
Enhanced Pentostatin Production in Actinomadura sp. by Combining ARTP Mutagenesis, Ribosome Engineering and Subsequent Fermentation Optimization


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains and Culture Conditions
2.3. Preparation of Single-Spore Suspension
2.4. Random Mutagenesis by ARTP
2.5. Resistant Screening of Actinomadura sp. ATCC 39365-Derived Mutants
2.6. Efficient Screening of Resistant Mutants with High-Yield Pentostatin by HPLC-MS/MS
2.7. Verification of Genetic Stability of the Mutant with High-Yield Pentostatin
2.8. Determination of Biomass
2.9. qRT-PCR Analysis of Key Genes in the Biosynthesis Gene Cluster of Pentostatin
2.10. Fermentation Optimization of the Mutant with High-Yield Pentostatin
3. Results
3.1. Optimal ARTP Exposure Time on Actinomadura sp. ATCC 39365 Mutagenesis
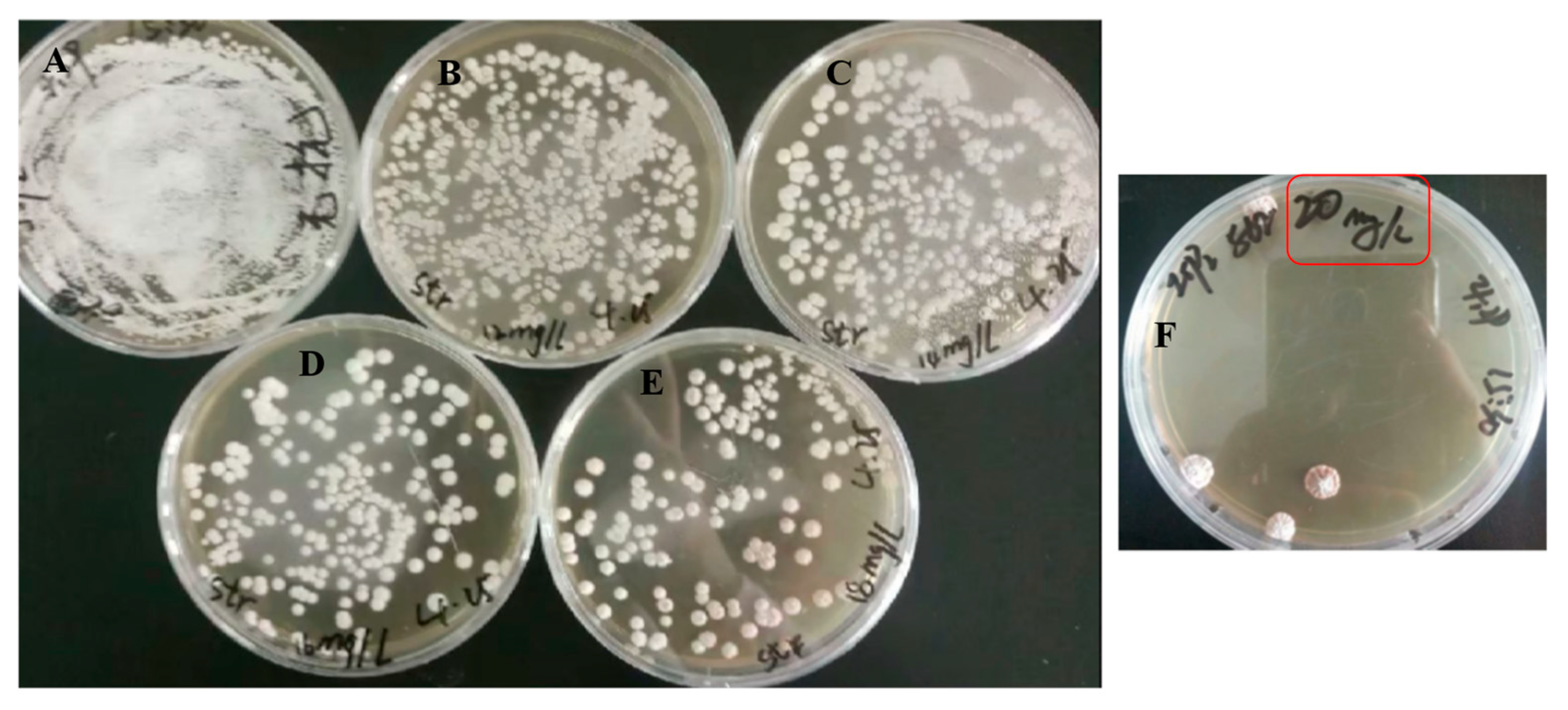
3.2. Screening of High-Yield Pentostatin Mutants with Streptomycin
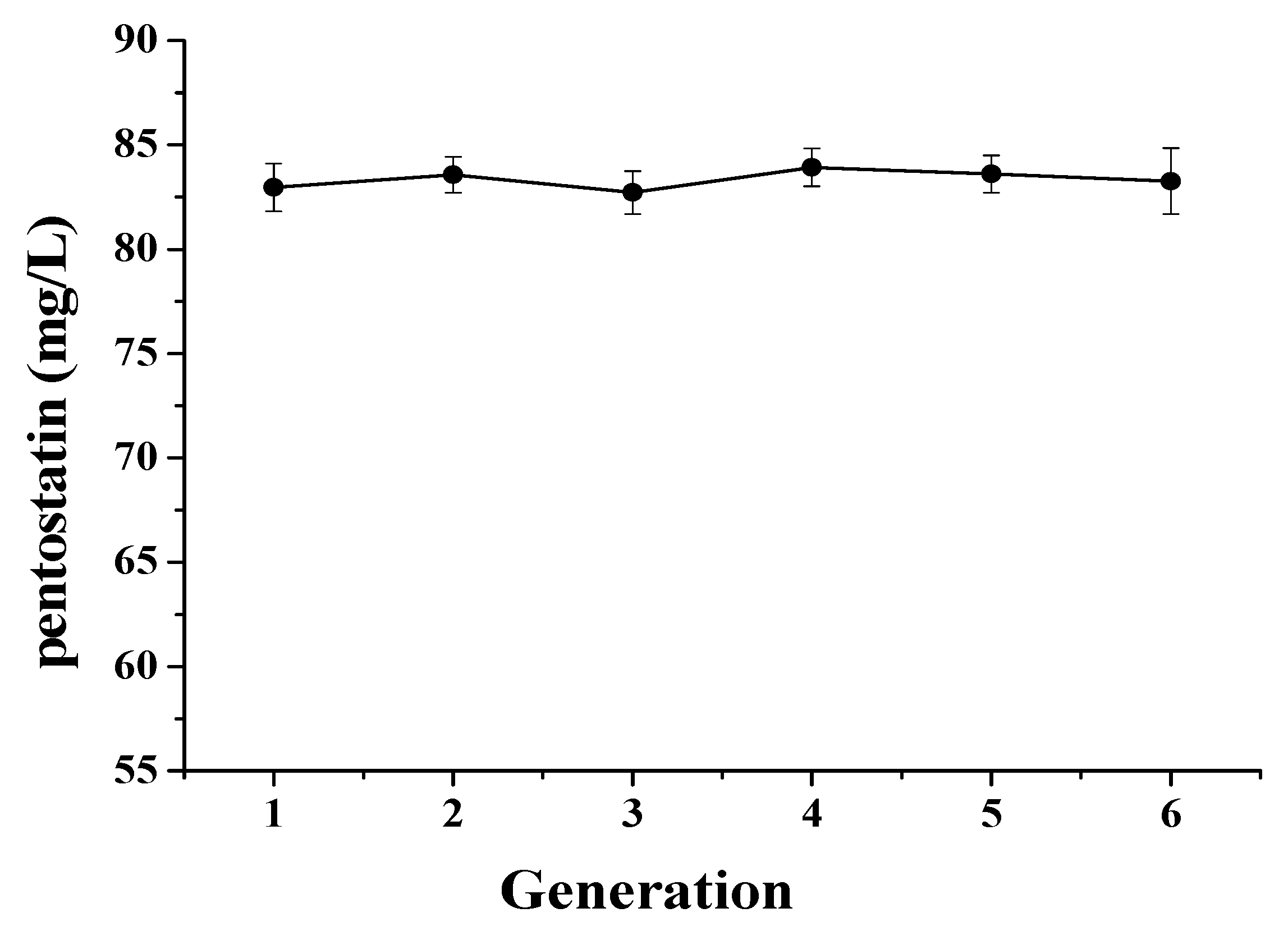
3.3. Genetic Stability of Mutant Actinomadura sp. S-15
3.4. Transcription Levels of Key Genes Related to the Biosynthesis of Pentostatin
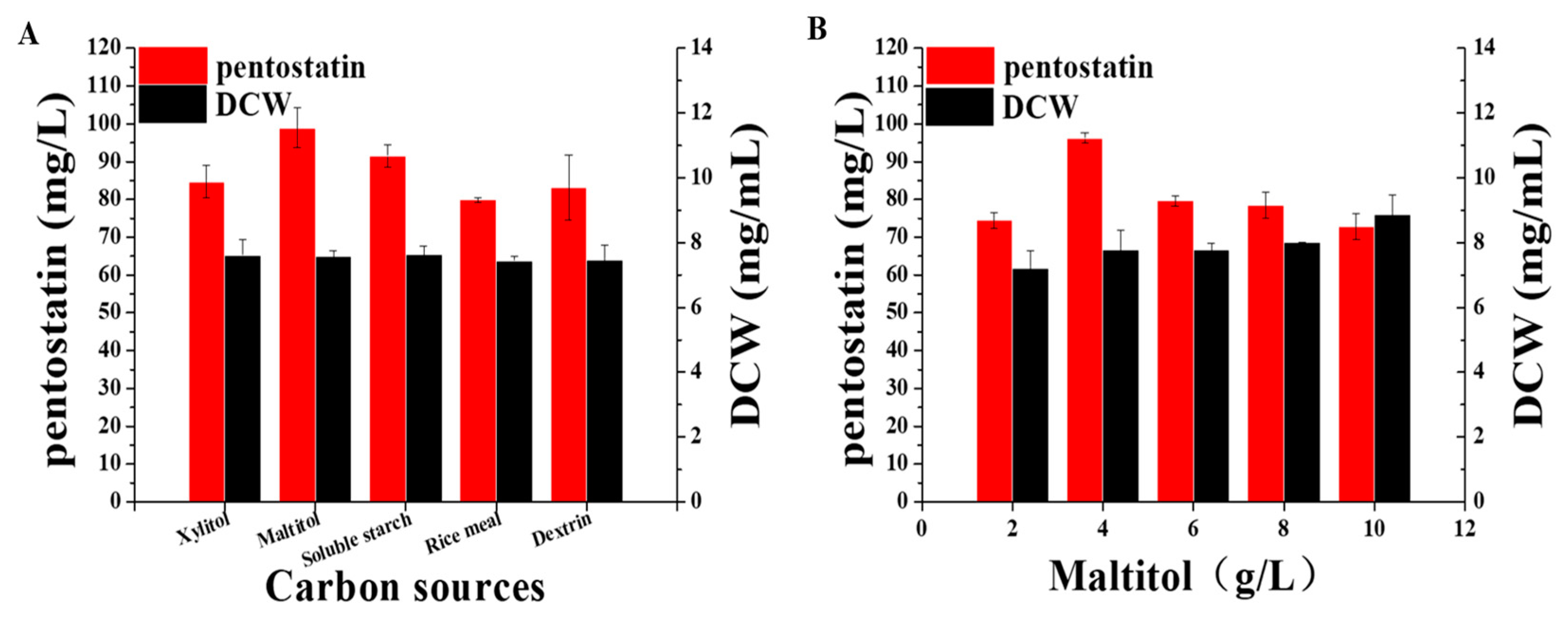
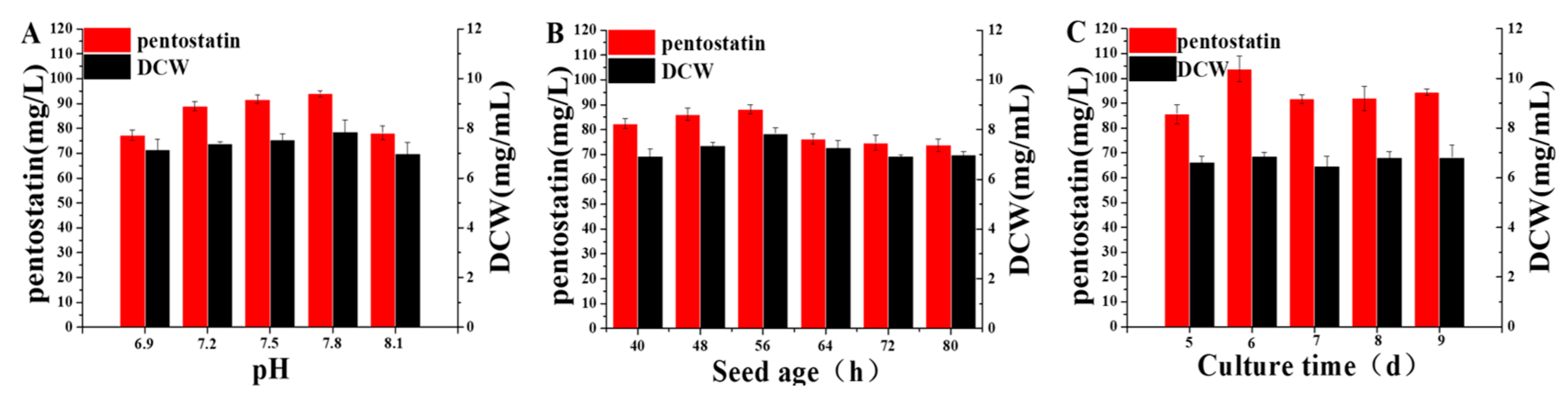
3.5. Optimization of Fermentation Conditions of Actinomadura sp. S-15
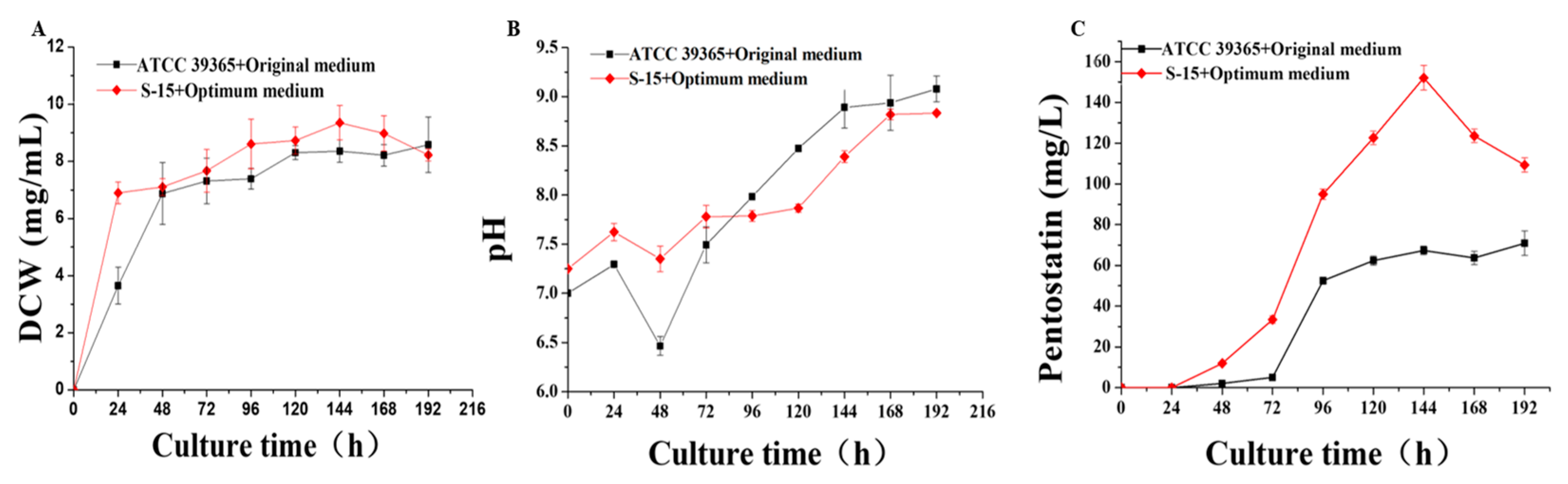
3.6. Difference in Fermentation Performance between Actinomadura sp. S-15 and Actinomadura sp. ATCC 39365
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gozzetti, A.; Sammartano, V.; Bacchiarri, F.; Raspadori, D.; Bocchia, M. A BRAF-Negative Classic Hairy Cell Leukemia Patient with Long-Lasting Complete Remission after Rituximab and Pentostatin. Turk. J. Hematol. 2020, 37, 286–287. [Google Scholar] [CrossRef]
- Ho, A.D.; Hensel, M. Pentostatin for the treatment of indolent lymphoproliferative disorders. Semin. Hematol. 2006, 43, S2–S10. [Google Scholar] [CrossRef]
- Iannitto, E.; Tripodo, C. How I Diagnose and Treat Splenic Lymphomas. Blood 2011, 117, 2585–2595. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, G.Y.; Li, C.Y.; Ling, J.Y. Cordycepin and Pentostatin Biosynthesis Gene Identified through Transcriptome and Proteomics Analysis of Cordyceps kyushuensis Kob. Microbiol. Res. 2019, 218, 12–21. [Google Scholar] [CrossRef]
- Xia, Y.L.; Luo, F.F.; Shang, Y.F.; Chen, P.L.; Lu, Y.Z.; Wang, C.S. Fungal Cordycepin Biosynthesis is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489. [Google Scholar] [CrossRef]
- Wu, P.; Wan, D.; Xu, G.; Wang, G.; Ma, H.; Wang, T.; Gao, Y.; Qi, J.; Chen, X.; Zhu, J.; et al. An Unusual Protector-Protege Strategy for the Biosynthesis of Purine Nucleoside Antibiotics. Cell Chem. Biol. 2017, 24, 171–181. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, G.; Wu, P.; Liu, J.; Cai, Y.S.; Deng, Z.; Chen, W. Biosynthesis of 2′-Chloropentostatin and 2′-Amino-2′-Deoxyadenosine Highlights a Single Gene Cluster Responsible for Two Independent Pathways in Actinomadura sp. Strain ATCC 39365. Appl. Environ. Microbiol. 2017, 83, e00078–e00087. [Google Scholar] [CrossRef]
- Kodama, K.; Kusakabe, H.; Machida, H.; Midorikawa, Y.; Shibuya, S.; Kuninaka, A.; Yoshino, H. Isolation of 2′-Deoxycoformycin and Cordycepin from Wheat Bran Culture of Aspergillus nidulans Y 176–2. Agric. Biol. Chem. 1979, 43, 2375–2377. [Google Scholar] [CrossRef]
- Truong, T.-V.; Rapoport, H. Chirospecific synthesis of the tetrahydroimidazodiaepinol aglycon of pentostatin and its analogues. J. Org. Chem. 1993, 58, 6090–6096. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, R.; Lou, T.; Zhao, P.; Wang, S.-Y. Pentostatin Biosynthesis Pathway Elucidation and Its Application. Fermentation 2022, 8, 459. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, Y.; Jiang, S.X.; Cai, X.; Huang, K.; Liu, Z.Q.; Zheng, Y.G. Comparative Metabolomics Analysis of Amphotericin B High-yield Mechanism for Metabolic Engineering. Microb. Cell Fact. 2021, 20, 66. [Google Scholar] [CrossRef]
- Yin, S.; Wang, W.; Wang, X.; Zhu, Y.; Jia, X.; Li, S.; Yuan, F.; Zhang, Y.; Yang, K. Identification of a Cluster-situated Activator of Oxytetracycline Biosynthesis and Manipulation of Its Expression for Improved Oxytetracycline Production in Streptomyces rimosus. Microb. Cell Fact. 2015, 14, 46. [Google Scholar] [CrossRef]
- Koshla, O.; Lopatniuk, M.; Borys, O.; Misaki, Y.; Kravets, V.; Ostash, I.; Shemediuk, A.; Ochi, K.; Luzhetskyy, A.; Fedorenko, V.; et al. Genetically Engineered rpsL Merodiploidy Impacts Secondary Metabolism and Antibiotic Resistance in Streptomyces. World J. Microbiol. Biotechnol. 2021, 37, 62. [Google Scholar] [CrossRef]
- Zhang, K.; Mohsin, A.; Dai, Y.; Chen, Z.; Guo, M. Combinatorial Effect of ARTP Mutagenesis and Ribosome Engineering on an Industrial Strain of Streptomyces albus S12 for Enhanced Biosynthesis of Salinomycin. Front. Bioeng. Biotechnol. 2019, 7, 212. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-W.; Ge, X.-Y.; Huang, Y.; Chai, X.-T.; Zhang, L.; Zhang, Y.-X.; Deng, L.-N.; Liu, C.-Q.; Xu, H.; Gao, J. High-yield Strain of Fusidic Acid Obtained by Atmospheric and Room Temperature Plasma Mutagenesis and the Transcriptional Changes Involved in Improving Its Production in Fungus Fusidium coccineum. J. Appl. Microbiol. 2021, 130, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Song, Z.; Ma, Z.; Bechthold, A.; Yu, X. Sequential Improvement of Rimocidin Production in Streptomyces rimosus M527 by Introduction of Cumulative Drug-resistance Mutations. J. Ind. Microbiol. Biotechnol. 2019, 46, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Funane, K.; Tanaka, Y.; Hosaka, T.; Murakami, K.; Miyazaki, T.; Shiwa, Y.; Gibu, S.; Inaoka, T.; Kasahara, K.; Fujita, N.; et al. Combined Drug-Resistance Mutations Substantially Enhance Enzyme Production in Paenibacillus agaridevorans. J. Bacteriol. 2018, 200, e00188-18. [Google Scholar] [CrossRef]
- Wang, X.-B.; Lu, M.-S.; Wang, S.-J.; Fang, Y.-W.; Wang, D.-L.; Ren, W.; Zhao, G.-M. The Atmospheric and Room Temperature Plasma (ARTP) Method on the Dextranase Activity and Structure. Int. J. Biol. Macromol. 2014, 70, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yang, H.; Chen, X.; Sun, B.; Du, G.; Zhou, Z.; Song, J.; Fan, Y.; Shen, W. Significantly Improving the Yield of Recombinant Proteins in Bacillus subtilis by a Novel Powerful Mutagenesis Tool (ARTP): Alkaline α-amylase as a Case Study. Protein Expr. Purif. 2015, 114, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Feng, L.R.; Wei, L.; Li, H.G.; Wang, L.; Zhou, Y.; Yu, X.B. Mutation Breeding of Lycopene-Producing Strain Blakeslea Trispora by a Novel Atmospheric and Room Temperature Plasma (ARTP). Appl. Biochem. Biotechnol. 2014, 174, 452–460. [Google Scholar]
- Choi, J.I.; Yoon, M.; Joe, M.; Park, H.; Lee, S.G.; Han, S.J.; Lee, P.C. Development of Microalga Scenedesmus Dimorphus Mutant with Higher Lipid Content by Radiation Breeding. Bioprocess Biosyst. Eng. 2014, 37, 2437–2444. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Nie, S.-X.; Hu, S.-J.; Wang, S.-J.; Wang, J.-J.; Guo, K. Screening of Beauveria bassiana with High Biocontrol Potential Based on ARTP Mutagenesis and High-throughput FACS. Pestic. Biochem. Physiol. 2021, 171, 104732. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Liu, Y.; Lin, H.; Liu, C. Improving Vanadium Extraction from Stone Coal via Combination of Blank Roasting and Bioleaching by ARTP-mutated Bacillus mucilaginosus. Trans. Nonferrous Met. Soc. 2019, 29, 849–858. [Google Scholar] [CrossRef]
- Yang, M.M.; An, Y.F.; Zabed, H.M.; Guoa, Q.; Yun, J.H.; Zhang, G.Y.; Awad, F.N.; Sun, W.J.; Qi, X.H. Random Mutagenesis of Clostridium butyricum Strain and Optimization of Biosynthesis Process for Enhanced Production of 1,3-propanediol. Bioresour. Technol. 2019, 284, 188–196. [Google Scholar] [CrossRef]
- Wei, L.; Mao, Y.; Liu, H.; Ke, C.; Liu, X.; Li, S. Effect of an Inorganic Nitrogen Source (NH4)2SO4 on the Production of Welan gum from Sphingomonas sp. Mutant Obtained through UV-ARTP Compound Mutagenesis. Int. J. Biol. Macromol. 2022, 210, 630–638. [Google Scholar] [CrossRef]
- Yun, J.; Zabed, H.-M.; Zhang, Y.; Zhang, G.; Zhao, M.; Qi, X. Improving Tolerance and 1,3-propanediol Production of Clostridium butyricum Using Physical Mutagenesis, Adaptive Evolution and Genome Shuffling. Bioresour. Technol. 2022, 363, 127967. [Google Scholar] [CrossRef]
- Ochi, K.; Okamoto, S.; Tozawa, Y.; Inaoka, T.; Hosaka, T.; Xu, J.; Kurosawa, K. Ribosome Engineering and Secondary Metabolite Production. Adv. Appl. Microbiol. 2004, 56, 155–184. [Google Scholar]
- Ochi, K.; Okamoto, S. New Development to Utilize Streptomyces by “Ribosome Engineering”. Nippon. Nogeikagaku Kaishi J. Agric. Chem. Soc. Jpn. 2004, 78, 1082–1085. [Google Scholar] [CrossRef]
- Okamoto-Hosoya, Y.; Sato, T.A.; Ochi, K. Resistance to Paromomycin is Conferred by rpsL Mutations, Accompanied by an Enhanced Antibiotic Production in Streptomyces coelicolor A3(2). J. Antibiot. 2000, 53, 1424–1427. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.H.; Ying, Y.H.; Kim, H.J.; Koh, Y.H.; Kim, C.J.; Lee, S.H.; Cha, C.Y.; Kook, Y.H.; Kim, B.J. Mechanism of Natural Rifampin Resistance of Streptomyces spp. Syst. Appl. Microbiol. 2005, 28, 398–404. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Dhakal, D.; Pham, V.T.; Nguyen, H.T.; Sohng, J.K. Recent Advances in Strategies for Activation and Discovery/Characterization of Cryptic Biosynthetic Gene Clusters in Streptomyces. Microorganisms 2020, 8, 616. [Google Scholar] [CrossRef]
- Zhu, S.B.; Duan, Y.W.; Huang, Y. The Application of Ribosome Engineering to Natural Product Discovery and Yield Improvement in Streptomyces. Antibiotics 2019, 8, 133. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Kasahara, K.; Izawa, M.; Ochi, K. Applicability of Ribosome Engineering to Vitamin B12 Production by Propionibacterium shermanii. Biosci. Biotechnol. Biochem. 2017, 81, 1636–1641. [Google Scholar] [CrossRef]
- Lv, X.A.; Jin, Y.Y.; Li, Y.D.; Zhang, H.; Liang, X.L. Genome Shuffling of Streptomyces viridochromogenes for Improved Production of Avilamycin. Appl. Microbiol. Biotechnol. 2013, 97, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Hui, M.; Li, R.; Chen, L.; Tian, H.; Wang, L. Enhancement of Daptomycin Production by the Method of Combining Ribosome Engineering and Genome Shuffling in Streptomyces roseosporus. Appl. Biochem. Microbiol. 2018, 54, 611–615. [Google Scholar] [CrossRef]
- Liu, H.M.; Jiang, C.Z.; Lin, J.; Zhuang, Z.K.; Kong, W.P.; Liu, L.; Huang, Y.; Duan, Y.W.; Zhu, X.C. Genome Shuffling Based on Different Types of Ribosome Engineering Mutants for Enhanced Production of 10-membered Enediyne tiancimycin-A. Appl. Microbiol. Biotechnol. 2020, 104, 4359–4369. [Google Scholar] [CrossRef]
- Zhu, X.C.; Kong, J.Q.; Yang, H.; Huang, R.; Huang, Y.; Yang, D.; Shen, B.; Duan, Y.W. Strain Improvement by Combined UV Mutagenesis and Ribosome Engineering and Subsequent Fermentation Optimization for Enhanced 6′-deoxy-bleomycin Z Production. Appl. Microbiol. Biotechnol. 2018, 102, 1651–1661. [Google Scholar] [CrossRef]
- Tong, Q.-Q.; Zhou, Y.-H.; Chen, X.-S.; Wu, J.-Y.; Wei, P.; Yuan, L.-X.; Yao, J.-M. Genome Shuffling and Ribosome Engineering of Streptomyces virginiae for Improved Virginiamycin Production. Bioprocess Biosyst. Eng. 2018, 41, 729–738. [Google Scholar] [CrossRef]
- Li, Y.M.; Li, J.Y.; Ye, Z.M.; Lu, L.C. Enhancement of Angucycline Production by Combined UV mutagenesis and Ribosome Engineering and Fermentation Optimization in Streptomyces dengpaensis XZHG99(T). Prep. Biochem. Biotechnol. 2021, 51, 173–182. [Google Scholar] [CrossRef]
- Ochi, K. Insights into Microbial Cryptic Gene Activation and Strain Improvement: Principle, Application and Technical Aspects. J. Antibiot. 2017, 70, 25–40. [Google Scholar] [CrossRef]
- Lee, J.; Kim, D.-S.; Jewett, M.-C. 6.17-Recent Advances in Engineering Ribosomes for Natural Product Biosynthesis. Compr. Nat. Prod. III 2020, 6, 377–397. [Google Scholar]
- Zhao, M.; Zhang, H.; Lou, T.; Zhao, K.; Wang, S. Simultaneous Determination of Pentostatin and 2′-amino-2′-deoxyadenosine in Fermentation Broth by High Performance Liquid Chromatography-tandem Mass Spectrometry. Se Pu 2021, 39, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Y.; Yang, S.-B.; Wu, Y.-J.; Shen, X.-F.; Chen, S.-X. Enhancement of A82846B Yield and Proportion by Overexpressing the Halogenase Gene in Amycolatopsis orientalis SIPI18099. Appl. Microbiol. Biotechnol. 2018, 102, 5635–5643. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.-J.; Schmittgen, T.-D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ma, X.-J.; Zhang, H.-M.; Lu, X.-F.; Han, J.; Zhu, H.-X.; Wang, H.; Yao, R.-S. Mutant Breeding of Starmerella bombicola by Atmospheric and Room-temperature Plasma (ARTP) for Improved Production of Specific or Total Sophorolipids. Bioprocess Biosyst. Eng. 2020, 43, 1869–1883. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (s) | Flow Rate (mL/min) | φ(A)/% | φ(B)/% |
|---|---|---|---|
| 0 | 0.3 | 95 | 5 |
| 120 | 0.3 | 95 | 5 |
| 360 | 0.3 | 5 | 95 |
| 180 | 0.3 | 0 | 100 |
| 360 | 0.3 | 95 | 5 |
| Target Genes | Primer Sequence | Tm (°C) |
|---|---|---|
| 16S rRNA-F | 5′-GAGCGAACAGGATTAGATACCC-3′ | 57.6 |
| 16S rRNA-R | 5′-TCCTTTGAGTTTTAGCCTTGC-3′ | 57.2 |
| Ade A-F | 5′-GACTACCGGACCGACCCC-3′ | 59.4 |
| Ade A-R | 5′-GCCGAAGATGAGGCAGAAGT-3′ | 59.2 |
| Ade B-F | 5′-GTCATCACATGAGACCCCGG-3′ | 60.2 |
| Ade B-R | 5′-GAGCGGTTCAGGGAGAAGAC-3′ | 60.1 |
| Ade C-F | 5′-GAGATCCCCCTGGTCGTC-3′ | 57.0 |
| Ade C-R | 5′-GCTCGGTGTTCCTGGAGTAG-3′ | 58.0 |
| Ade K-F | 5′-ATCTCCAGCCTGGACGACCT-3′ | 62.5 |
| Ade K-R | 5′-CTAGGGCTCCGGTTTGCGTA-3′ | 62.0 |
| Ade M-F | 5′-CTCATGGTCCAGCTCGAAGG-3′ | 60.2 |
| Ade M-R | 5′-ATGACCTCGCAGAACACGTC-3′ | 60.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zhang, D.; Liu, R.; Lou, T.; Tan, R.; Wang, S. Enhanced Pentostatin Production in Actinomadura sp. by Combining ARTP Mutagenesis, Ribosome Engineering and Subsequent Fermentation Optimization. Fermentation 2023, 9, 398. https://doi.org/10.3390/fermentation9040398
Zhang H, Zhang D, Liu R, Lou T, Tan R, Wang S. Enhanced Pentostatin Production in Actinomadura sp. by Combining ARTP Mutagenesis, Ribosome Engineering and Subsequent Fermentation Optimization. Fermentation. 2023; 9(4):398. https://doi.org/10.3390/fermentation9040398
Chicago/Turabian StyleZhang, Hongyu, Deguang Zhang, Ran Liu, Tingting Lou, Ruyue Tan, and Suying Wang. 2023. "Enhanced Pentostatin Production in Actinomadura sp. by Combining ARTP Mutagenesis, Ribosome Engineering and Subsequent Fermentation Optimization" Fermentation 9, no. 4: 398. https://doi.org/10.3390/fermentation9040398