Discovery of Fungal Denitrification Inhibitors by Targeting Copper Nitrite Reductase from Fusarium oxysporum. J. Chem. Inf. Model. 2017, 57, 203–213. [Google Scholar] [CrossRef]
- Hira, D.; Toh, H.; Migita, C.T.; Okubo, H.; Nishiyama, T.; Hattori, M.; Furukawa, K.; Fujii, T. Anammox organism KSU-1 Expresses a NirK-Type Copper-Containing Nitrite Reductase Instead of a NirS-Type with Cytochrome Cd1. FEBS Lett. 2012, 586, 1658–1663. [Google Scholar] [CrossRef]
- Gloekner, A.B.; Jiingst, A.; Zumft, W.G. Copper-Containing Nitrite Reductase from Pseudomonas aureofaciens Is Functional in a Mutationally Cytochrome Cdl-Free Background (NirS-) of Pseudomonas stutzeri. Arch. Microbiol. 1993, 160, 18–26. [Google Scholar] [CrossRef]
- Ferroni, F.M.; Guerrero, S.A.; Rizzi, A.C.; Brondino, C.D. Overexpression, Purification, and Biochemical and Spectroscopic Characterization of Copper-Containing Nitrite Reductase from Sinorhizobium meliloti 2011. Study of the Interaction of the Catalytic Copper Center with Nitrite and NO. J. Inorg. Biochem. 2012, 114, 8–14. [Google Scholar] [CrossRef]
- Martí, M.A.; Crespo, A.; Bari, S.E.; Doctorovich, F.A.; Estrin, D.A. QM-MM Study of Nitrite Reduction by Nitrite Reductase of Pseudomonas aeruginosa. J. Phys. Chem. B 2004, 108, 18073–18080. [Google Scholar] [CrossRef]
- Orea, A.; Pajuelo, P.; Pajuelo, E.; Márquez, A.J.; Romero, J.M. Characterisation and Expression Studies of a Root CDNA Encoding for Ferredoxin-Nitrite Reductase from Lotus japonicus. Physiol. Plant. 2001, 113, 193–202. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, B.J.; Biddle, J.F.; Siebert, J.R.; Staunton, E.; Hegg, E.L.; Matthysse, A.G.; Teske, A. Why Orange Guaymas Basin Beggiatoa Spp. Are Orange: Single-Filament-Genome-Enabled Identification of an Abundant Octaheme Cytochrome with Hydroxylamine Oxidase, Hydrazine Oxidase, and Nitrite Reductase Activities. Appl. Environ. Microbiol. 2013, 79, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Zennaro, E.; Ciabatti, I.; Cutruzzola, F.; D’Alessandro, R.; Silvestrini, M.C. The Nitrite Reductase Gene of Pseudomonas Aeruginosa: Effect of Growth Conditions on the Expression and Construction of a Mutant by Gene Disruption. FEMS Microbiol. Lett. 1993, 109, 243–250. [Google Scholar] [CrossRef]
- Shahid, S.; Ali, M.; Legaspi-Humiston, D.; Wilcoxen, J.; Pacheco, A.A. A Kinetic Investigation of the Early Steps in Cytochrome c Nitrite Reductase (CcNiR)-Catalyzed Reduction of Nitrite. Biochemistry 2021, 60, 2098–2115. [Google Scholar] [CrossRef] [PubMed]
- Uppal, S.; Khan, M.A.; Kundu, S. Identification and Characterization of a Recombinant Cognate Hemoglobin Reductase from Synechocystis Sp. PCC 6803. Int. J. Biol. Macromol. 2020, 162, 1054–1063. [Google Scholar] [CrossRef]
- Yamazaki, T.; Oyanagi, H.; Fujiwara, T.; Fukumori, Y. Nitrite Reductase from the Magnetotactic Bacterium Magnetospirillum magnetotacticum. A Novel Cytochrome Cd1 with Fe(II): Nitrite Oxidoreductase Activity. Eur. J. Biochem. 1995, 233, 665–671. [Google Scholar] [CrossRef]
- Zeamari, K.; Gerbaud, G.; Grosse, S.; Fourmond, V.; Chaspoul, F.; Biaso, F.; Arnoux, P.; Sabaty, M.; Pignol, D.; Guigliarelli, B.; et al. Tuning the Redox Properties of a [4Fe-4S] Center to Modulate the Activity of Mo-BisPGD Periplasmic Nitrate Reductase. Biochim. Biophys. Acta BBA-Bioenerg. 2019, 1860, 402–413. [Google Scholar] [CrossRef]
- Sundermeyer-Klinger, H.; Meyer, W.; Warninghoff, B.; Bock, E. Membrane-Bound Nitrite Oxidoreductase of Nitrobacter: Evidence for a Nitrate Reductase System. Arch. Microbiol. 1984, 140, 153–158. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Result | Experiment | Result |
|---|---|---|---|
| Starch hydrolysis | − | Indole production | − |
| Starch fermentation | + | Voges–Proskauer reaction | − |
| Sucrose fermentation | + | Methyl red test | + |
| Glucose fermentation | + | Gelatin hydrolysis | + |
| Salicin fermentation | − | Citrate utilization | + |
| D-fructose fermentation | + | Nitrate reduction | + |
| D-mannose fermentation | − | Catalase | − |
| Cellobiose fermentation | − | Arginine dihydrolase | + |
| Fatty Acid | Percent (%) | Fatty Acid | Percent (%) |
|---|---|---|---|
| 12:0 iso | 0.39 | 16:0 anteiso | 0.25 |
| 12:0 | 0.21 | 16:1 w11c | 0.43 |
| 13:0 iso | 6.39 | Sum In Feature 3 | 7.81 |
| 13:0 anteiso | 0.97 | 16:0 | 4.32 |
| 14:0 iso | 3.53 | 15:0 2OH | 0.77 |
| 14:0 | 3.09 | 17:1 iso w10c | 2.14 |
| 15:0 iso | 35.12 | 17:1 iso w5c | 5.12 |
| 15:0 anteiso | 5.56 | 17:1 anteiso A | 2.18 |
| 15:1 w5c | 0.36 | 17:0 iso | 9.45 |
| 15:0 | - | 17:0 anteiso | 2.02 |
| 16:1 w7c alcohol | 0.53 | 17:0 | 0.17 |
| Sum In Feature 2 | 2.57 | 18:1 w9c | 0.30 |
| 16:0 iso | 5.52 | 18:0 | 0.79 |
| No. | Carbon Source | C/N | Rotary Speed/r·min−1 | Inoculum Dose/% | pH | Degradation Rate/% |
|---|---|---|---|---|---|---|
| 1 | mannitol | 0.5 | 80 | 1% | 6 | 12.4 |
| 2 | mannitol | 1 | 120 | 3% | 7 | 45.9 |
| 3 | mannitol | 5 | 160 | 5% | 8 | 97.5 |
| 4 | mannitol | 10 | 200 | 8% | 9 | 16.9 |
| 5 | sodium acetate | 0.5 | 120 | 5% | 9 | 34.8 |
| 6 | sodium acetate | 1 | 80 | 8% | 8 | 33.2 |
| 7 | sodium acetate | 5 | 200 | 1% | 7 | 43.5 |
| 8 | sodium acetate | 10 | 160 | 3% | 6 | 72.1 |
| 9 | sodium succinate | 0.5 | 160 | 8% | 7 | 54.3 |
| 10 | sodium succinate | 1 | 80 | 5% | 6 | 48.9 |
| 11 | sodium succinate | 5 | 200 | 3% | 9 | 41.1 |
| 12 | sodium succinate | 10 | 120 | 1% | 8 | 54.4 |
| 13 | sodium citrate | 0.5 | 200 | 3% | 8 | 50.7 |
| 14 | sodium citrate | 1 | 160 | 1% | 9 | 33.7 |
| 15 | sodium citrate | 5 | 120 | 8% | 6 | 45.9 |
| 16 | sodium citrate | 10 | 80 | 5% | 7 | 95.0 |
| K1 | 43.2 | 38.1 | 47.4 | 36 | 44.8 | |
| K2 | 45.9 | 40.4 | 45.3 | 52.5 | 59.7 | |
| K3 | 49.7 | 57 | 64.4 | 69.1 | 59 | |
| K4 | 56.3 | 59.6 | 38.1 | 37.6 | 31.6 | |
| Range | 13.1 | 21.5 | 26.3 | 33.1 | 28.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Liu, H.; Cui, T. Identification of a Strain Degrading Ammonia Nitrogen, Optimization of Ammonia Nitrogen Degradation Conditions, and Gene Expression of Key Degrading Enzyme Nitrite Reductase. Fermentation 2023, 9, 397. https://doi.org/10.3390/fermentation9040397
Wang Z, Liu H, Cui T. Identification of a Strain Degrading Ammonia Nitrogen, Optimization of Ammonia Nitrogen Degradation Conditions, and Gene Expression of Key Degrading Enzyme Nitrite Reductase. Fermentation. 2023; 9(4):397. https://doi.org/10.3390/fermentation9040397
Chicago/Turabian StyleWang, Zhenhao, Huijing Liu, and Tangbing Cui. 2023. "Identification of a Strain Degrading Ammonia Nitrogen, Optimization of Ammonia Nitrogen Degradation Conditions, and Gene Expression of Key Degrading Enzyme Nitrite Reductase" Fermentation 9, no. 4: 397. https://doi.org/10.3390/fermentation9040397