
Transcriptomics-Guided Investigation of the SLCG_Lrp Regulon Provides New Insights into Its Role for Lincomycin Biosynthesis
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Strains, Plasmids along with Culture
2.2. Gene Deletion and Overexpression Mutant Establishment
2.3. Fermentation and Lincomycin Determination
2.4. RNA-Seq
2.5. RNA Extraction and RT-qPCR Analysis
2.6. Protein Expression and Purification
2.7. Electrophoretic Mobility Shift Assays (EMSAs)
2.8. Statistical Analysis
3. Results
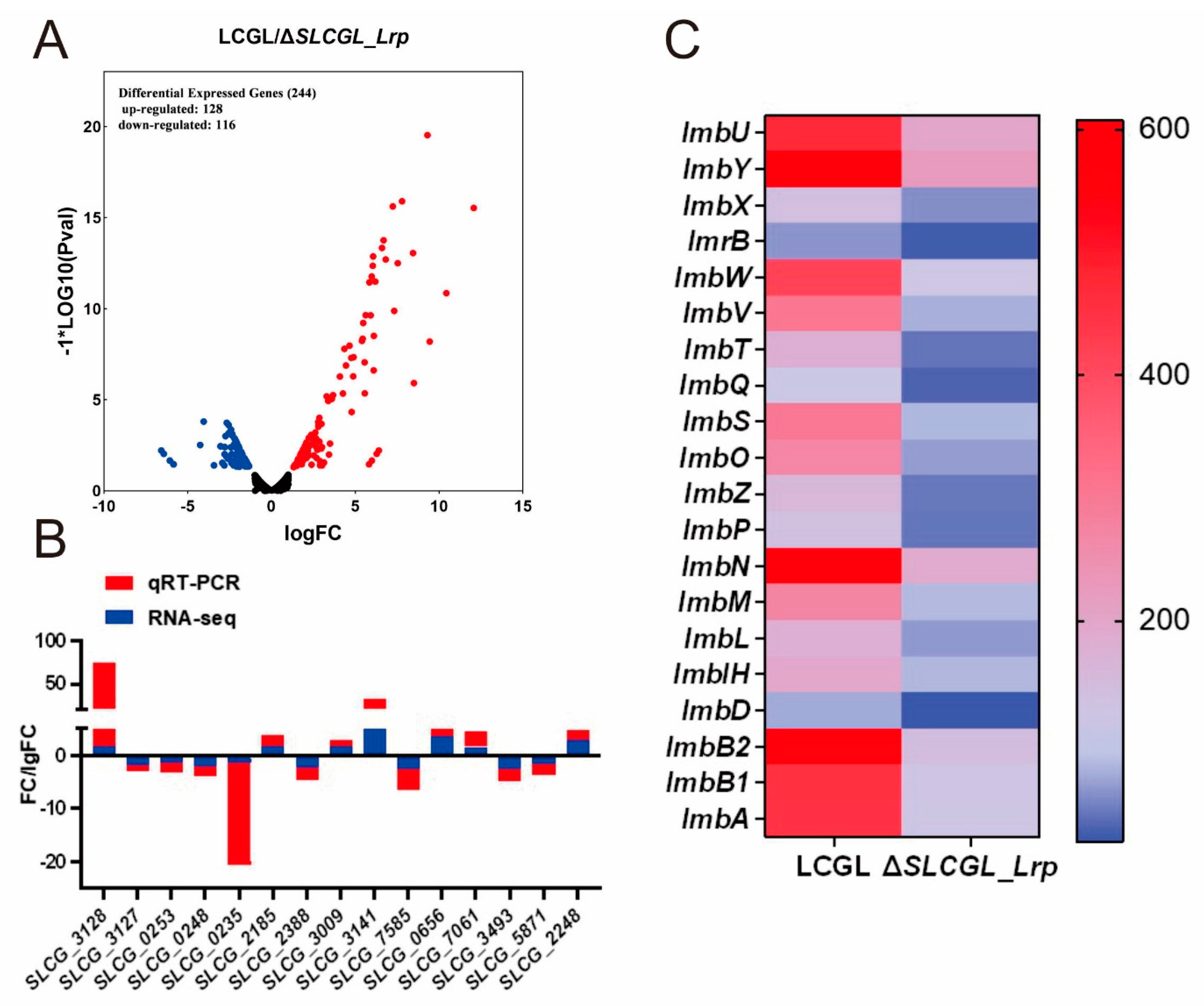
3.1. Transcriptome Analysis of LCGL and ΔSLCGL_Lrp
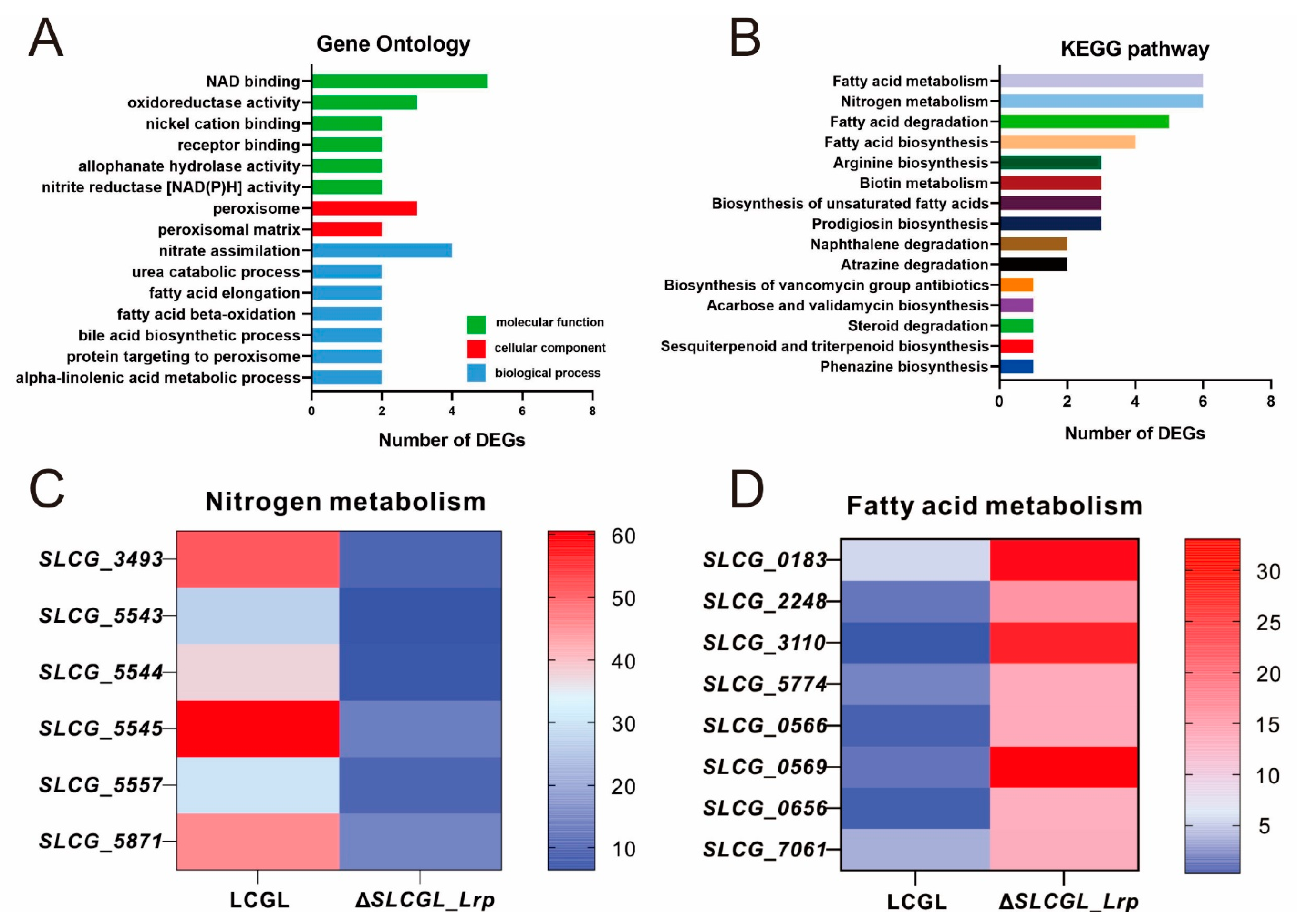
3.2. GO as Well as KEGG Analysis on DEGs
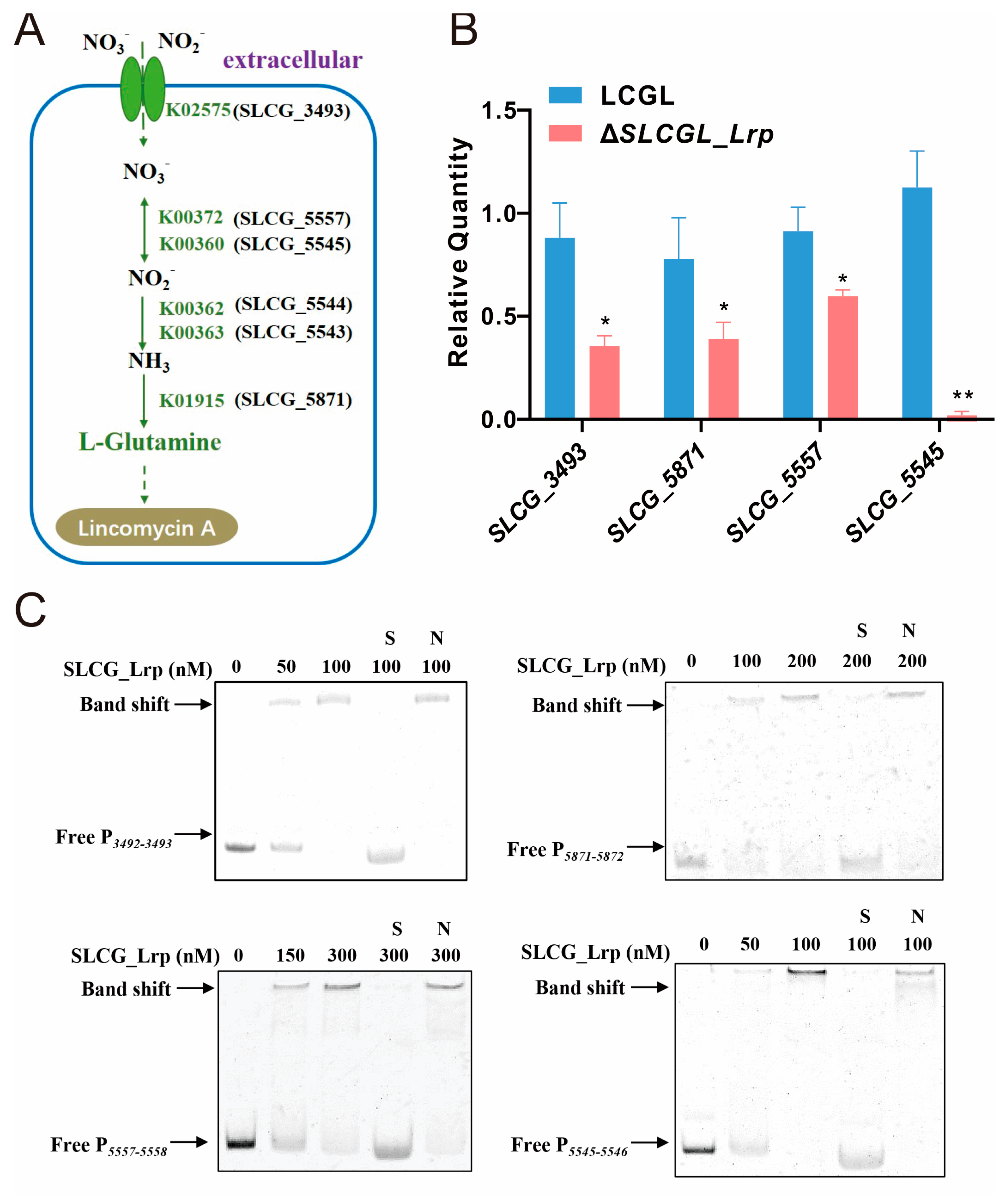
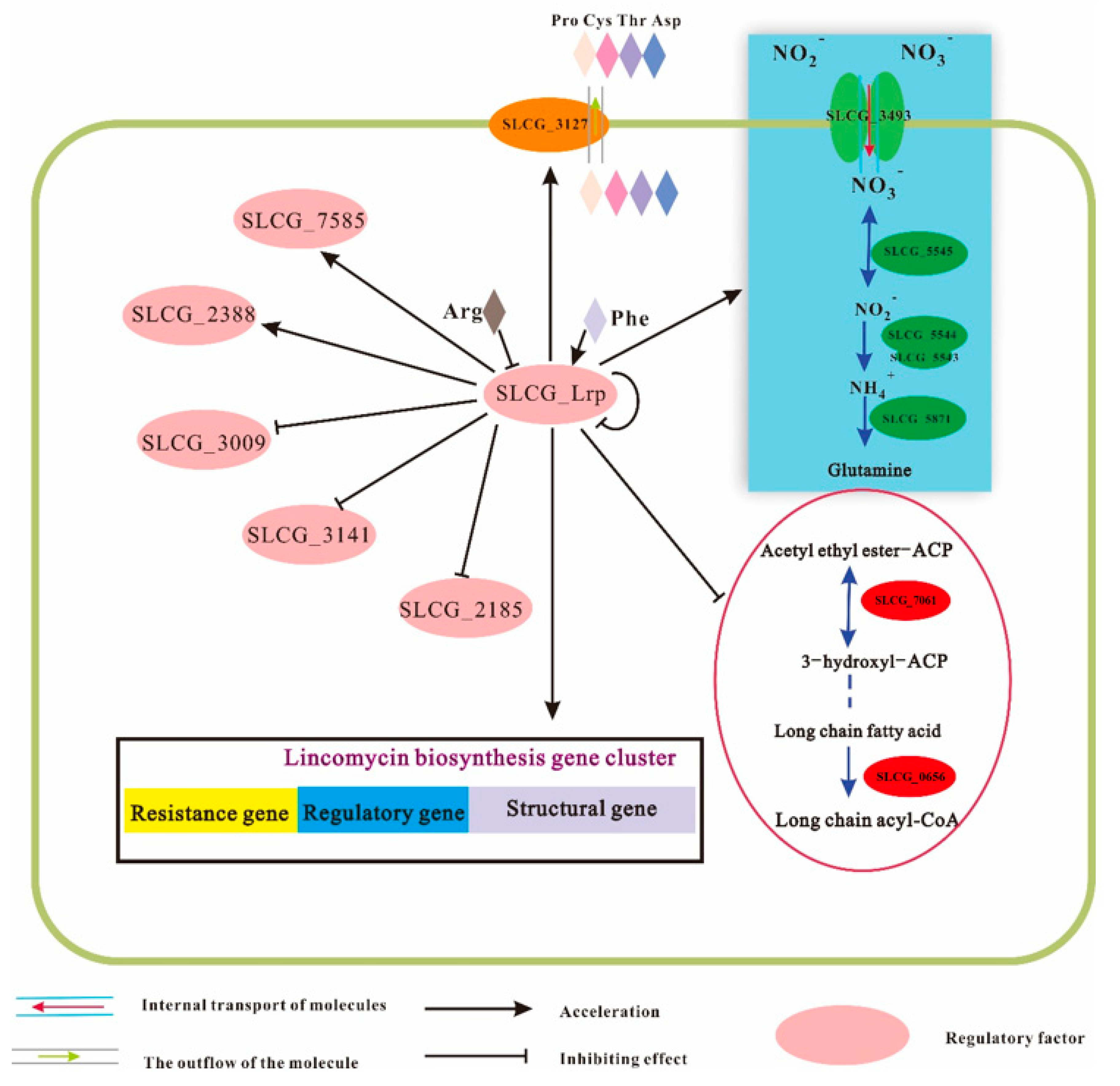
3.3. SLCG_Lrp Directly Promotes the Transcription of Nitrogen Metabolism Genes
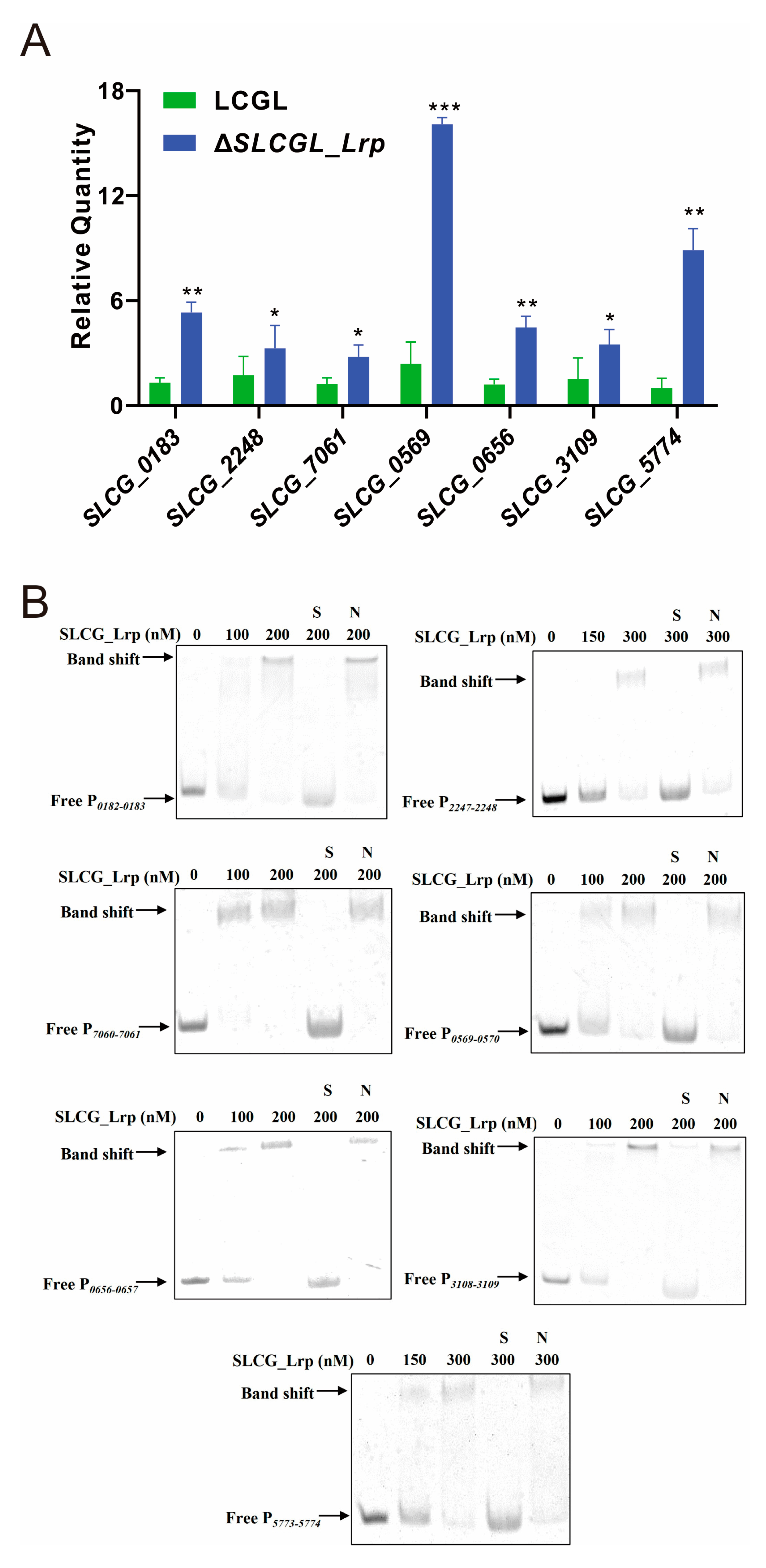
3.4. SLCG_Lrp Directly Represses the Transcription of Fatty Acid Metabolism Genes
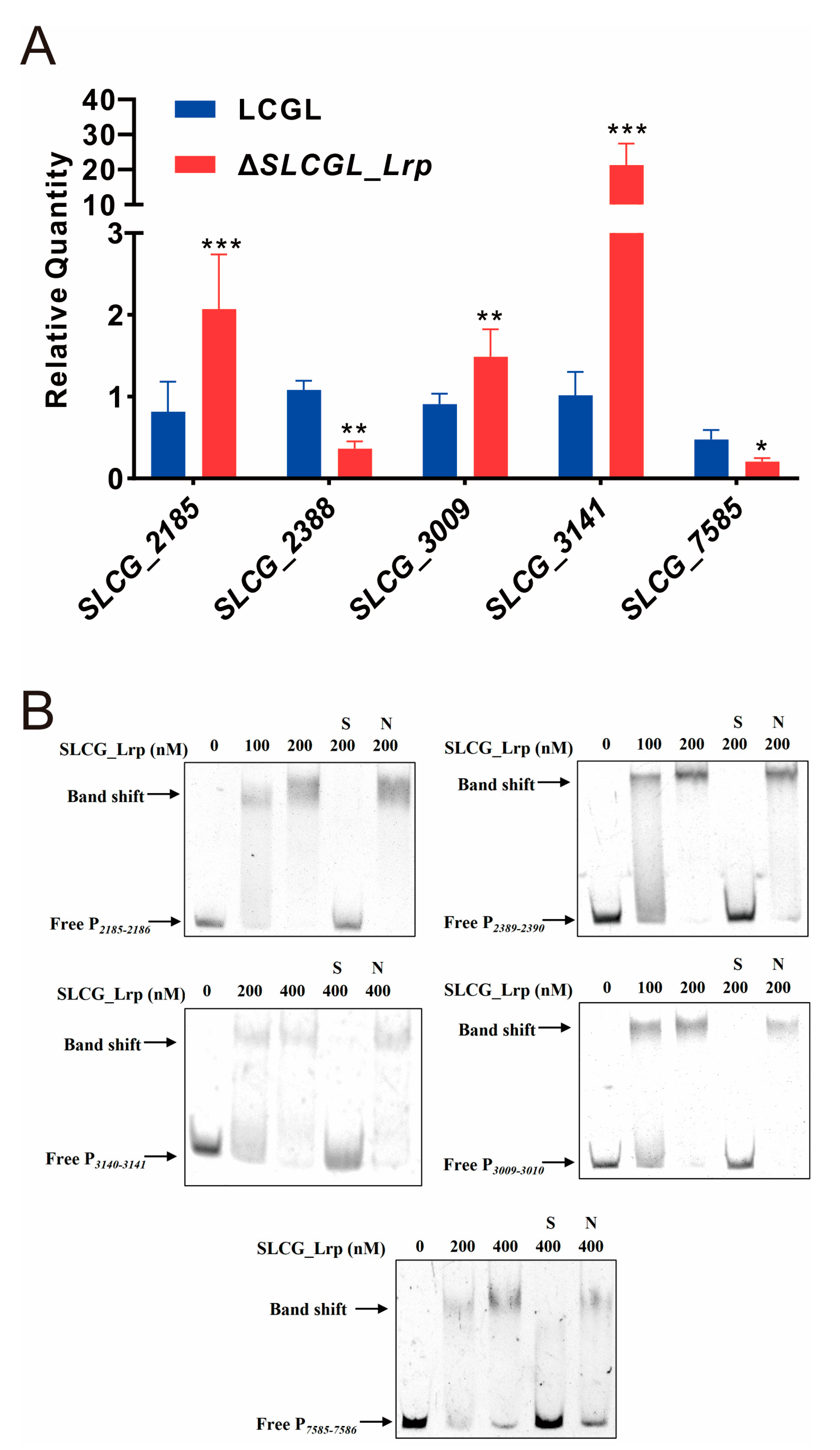
3.5. SLCG_Lrp Transcriptionally Modulates Five Regulatory Gene in S. lincolnensis
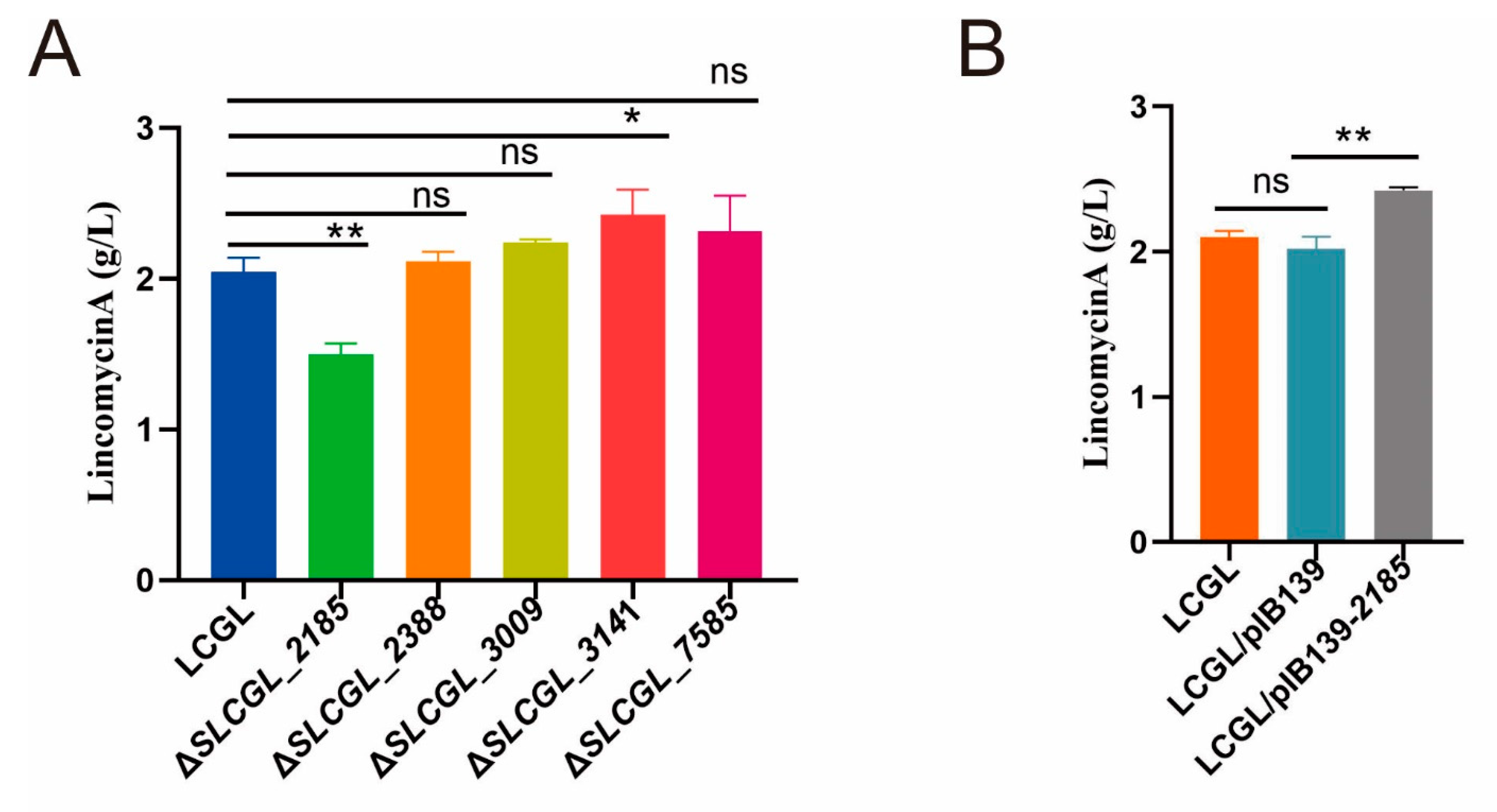
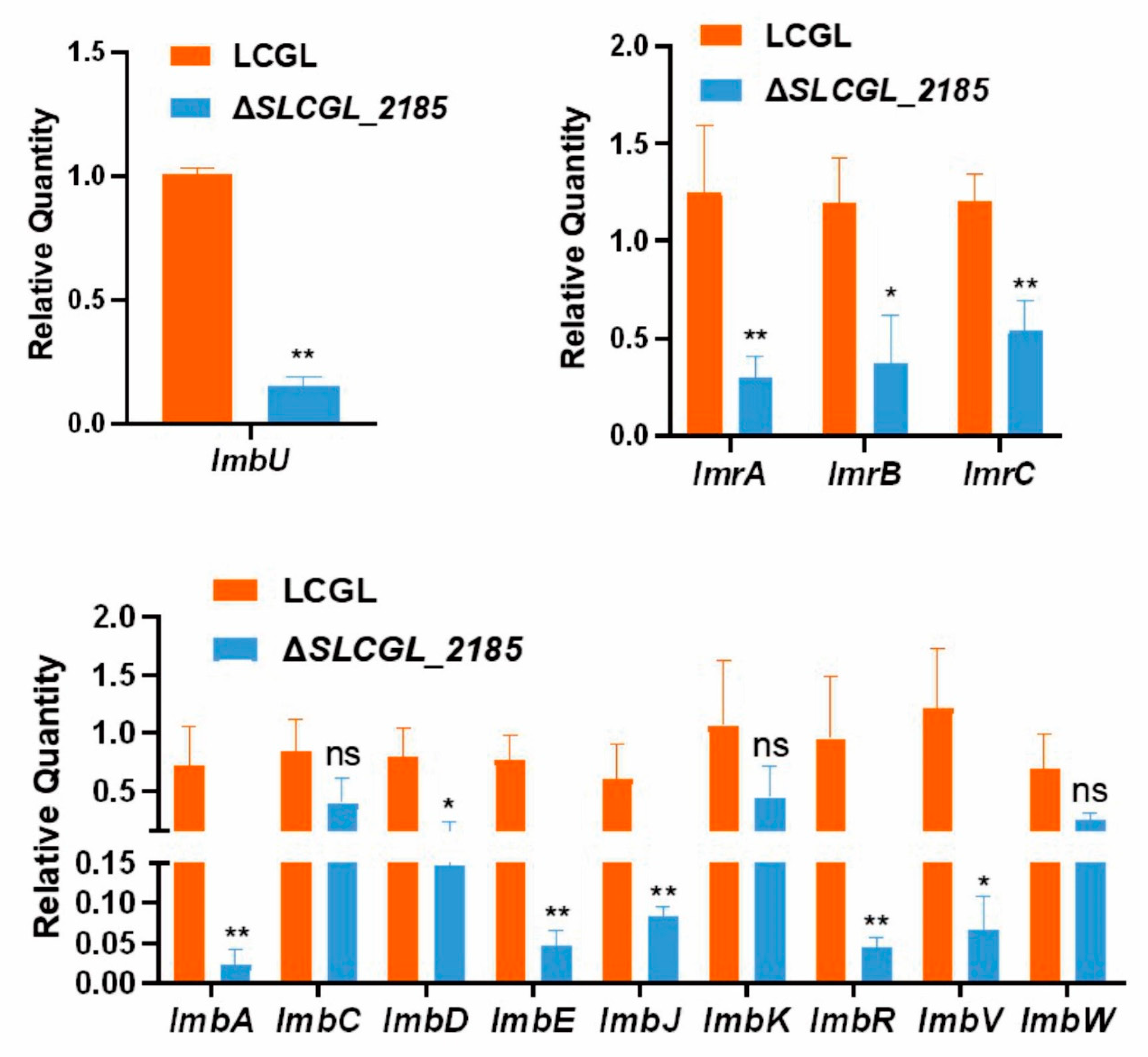
3.6. SLCG_2185 Shows Direct Transcriptional Activation on Lincomycin Biosynthetic Cluster
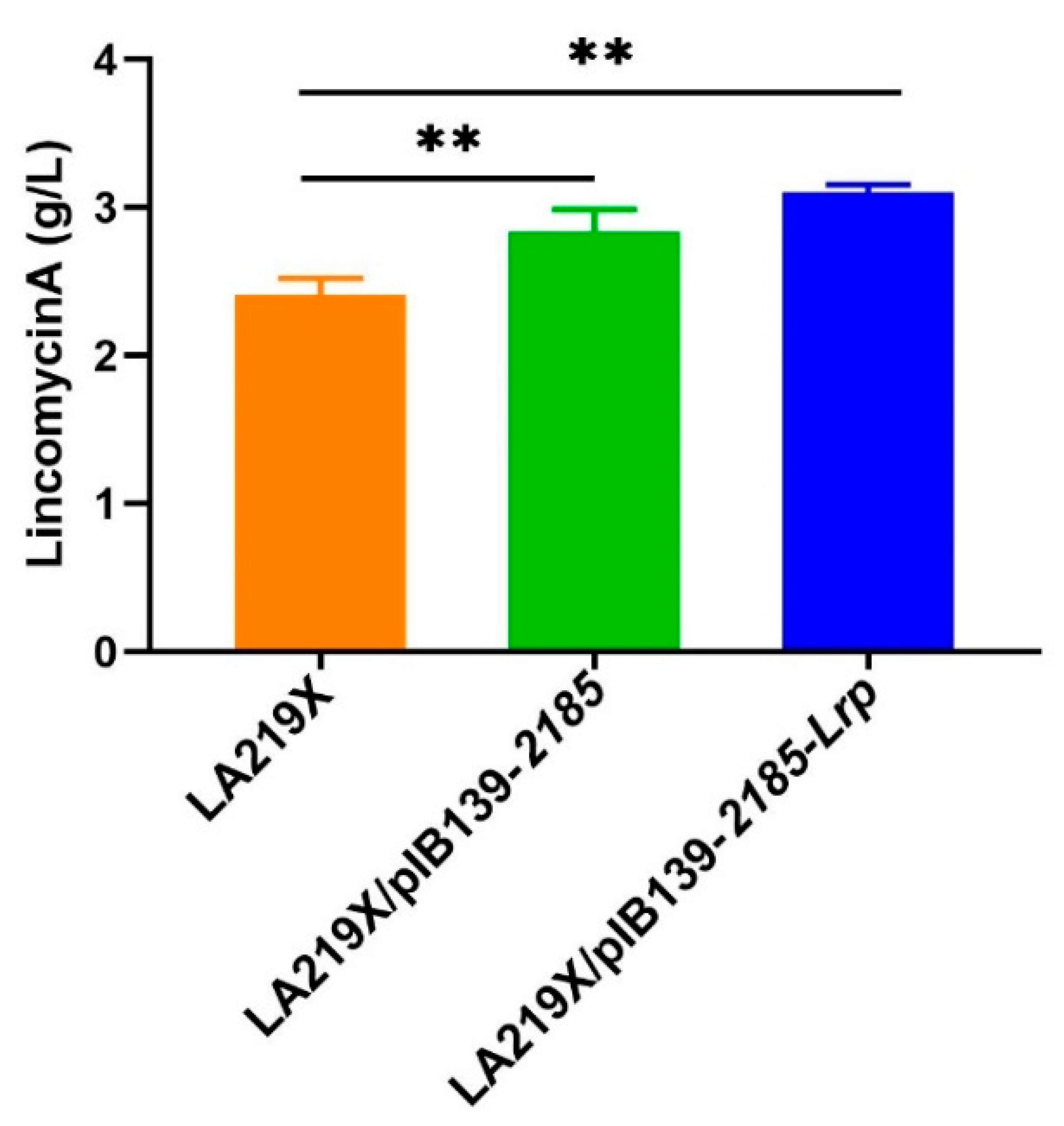
3.7. Rational Improvement of the Industrial High-Yield S. lincolnensis LA129X Strain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Spížek, J.; Řezanka, T. Lincomycin, cultivation of producing strains and biosynthesis. Appl. Microbiol. Biotechnol. 2003, 63, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, M.; Xu, D.; Zhang, Q.; Liu, W. Metabolic coupling of two small-molecule thiols programs the biosynthesis of lincomycin A. Nature 2015, 518, 115–119. [Google Scholar] [CrossRef]
- Wu, H.; Wang, Y.; Yuan, L.; Mao, Y.; Wang, W.; Zhu, L.; Wu, P.; Fu, C.; Müller, R.; Weaver, D.; et al. Inactivation of SACE_3446, a TetR family transcriptional regulator, stimulates erythromycin production in Saccharopolyspora erythraea. Synth Syst. Biotechnol. 2016, 1, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Koberská, M.; Kopecký, J.; Olsovská, J.; Jelínková, M.; Ulanova, D.; Man, P.; Flieger, M.; Janata, J. Sequence analysis and heterologous expression of the lincomycin biosynthetic cluster of the type strain Streptomyces lincolnensis ATCC 25466. Folia Microbiol. 2008, 53, 395–401. [Google Scholar] [CrossRef]
- Koberska, M.; Vesela, L.; Vimberg, V.; Lenart, J.; Vesela, J.; Kamenik, Z.; Janata, J.; Balikova Novotna, G. Beyond Self-Resistance: ABCF ATPase LmrC Is a Signal-Transducing Component of an Antibiotic-Driven Signaling Cascade Accelerating the Onset of Lincomycin Biosynthesis. mBio 2021, 12, e0173121. [Google Scholar] [CrossRef]
- Hou, B.; Zhu, X.; Kang, Y.; Wang, R.; Wu, H.; Ye, J.; Zhang, H. LmbU, a Cluster-Situated Regulator for Lincomycin, Consists of a DNA-Binding Domain, an Auto-Inhibitory Domain, and Forms Homodimer. Front. Microbiol. 2019, 10, 989. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, Q.; Zhang, Q.; Liu, W. Differences in PLP-Dependent Cysteinyl Processing Lead to Diverse S-Functionalization of Lincosamide Antibiotics. J. Am. Chem. Soc. 2016, 138, 6348–6351. [Google Scholar] [CrossRef] [PubMed]
- van der Heul, H.; Bilyk, B.; McDowall, K.; Seipke, R.; van Wezel, G. Regulation of antibiotic production in Actinobacteria: New perspectives from the post-genomic era. Nat. Prod. Rep. 2018, 35, 575–604. [Google Scholar] [CrossRef]
- Wang, R.; Kong, F.; Wu, H.; Hou, B.; Kang, Y.; Cao, Y.; Duan, S.; Ye, J.; Zhang, H. Complete genome sequence of high-yield strain S. lincolnensis B48 and identification of crucial mutations contributing to lincomycin overproduction. Synth Syst. Biotechnol. 2020, 5, 37–48. [Google Scholar] [CrossRef]
- Wu, H.; Chu, Z.; Zhang, W.; Zhang, C.; Ni, J.; Fang, H.; Chen, Y.; Wang, Y.; Zhang, L.; Zhang, B. Transcriptome-guided target identification of the TetR-like regulator SACE_5754 and engineered overproduction of erythromycin in Saccharopolyspora erythraea. J. Biol. Eng. 2019, 13, 11. [Google Scholar] [CrossRef]
- Ziegler, C.; Freddolino, P. The leucine-responsive regulatory proteins/feast-famine regulatory proteins: An ancient and complex class of transcriptional regulators in bacteria and archaea. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 373–400. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, H.; Xie, J. Regulatory and pathogenesis roles of Mycobacterium Lrp/AsnC family transcriptional factors. J. Cell Biochem. 2011, 112, 2655–2662. [Google Scholar] [CrossRef]
- Cho, B.; Barrett, C.; Knight, E.; Park, Y.; Palsson, B. Genome-scale reconstruction of the Lrp regulatory network in Escherichia coli. Proc. Natl. Acad. Sci. USA 2008, 105, 19462–19467. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Charlier, D. The Lrp family of transcription regulators in archaea. Archaea 2010, 2010, 750457. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, Y.; Wang, W.; Ren, M.; Wu, P.; Wang, Y.; Li, C.; Zhang, L.; Wu, H.; Weaver, D.; et al. Engineering of an Lrp family regulator SACE_Lrp improves erythromycin production in Saccharopolyspora erythraea. Metab. Eng. 2017, 39, 29–37. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Y.; Li, L.; Yang, E.; Wang, Y.; Wu, H.; Zhang, L.; Wang, W.; Zhang, B. Characterization and engineering of the Lrp/AsnC family regulator SACE_5717 for erythromycin overproduction in Saccharopolyspora erythraea. J. Ind. Microbiol. Biotechnol. 2019, 46, 1013–1024. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.; Dong, H.; Chen, Y.; Wang, Y.; Wu, H.; Li, C.; Weaver, D.; Zhang, L.; Zhang, B. Characterization of an Lrp/AsnC family regulator SCO3361, controlling actinorhodin production and morphological development in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2017, 101, 5773–5783. [Google Scholar] [CrossRef]
- Lu, Z.; Zhang, X.; Dai, J.; Wang, Y.; He, W. Engineering of leucine-responsive regulatory protein improves spiramycin and bitespiramycin biosynthesis. Microb. Cell Fact 2019, 18, 38. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; He, H.; Dong, S.; Tang, L.; Yang, E.; Wang, W.; Zhang, B. The leucine-responsive regulatory protein SCAB_Lrp modulates thaxtomin biosynthesis, pathogenicity, and morphological development in Streptomyces scabies. Mol. Plant Pathol. 2023, 24, 167–178. [Google Scholar] [CrossRef]
- Xu, Y.; Tang, Y.; Wang, N.; Liu, J.; Cai, X.; Cai, H.; Li, J.; Tan, G.; Liu, R.; Bai, L.; et al. Transcriptional regulation of a leucine-responsive regulatory protein for directly controlling lincomycin biosynthesis in Streptomyces lincolnensis. Appl. Microbiol. Biotechnol. 2020, 104, 2575–2587. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Du, L.; Liu, R.; Ying, L.; Zhao, G. An Efficient Intergeneric Conjugation of DNA from Escherichia coli to Mycelia of the Lincomycin-Producer Streptomyces lincolnensis. Int. J. Mol. Sci. 2012, 13, 4797–4806. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tan, G.; Ke, M.; Li, J.; Tang, Y.; Meng, S.; Niu, J.; Wang, Y.; Liu, R.; Wu, H.; et al. Enhanced lincomycin production by co-overexpression of metK1 and metK2 in Streptomyces lincolnensis. J. Ind. Microbiol. Biotechnol. 2018, 45, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Bierman, M.; Logan, R.; O’brien, K.; Seno, E.; Rao, R.N.; Schoner, B. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 1992, 116, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, M.; Mao, Y.; Li, W.; Liu, J.; Huang, X.; Zhou, Y.; Ye, B.; Zhang, L.; Weaver, D.; et al. Dissecting and engineering of the TetR family regulator SACE_7301 for enhanced erythromycin production in Saccharopolyspora erythraea. Microb. Cell Fact. 2014, 13, 158. [Google Scholar] [CrossRef]
- Xu, Y.; Ke, M.; Li, J.; Tang, Y.; Wang, N.; Tan, G.; Wang, Y.; Liu, R.; Bai, L.; Zhang, L.; et al. TetR-Type Regulator SLCG_2919 Is a Negative Regulator of Lincomycin Biosynthesis in Streptomyces lincolnensis. Appl. Environ. Microbiol. 2019, 85, e02091-18. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hellman, L.M.; Fried, M.G. Electrophoretic mobility shift assay (EMSA) for detecting protein–nucleic acid interactions. Nat. Protoc. 2007, 2, 1849–1861. [Google Scholar] [CrossRef] [PubMed]
- Lintner, R.; Mishra, P.; Srivastava, P.; Martinez-Vaz, B.; Khodursky, A.; Blumenthal, R. Limited functional conservation of a global regulator among related bacterial genera: Lrp in Escherichia, Proteus and Vibrio. BMC Microbiol. 2008, 8, 60. [Google Scholar] [CrossRef]
- Fukuda, M.; Takeda, H.; Kato, H.; Doki, S.; Ito, K.; Maturana, A.; Ishitani, R.; Nureki, O. Structural basis for dynamic mechanism of nitrate/nitrite antiport by NarK. Nat. Commun. 2015, 6, 7097. [Google Scholar] [CrossRef]
- Chu, S.; Zhang, D.; Wang, D.; Zhi, Y.; Zhou, P. Heterologous expression and biochemical characterization of assimilatory nitrate and nitrite reductase reveals adaption and potential of Bacillus megaterium NCT-2 in secondary salinization soil. Int. J. Biol. Macromol. 2017, 101, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Wu, H.; Wang, L.; Zhang, B.; Bai, L. Enhancement of antibiotic productions by engineered nitrate utilization in actinomycetes. Appl. Microbiol. Biotechnol. 2017, 101, 5341–5352. [Google Scholar] [CrossRef] [PubMed]
- Neusser, D.; Schmidt, H.; Spizèk, J.; Novotnà, J.; Peschke, U.; Kaschabeck, S.; Tichy, P.; Piepersberg, W. The genes lmbB1 and lmbB2 of Streptomyces lincolnensis encode enzymes involved in the conversion of L-tyrosine to propylproline during the biosynthesis of the antibiotic lincomycin A. Arch. Microbiol. 1998, 169, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Perli, T.; van der Vorm, D.; Wassink, M.; van den Broek, M.; Pronk, J.; Daran, J. Engineering heterologous molybdenum-cofactor-biosynthesis and nitrate-assimilation pathways enables nitrate utilization by Saccharomyces cerevisiae. Metab. Eng. 2021, 65, 11–29. [Google Scholar] [CrossRef]
- Fujihashi, M.; Nakatani, T.; Hirooka, K.; Matsuoka, H.; Fujita, Y.; Miki, K. Structural characterization of a ligand-bound form of Bacillus subtilis FadR involved in the regulation of fatty acid degradation. Proteins 2014, 82, 1301–1310. [Google Scholar] [CrossRef]
- Grau, F.; Burkovski, A.; Muller, Y. Crystal structures of adenylylated and unadenylylated P protein GlnK from Corynebacterium glutamicum. Acta Crystallogr. D Struct. Biol. 2021, 77, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Wang, L.; Li, S.; Pan, C.; Cheng, K.; Luo, Y.; Xu, H.; Tian, B.; Zhao, Y.; Hua, Y. Structure and DNA damage-dependent derepression mechanism for the XRE family member DG-DdrO. Nucleic Acids Res. 2019, 47, 9925–9933. [Google Scholar] [CrossRef]
- Zhu, Y.; Lu, T.; Zhang, J.; Zhang, P.; Tao, M.; Pang, X. A novel XRE family regulator that controls antibiotic production and development in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2020, 104, 10075–10089. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, M.; Zhou, H.; Li, C.; Luk, A.; Zhao, G.; Fung, K.; Ip, M. Role of Two-Component System Response Regulator bceR in the Antimicrobial Resistance, Virulence, Biofilm Formation, and Stress Response of Group B Streptococcus. Front. Microbiol. 2019, 10, 10. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Description | Reference |
|---|---|---|
| E. coli | ||
| DH5α | F recA lacZM15 | [21] |
| BL21 (DE3) | F− ompT hsdSB(rB− mB−) dcm gal λ(DE3) | Novagen |
| S. lincolnensis | ||
| LCGL | A lincomycin producer with artificially integrated attBΦC31 site | [23] |
| ΔSLCGL_Lrp | LCGL derivative with SLCG_Lrp deleted | [20] |
| ΔSLCGL_2185 | LCGL derivative with SLCG_2185 deleted | This study |
| ΔSLCGL_2388 ΔSLCGL_3009 ΔSLCGL_3141 ΔSLCGL_7585 | LCGL derivative with SLCG_2388 deleted LCGL derivative with SLCG_3009 deleted LCGL derivative with SLCG_3141 deleted LCGL derivative with SLCG_7585 deleted | This study This study This study This study |
| LCGL/pIB139 LCGL/pIB139-2185 | LCGL carrying pIB139 LCGL carrying pIB139-2185 | This study This study |
| LA219X LA219X/pIB139-2185 LA219X/pIB139-2185-Lrp | A lincomycin high-yield strain with artificial integrated attBφC31 site LA219X carrying pIB139-2185 LA219X carrying pIB139-2185-Lrp | [23] This study This study |
| Plasmids | ||
| pKC1139 | ori (pSG5), aac(3)IV, lacZ | [24] |
| pKC1139-Δ2185 | pKC1139 derivative containing two 1.8 kb fragments, the upstream and downstream regions of SLCG_2185 | This study |
| pKC1139-Δ2388 | pKC1139 derivative containing two 1.8 kb fragments, the upstream and downstream regions of SLCG_2388 | This study |
| pKC1139-Δ3009 | pKC1139 derivative containing two 1.8 kb fragments, the upstream and downstream regions of SLCG_3009 | This study |
| pKC1139-Δ3141 | pKC1139 derivative containing two 1.8 kb fragments, the upstream and downstream regions of SLCG_3141 | This study |
| pKC1139-Δ7585 | pKC1139 derivative containing two 1.8 kb fragments, the upstream and downstream regions of SLCG_7585 | This study |
| pIB139 pIB139-2185 pIB139-2185-Lrp | φC31 attP-int locus, acc(3)IV, oriT, PermE* pIB139 carrying an extra SLCG_2185 for gene overexpression pIB139 carrying an extra SLCG_2185 and SLCG_Lrp for gene overexpression | [20] This study This study |
| pET28a | T7 promoter, His-tag, kan | Novagen |
| pET28a-Lrp | pET28a-derived plasmid carrying SLCG_Lrp | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Xu, W.; Yi, J.; Li, B.; Liu, M.; Zhang, M.; Zheng, Y.; Liu, R.; Wu, H.; Zhang, B. Transcriptomics-Guided Investigation of the SLCG_Lrp Regulon Provides New Insights into Its Role for Lincomycin Biosynthesis. Fermentation 2023, 9, 396. https://doi.org/10.3390/fermentation9040396
Xu Y, Xu W, Yi J, Li B, Liu M, Zhang M, Zheng Y, Liu R, Wu H, Zhang B. Transcriptomics-Guided Investigation of the SLCG_Lrp Regulon Provides New Insights into Its Role for Lincomycin Biosynthesis. Fermentation. 2023; 9(4):396. https://doi.org/10.3390/fermentation9040396
Chicago/Turabian StyleXu, Yurong, Wanlian Xu, Jing Yi, Binglin Li, Meng Liu, Maifei Zhang, Yang Zheng, Ruihua Liu, Hang Wu, and Buchang Zhang. 2023. "Transcriptomics-Guided Investigation of the SLCG_Lrp Regulon Provides New Insights into Its Role for Lincomycin Biosynthesis" Fermentation 9, no. 4: 396. https://doi.org/10.3390/fermentation9040396