
Recent Advances in the Hydroxylation of Amino Acids and Its Derivatives
 , and
, and
Abstract
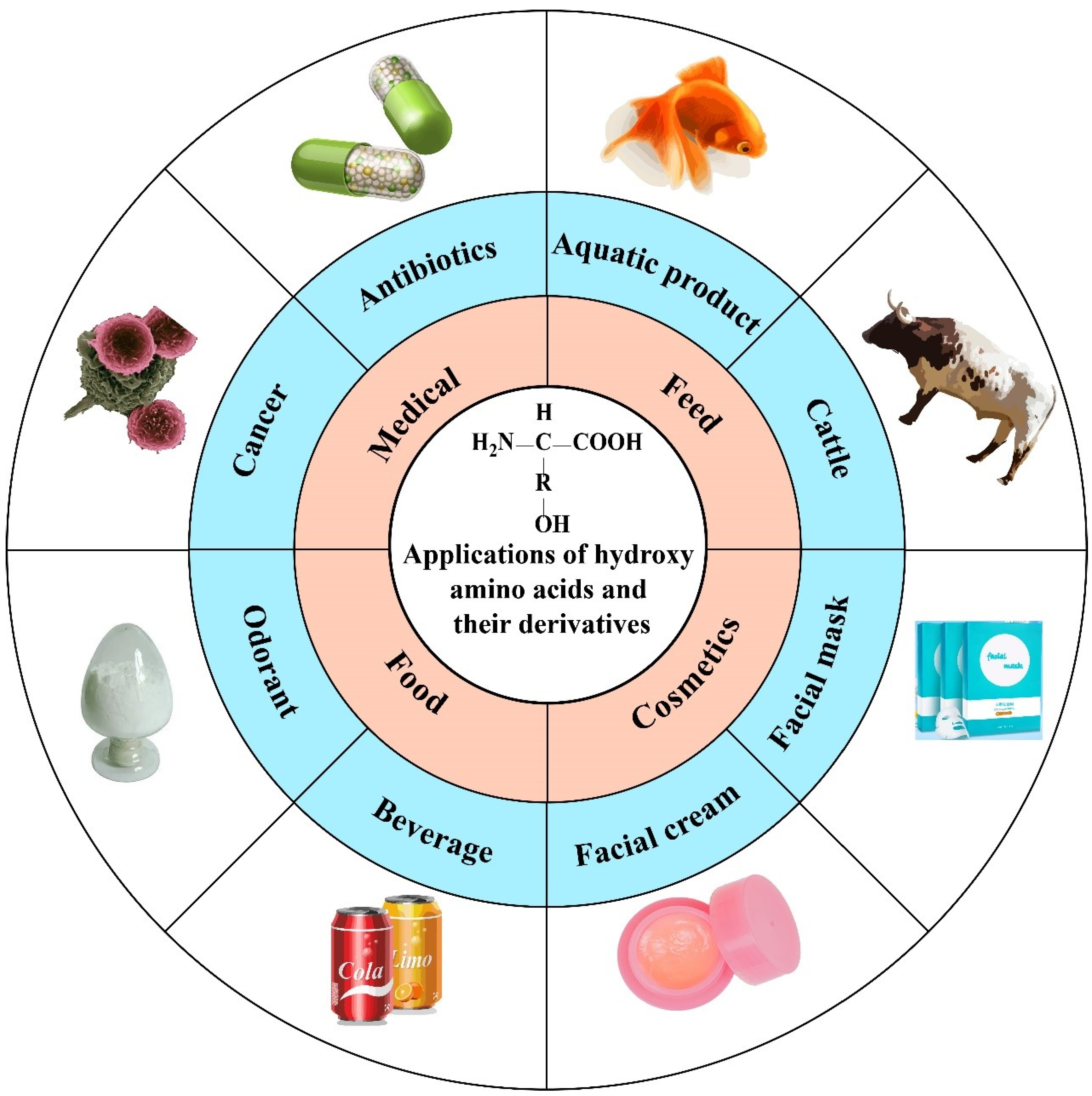
:1. Introduction
2. Hydroxylation of Aromatic Amino Acids
2.1. Hydroxylation of Tryptophan and Tryptophan Derivatives
2.2. Hydroxylation of Phenylalanine and Phenylalanine Derivatives
2.3. Hydroxylation of Tyrosine and Tyrosine Derivatives
3. Hydroxylation of Aliphatic Amino Acids and Their Derivatives
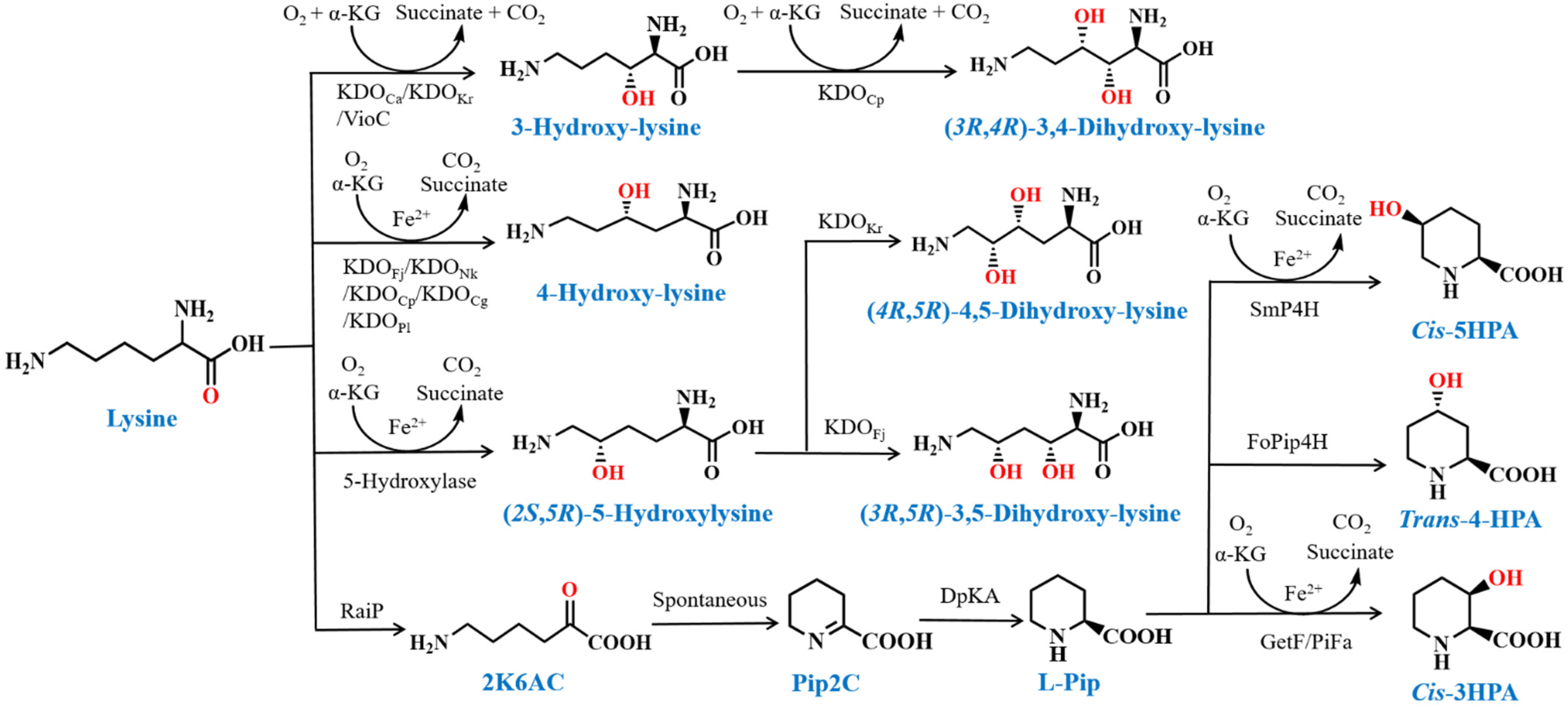
3.1. Hydroxylation of Lysine
3.2. Hydroxylation of Lysine Derivatives
3.3. Hydroxylation of Arginine
3.4. Hydroxylation of Aspartic Acid
3.5. Hydroxylation of Aspartic Acid Derivatives
3.6. Hydroxylation of Glutamic Acid
3.7. Hydroxylation of Leucine and Isoleucine
3.8. Hydroxylation of Leucine and Isoleucine Derivatives
4. Hydroxylation of Heterocyclic Amino Acids and Their Derivatives
Hydroxylation of Proline
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Wendisch, V.F. Metabolic engineering advances and prospects for amino acid production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef]
- Ren, X.; Fasan, R. Engineered and Artificial Metalloenzymes for Selective C-H Functionalization. Curr. Opin. Green Sustain. Chem. 2021, 31, 100494. [Google Scholar] [CrossRef] [PubMed]
- Münch, J.; Püllmann, P.; Zhang, W.; Weissenborn, M.J. Enzymatic Hydroxylations of sp3-Carbons. ACS Catal. 2021, 11, 9168–9203. [Google Scholar] [CrossRef]
- Hu, S.; Li, Y.; Zhang, A.; Li, H.; Chen, K.; Ouyang, P. Designing of an Efficient Whole-Cell Biocatalyst System for Converting L-Lysine into Cis-3-Hydroxypipecolic Acid. Front. Microbiol. 2022, 13, 945184. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zabriskie, T.M. VioC is a non-heme iron, alpha-ketoglutarate-dependent oxygenase that catalyzes the formation of 3S-hydroxy-L-arginine during viomycin biosynthesis. Chembiochem 2004, 5, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Sheng, H.; Li, Z.; Ye, Q. Biosynthesis of trans-4-hydroxyproline by recombinant strains of Corynebacterium glutamicum and Escherichia coli. BMC Biotechnol. 2014, 14, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, G.A.; Meisner, L.; Selby, K.; Stowe, R.; Carleton, B. Improved strength on 5-hydroxytryptophan and carbidopa in spinal cord atrophy. J. Neurol. Sci. 2017, 378, 59–62. [Google Scholar] [CrossRef]
- Remuzon, P. Trans-4-hydroxy-L-proline, a useful and versatile chiral starting block. Tetrahedron Lett. 1996, 52, 13803–13835. [Google Scholar] [CrossRef]
- Kurpejovic, E.; Wendisch, V.F.; Akbulut, B.S. Tyrosinase-based production of L-DOPA by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2021, 105, 9103–9111. [Google Scholar] [CrossRef]
- Xu, L.; Li, T.; Huo, Z.; Chen, Q.; Xia, Q.; Jiang, B. Directed Evolution Improves the Enzymatic Synthesis of L-5-Hydroxytryptophan by an Engineered Tryptophan Synthase. Appl. Biochem. Biotechnol. 2021, 193, 3407–3417. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, W.; Shi, F.; Huang, L.; Lian, J.; Qu, L.; Cai, J.; Xu, Z. Metabolic pathway engineering for high-level production of 5-hydroxytryptophan in Escherichia coli. Metab. Eng. 2018, 48, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; He, Y.; Su, N.; Bharath, S.R.; Tao, Y.; Jin, J.M.; Chen, W.; Song, H.; Tang, S.Y. Developing a highly efficient hydroxytyrosol whole-cell catalyst by de-bottlenecking rate-limiting steps. Nat. Commun. 2020, 11, 1515. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Chen, X.; Yang, C.; Chang, J.; Shen, W.; Fan, Y. Engineering Eschericha coli for Enhanced Tyrosol Production. J. Agric. Food Chem. 2017, 65, 4708–4714. [Google Scholar] [CrossRef] [PubMed]
- Rolf, J.; Nerke, P.; Britner, A.; Krick, S.; Lütz, S.; Rosenthal, K. From Cell-Free Protein Synthesis to Whole-Cell Biotransformation: Screening and Identification of Novel α-Ketoglutarate-Dependent Dioxygenases for Preparative-Scale Synthesis of Hydroxy-l-Lysine. Catalysts 2021, 11, 1038. [Google Scholar] [CrossRef]
- Hara, R.; Yamagata, K.; Miyake, R.; Kawabata, H.; Uehara, H.; Kino, K. Discovery of Lysine Hydroxylases in the Clavaminic Acid Synthase-like Superfamily for Efficient Hydroxylysine Bioproduction. Appl. Environ. Microbiol. 2017, 83, e00693-17. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Chen, J.; Ye, H.; Wu, H.; Sha, F.; Huang, F.; Cao, F.; Wei, P. Enzymatic hydroxylation of L-pipecolic acid by L-proline cis-4-hydroxylases and isomers separation. Biotechnol. Lett. 2020, 42, 2607–2617. [Google Scholar] [CrossRef]
- Mao, S.; Liu, X.; Gao, X.; Zhu, Z.; Sun, D.; Lu, F.; Qin, H.M. Design of an efficient whole-cell biocatalyst for the production of hydroxyarginine based on a multi-enzyme cascade. Bioresour. Technol. 2020, 318, 124261. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, S.V.; Kodera, T.; Samsonova, N.N.; Kotlyarova, V.A.; Rushkevich, N.Y.; Kivero, A.D.; Sokolov, P.M.; Hibi, M.; Ogawa, J.; Shimizu, S. Metabolic engineering of Escherichia coli to produce (2S, 3R, 4S)-4-hydroxyisoleucine. Appl. Microbiol. Biotechnol. 2010, 88, 719–726. [Google Scholar] [CrossRef]
- Sun, D.; Liu, X.; Zhu, M.; Chen, Y.; Li, C.; Cheng, X.; Zhu, Z.; Lu, F.; Qin, H.M. Efficient Biosynthesis of High-Value Succinic Acid and 5-Hydroxyleucine Using a Multienzyme Cascade and Whole-Cell Catalysis. J. Agric. Food Chem. 2019, 67, 12502–12510. [Google Scholar] [CrossRef]
- Johnston, R.M.; Chu, L.N.; Liu, M.; Goldberg, S.L.; Goswami, A.; Patel, R.N. Hydroxylation of l-proline to cis-3-hydroxy-l-proline by recombinant Escherichia coli expressing a synthetic l-proline-3-hydroxylase gene. Enzym. Microb. Technol. 2009, 45, 484–490. [Google Scholar] [CrossRef]
- Falcioni, F.; Buhler, B.; Schmid, A. Efficient hydroxyproline production from glucose in minimal media by Corynebacterium glutamicum. Biotechnol. Bioeng. 2015, 112, 322–330. [Google Scholar] [CrossRef]
- Prell, C.; Vonderbank, S.-A.; Meyer, F.; Pérez-García, F.; Wendisch, V.F. Metabolic engineering of Corynebacterium glutamicum for de novo production of 3-hydroxycadaverine. Curr. Res. Biotechnol. 2022, 4, 32–46. [Google Scholar] [CrossRef]
- Gendle, M.H.; Young, E.L.; Romano, A.C. Effects of oral 5-hydroxytryptophan on a standardized planning task: Insight into possible dopamine/serotonin interactions in the forebrain. Hum. Psychopharmacol. 2013, 28, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Jangid, P.; Malik, P.; Singh, P.; Sharma, M.; Gulia, A.K. Comparative study of efficacy of l-5-hydroxytryptophan and fluoxetine in patients presenting with first depressive episode. Asian J. Psychiatr. 2013, 6, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M.A.; Deive, F.J.; Alvarez, M.S.; Longo, M.A.; Rodriguez, A.; Bernal, C.; Martinez, R. Effect of process parameters and surfactant additives on the obtained activity of recombinant tryptophan hydroxylase (TPH1) for enzymatic synthesis of 5-hydroxytryptophan (5-HTP). Enzym. Microb. Technol. 2022, 154, 109975. [Google Scholar] [CrossRef] [PubMed]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F., 3rd; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, G.A.; Wagner, Z.; Markό, L.; Kszegi, T.; Mohás, M.; Kocsis, B.; Matus, Z.; Wagner, L.; Tamaskό, M.; Mazák, I.; et al. Urinary ortho-tyrosine excretion in diabetes mellitus and renal failure: Evidence for hydroxyl radical production. Kidney Int. 2005, 68, 2281–2287. [Google Scholar] [CrossRef] [Green Version]
- Molnár, G.A.; Nemes, V.; Biró, Z.; Ludány, A.; Wagner, Z.; Wittmann, I. Accumulation of the hydroxyl free radical markers meta-, ortho-tyrosine and DOPA in cataractous lenses is accompanied by a lower protein and phenylalanine content of the water-soluble phase. Free Radic. Res. 2005, 39, 1359–1366. [Google Scholar] [CrossRef]
- Zhang, W.; Ames, B.D.; Walsh, C.T. Identification of phenylalanine 3-hydroxylase for meta-tyrosine biosynthesis. Biochemistry 2011, 50, 5401–5403. [Google Scholar] [CrossRef] [Green Version]
- Seide, S.; Arnold, L.; Wetzels, S.; Bregu, M.; Gätgens, J.; Pohl, M. From Enzyme to Preparative Cascade Reactions with Immobilized Enzymes: Tuning Fe(II)/α-Ketoglutarate-Dependent Lysine Hydroxylases for Application in Biotransformations. Catalysts 2022, 12, 354. [Google Scholar] [CrossRef]
- Bastard, K.; Isabet, T.; Stura, E.A.; Legrand, P.; Zaparucha, A. Structural Studies based on two Lysine Dioxygenases with Distinct Regioselectivity Brings Insights into Enzyme Specificity within the Clavaminate Synthase-Like Family. Sci. Rep. 2018, 8, 16587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Zhu, M.; Song, Z.; Li, C.; Wang, Y.; Zhu, Z.; Sun, D.; Lu, F.; Qin, H.-M. Reshaping the Binding Pocket of Lysine Hydroxylase for Enhanced Activity. ACS Catal. 2020, 10, 13946–13956. [Google Scholar] [CrossRef]
- Amatuni, A.; Renata, H. Identification of a lysine 4-hydroxylase from the glidobactin biosynthesis and evaluation of its biocatalytic potential. Org. Biomol. Chem. 2019, 17, 1736–1739. [Google Scholar] [CrossRef]
- Baud, D.; Saaidi, P.-L.; Monfleur, A.; Harari, M.; Cuccaro, J.; Fossey, A.; Besnard, M.; Debard, A.; Mariage, A.; Pellouin, V.; et al. Synthesis of Mono- and Dihydroxylated Amino Acids with New α-Ketoglutarate-Dependent Dioxygenases: Biocatalytic Oxidation of C-H Bonds. ChemCatChem 2014, 6, 3012–3017. [Google Scholar] [CrossRef] [Green Version]
- Allevi, P.; Anastasia, M. A practical and simple synthesis of (2S,5R)- and (2S,5S)-5-hydroxylysine and of a related α-amino acid required for the synthesis of the collagen cross-link pyridinoline. Tetrahedron Asymmetry 2004, 15, 2091–2096. [Google Scholar] [CrossRef]
- Cheng, J.; Luo, Z.; Wang, B.; Yan, L.; Zhang, S.; Zhang, J.; Lu, Y.; Wang, W. An artificial pathway for trans-4-hydroxy-L-pipecolic acid production from L-lysine in Escherichia coli. Biosci. Biotechnol. Biochem. 2022, 86, 1476–1481. [Google Scholar] [CrossRef]
- Hibi, M.; Mori, R.; Miyake, R.; Kawabata, H.; Kozono, S.; Takahashi, S.; Ogawa, J. Novel Enzyme Family Found in Filamentous Fungi Catalyzing trans-4-Hydroxylation of L-Pipecolic Acid. Appl. Environ. Microbiol. 2016, 82, 2070–2077. [Google Scholar] [CrossRef] [Green Version]
- Helmetag, V.; Samel, S.A.; Thomas, M.G.; Marahiel, M.A.; Essen, L.O. Structural basis for the erythro-stereospecificity of the L-arginine oxygenase VioC in viomycin biosynthesis. FEBS J. 2009, 276, 3669–3682. [Google Scholar] [CrossRef] [Green Version]
- Ju, J.; Ozanick, S.G.; Shen, B.; Thomas, M.G. Conversion of (2S)-arginine to (2S,3R)-capreomycidine by VioC and VioD from the viomycin biosynthetic pathway of Streptomyces sp. strain ATCC11861. Chembiochem 2004, 5, 1281–1285. [Google Scholar] [CrossRef]
- Thomas, M.; Chan, Y.; Ozanick, S. Deciphering Tuberactinomycin Biosynthesis: Isolation, Sequencing, and Annotation of the Viomycin Biosynthetic Gene Cluster. Antimicrob. Agents Chemother. 2003, 47, 2823–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wang, B.; Bi, C.; He, G.; Chen, G. Efficient preparation of β-hydroxy aspartic acid and its derivatives. Chin. Chem. Lett. 2018, 29, 1113–1115. [Google Scholar] [CrossRef]
- Hara, R.; Nakano, M.; Kino, K. One-Pot Production of L-threo-3-Hydroxyaspartic Acid Using Asparaginase-Deficient Escherichia coli Expressing Asparagine Hydroxylase of Streptomyces coelicolor A3(2). Appl. Environ. Microbiol. 2015, 81, 3648–3654. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Chow, H.Y.; Po, K.H.L.; Ma, W.; Leung, E.L.Y.; Sun, Z.; Liu, M.; Chen, S.; Li, X. Total Synthesis and Structural Establishment/Revision of Antibiotics A54145. Org. Lett. 2019, 21, 5639–5644. [Google Scholar] [CrossRef]
- Jungmann, L.; Hoffmann, S.L.; Lang, C.; De Agazio, R.; Becker, J.; Kohlstedt, M.; Wittmann, C. High-efficiency production of 5-hydroxyectoine using metabolically engineered Corynebacterium glutamicum. Microb. Cell Factories 2022, 21, 274. [Google Scholar] [CrossRef]
- Strieker, M.; Nolan, E.M.; Walsh, C.T.; Marahiel, M.A. Stereospecific Synthesis of threo- and erythro-β-Hydroxyglutamic Acid During Kutzneride Biosynthesis. J. Am. Chem. Soc. 2009, 131, 13523–13530. [Google Scholar] [CrossRef] [Green Version]
- Ries, O.; Buschleb, M.; Granitzka, M.; Stalke, D.; Ducho, C. Amino acid motifs in natural products: Synthesis of O-acylated derivatives of (2S,3S)-3-hydroxyleucine. Beilstein J. Org. Chem. 2014, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Smirnov, S.V.; Sokolov, P.M.; Kodera, T.; Sugiyama, M.; Hibi, M.; Shimizu, S.; Yokozeki, K.; Ogawa, J. A novel family of bacterial dioxygenases that catalyse the hydroxylation of free L-amino acids. FEMS Microbiol. Lett. 2012, 331, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Hibi, M.; Kasahara, T.; Kawashima, T.; Yajima, H.; Kozono, S.; Smirnov, S.V.; Kodera, T.; Sugiyama, M.; Shimizu, S.; Yokozeki, K.; et al. Multi-Enzymatic Synthesis of Optically Pure β-Hydroxy α-Amino Acids. Adv. Synth. Catal. 2015, 357, 767–774. [Google Scholar] [CrossRef]
- Hansen, D.B.; Starr, M.-L.; Tolstoy, N.; Joullié, M.M. A stereoselective synthesis of (2S,4R)-δ-hydroxyleucine methyl ester: A component of cyclomarin A. Tetrahedron Asymmetry 2005, 16, 3623–3627. [Google Scholar] [CrossRef]
- Wu, L.; An, J.; Jing, X.; Chen, C.-C.; Dai, L.; Xu, Y.; Liu, W.; Guo, R.-T.; Nie, Y. Molecular Insights into the Regioselectivity of the Fe(II)/2-Ketoglutarate-Dependent Dioxygenase-Catalyzed C–H Hydroxylation of Amino Acids. ACS Catal. 2022, 12, 11586–11596. [Google Scholar] [CrossRef]
- Zwick, C.R., 3rd; Renata, H. Remote C-H Hydroxylation by an alpha-Ketoglutarate-Dependent Dioxygenase Enables Efficient Chemoenzymatic Synthesis of Manzacidin C and Proline Analogs. J. Am. Chem. Soc. 2018, 140, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, H.; Zhang, T.; Pijning, T.; Yu, L.; Zhang, W.; Liu, W.; Meng, X. Biochemical and genetic characterization of fungal proline hydroxylase in echinocandin biosynthesis. Appl. Microbiol. Biotechnol. 2018, 102, 7877–7890. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Hüttel, W. A Simple Procedure for Selective Hydroxylation of L-Proline and L-Pipecolic Acid with Recombinantly Expressed Proline Hydroxylases. Adv. Synth. Catal. 2011, 353, 1375–1383. [Google Scholar] [CrossRef]
- Davies, S.G.; Fletcher, A.M.; Linsdall, S.M.; Roberts, P.M.; Thomson, J.E. Asymmetric Syntheses of (2 R,3 S)-3-Hydroxyproline and (2 S,3 S)-3-Hydroxyproline. Org. Lett. 2018, 20, 4135–4139. [Google Scholar] [CrossRef] [PubMed]
- Theodosiou, E.; Frick, O.; Buhler, B.; Schmid, A. Metabolic network capacity of Escherichia coli for Krebs cycle-dependent proline hydroxylation. Microb. Cell Factories 2015, 14, 108. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Fernández, M.; Avenoza, A.; Busto, J.H.; Peregrina, J.M.; Rodríguez, F. Cyclobutane serine amino acid derivatives as 5-hydroxyproline precursors. ARKIVOC 2009, 2010, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Xia, S.; Lin, H.; Liu, J.; Huang, W. Microbial Proline Racemase-Proline Dehydrogenase Cascade for Efficient Production of D-proline and N-boc-5-hydroxy-L-proline from L-proline. Appl. Biochem. Biotechnol. 2022, 194, 4135–4146. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Product | Chemical Structure | Host | Substrate | Titer (g/L) | Strategy | Reference |
|---|---|---|---|---|---|---|
| 3,4-Dihydroxyphenyl-L-alanine |  | Corynebacterium glutamicum | L-Tyrosine | 0.26 | Heterologous expression of tyrosinase from Ralstonia solanacearum | [10] |
| 5-HTP |  | E. coli | L-Serine | 17.79 | Directed evolution of tryptophan synthase | [11] |
| 5-HTP |  | E. coli | L-Tryptophan | 5.1 | Expression of TPH1 | [12] |
| Hydroxytyrosol |  | E. coli | Glucose | 4.69 | Directed Optimization of VanR and TYO | [13] |
| Tyrosol |  | E. coli | Tyrosine | 1.18 | Introducing a yeast pyruvate decarboxylase ARO10 into E. coli | [14] |
| (3S)-3-Hydroxy-L-lysine |  | E. coli | L-Lysine | 5 | Screening and identifying novel KDOs | [15] |
| (2S,3S)-3-Hydroxylysine |  | E. coli | L-Lysine | 86.1 | Gene mining and phylogenetic analysis from Clavaminic acid synthase-like superfamily | [16] |
| (2S,4R)-4-Hydroxylysine |  | E. coli | L-Lysine | 43.0 | Gene mining and phylogenetic analysis from Clavaminic acid synthase-like superfamily | [16] |
| cis-3HPA |  | E. coli | L-Lysine | 47.9 | Expression of GetF and SpLCD | [5] |
| cis-5HPA |  | E. coli | L-Lysine | 13.95 | Established a complete biological method | [17] |
| 3-OH-Arg |  | E. coli | L-Arginine | 9.9 | Using protein engineering and recombinant whole-cell biocatalysis | [18] |
| 4-HIL |  | E. coli | L-Isoleucine | 10.83 | Cloning of the IDO gene from Bacillus thuringiensis and B. thuringiensis strains | [19] |
| 5-HLeu |  | E. coli | L-Leucine | 18.83 | Constructing two bioconversion systems | [20] |
| c3Hyp |  | E. coli | L-Proline | 9.1 | Cloning and heterologous expression of P3H | [21] |
| Trans-4-hydroxy-L-proline |  | Corynebacterium glutamicum | Glucose | 7.1 | Constructed a plasmid (pEKE_p4h1of) containing the P4H gene | [22] |
| Trans-4-hydroxy-L-proline |  | Corynebacterium glutamicum | L-proline | 0.113 | Cloned and expressed the genes of trans-proline 4-hydroxylase from diverse resources | [7] |
| 5-Hydroxylysine |  | Corynebacterium glutamicum | Lysine | 0.88 | Overexpressed KDOs genes | [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Xiao, S.; Zhao, X.; Zhao, L.; Zhang, Y.; Cheng, J.; Zhang, J. Recent Advances in the Hydroxylation of Amino Acids and Its Derivatives. Fermentation 2023, 9, 285. https://doi.org/10.3390/fermentation9030285
Wang B, Xiao S, Zhao X, Zhao L, Zhang Y, Cheng J, Zhang J. Recent Advances in the Hydroxylation of Amino Acids and Its Derivatives. Fermentation. 2023; 9(3):285. https://doi.org/10.3390/fermentation9030285
Chicago/Turabian StyleWang, Bangxu, Shujian Xiao, Xingtao Zhao, Liming Zhao, Yin Zhang, Jie Cheng, and Jiamin Zhang. 2023. "Recent Advances in the Hydroxylation of Amino Acids and Its Derivatives" Fermentation 9, no. 3: 285. https://doi.org/10.3390/fermentation9030285