

Fermented (By Monascus purpureus or Aspergillus oryzae) and Non-Fermented Defatted Soybean Flour Extracts: Biological Insight and Mechanism Differences in Inflammatory Pain and Peritonitis
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Defatted Soybean Fermentation by M. purpureus and A. ooryzae (DSFF-Mp/DSFF-Ao)
2.2. Determination of Total Phenolic Content and Isoflavones, and Soluble Sugar Composition of Extracts
2.3. Chemicals
2.4. Animals
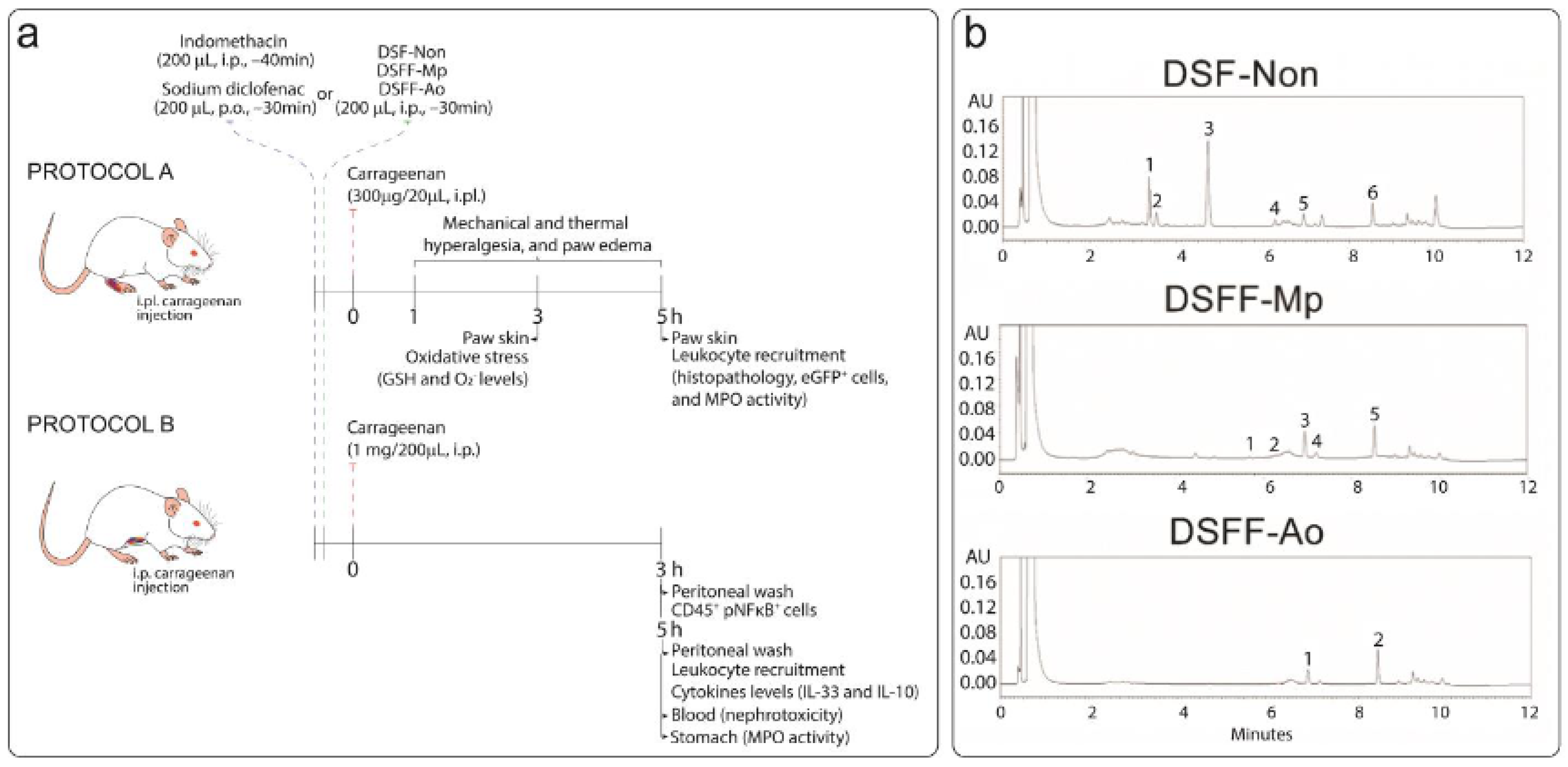
2.5. In Vivo Experimental Approaches
2.6. Mechanical Hyperalgesia Evaluation
2.7. Thermal Hyperalgesia Test
2.8. Paw Edema Assessment
2.9. Histopathology
2.10. Paw Tissue Immunofluorescence
2.11. MPO Activity Assay
2.12. Evaluation of GSH and O2− Levels
2.13. Leukocyte Recruitment into Peritoneal Cavity
2.14. Cytokine Quantitation Procedure
2.15. Immunofluorescence Staining in Samples of Peritoneal Cavity Cells
2.16. Renal Function Tests
2.17. Statistical Analysis
3. Results
3.1. Characterization of DSF Extracts
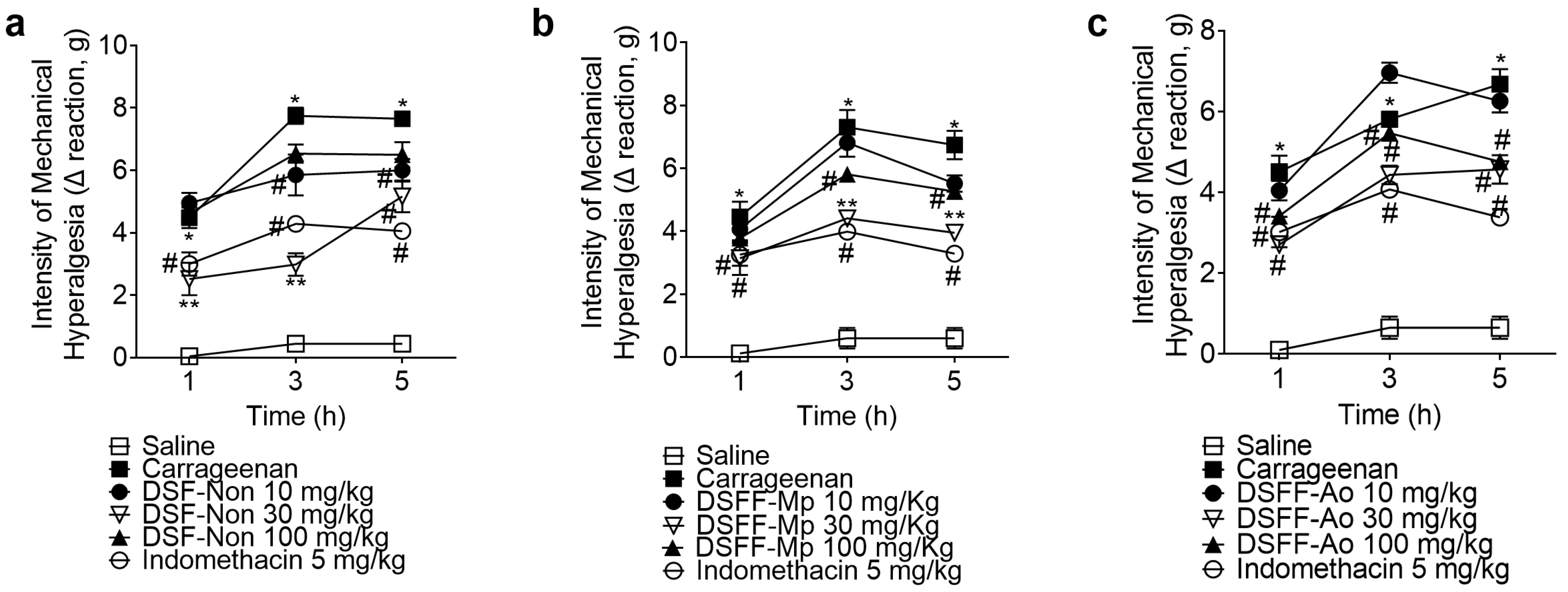
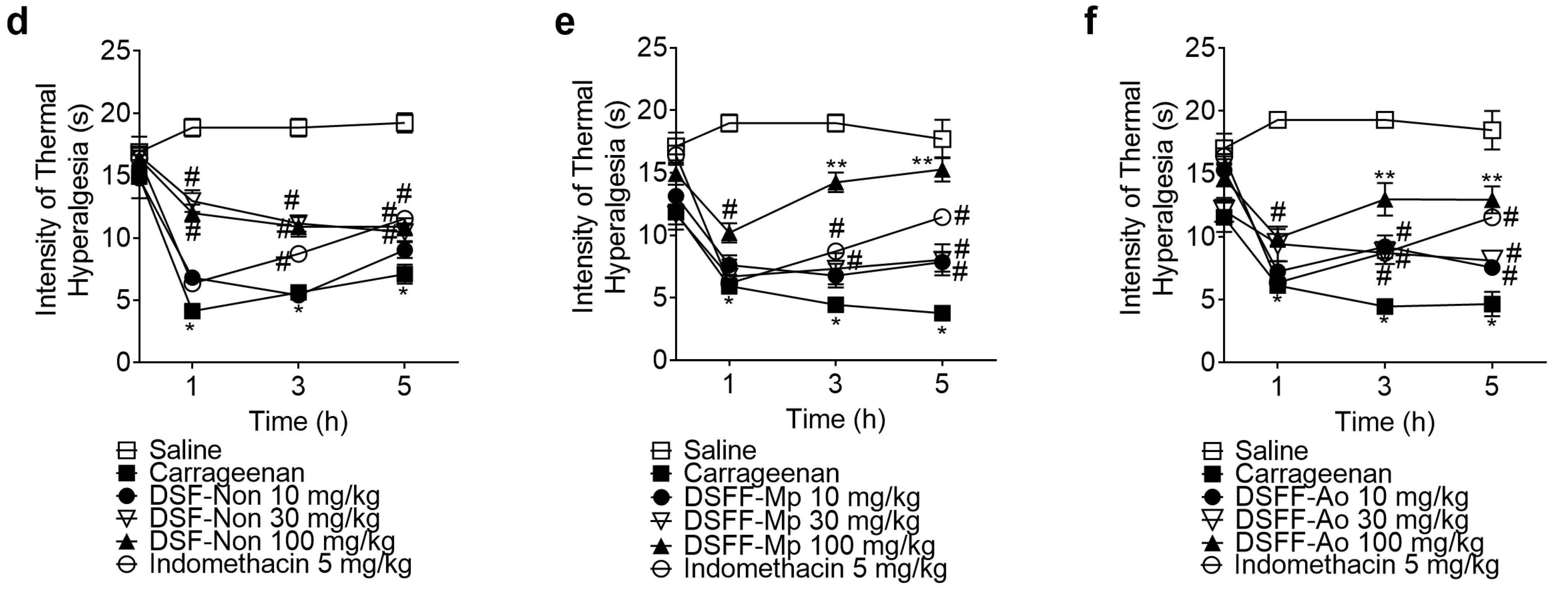
3.2. DSF-Non, DSFF-Mp, and DSFF-Ao Inhibited Carrageenan-Induced Mechanical Hyperalgesia and Thermal Hyperalgesia
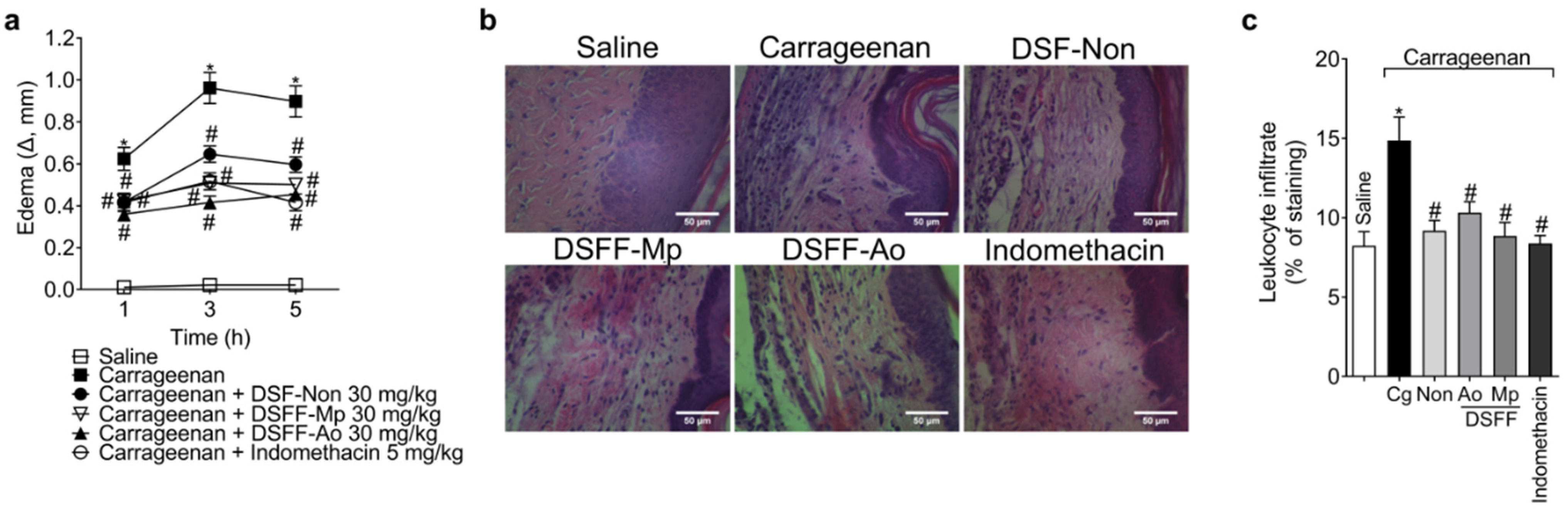
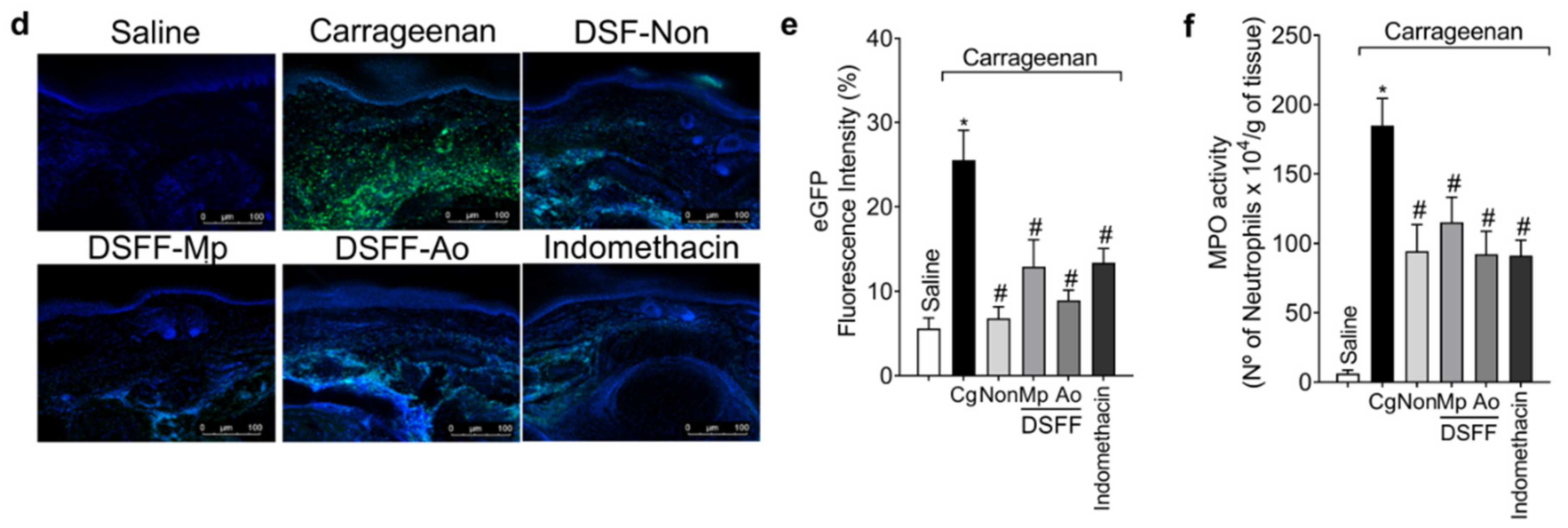
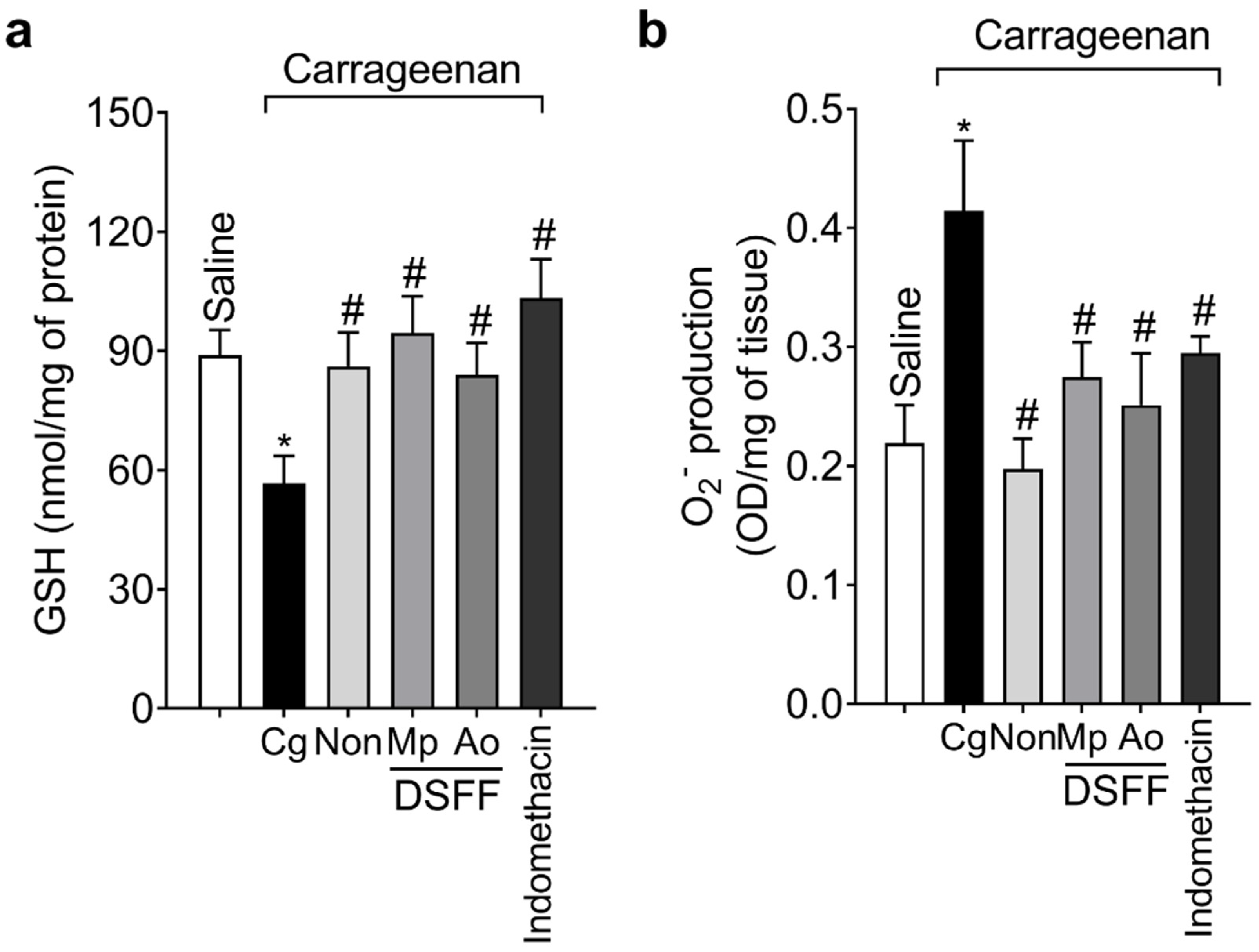
3.3. DSF-Non, DSFF-Mp, and DSFF-Ao Inhibited Carrageenan-Induced Edema, Leukocyte Recruitment, and Oxidative Stress in the Paw Tissue
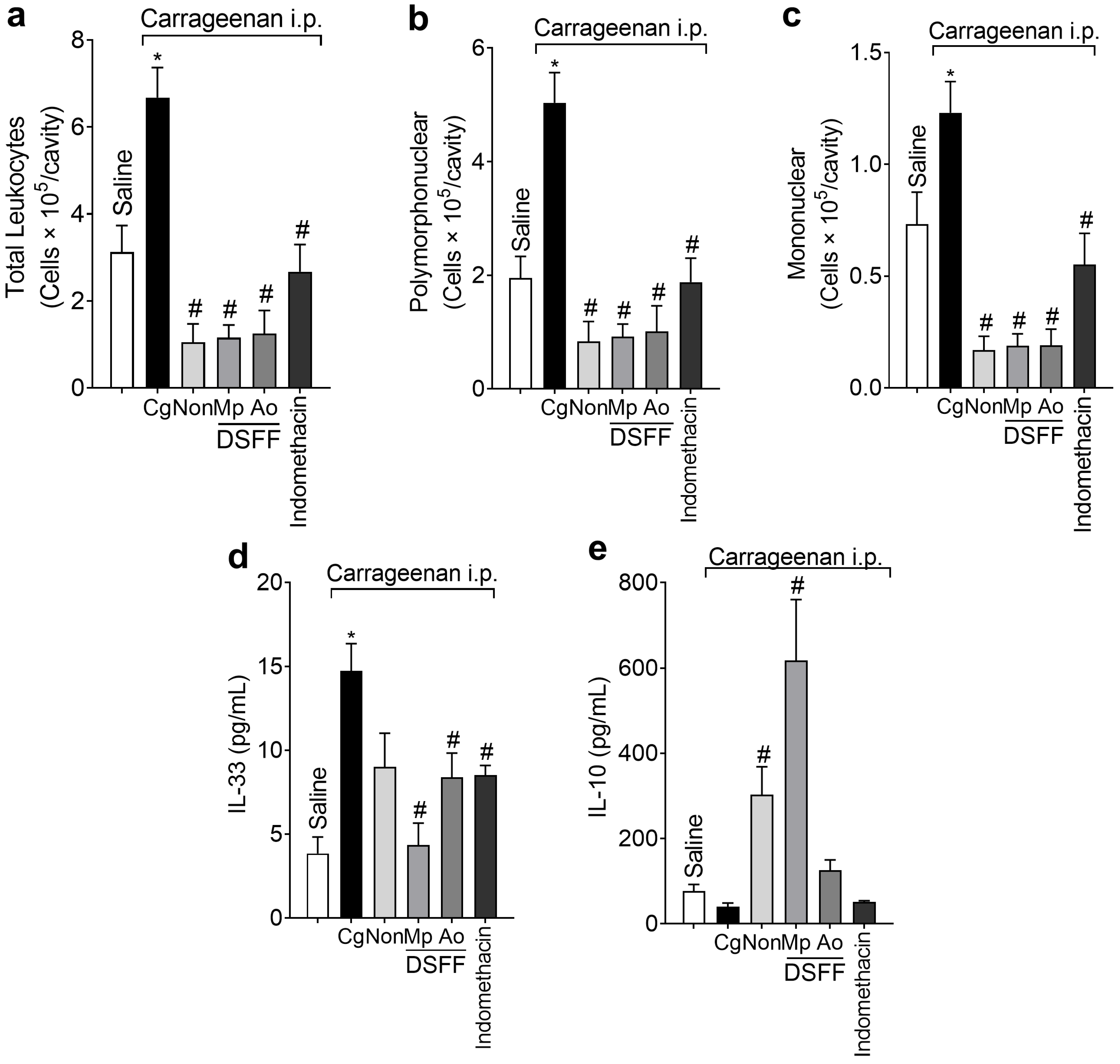
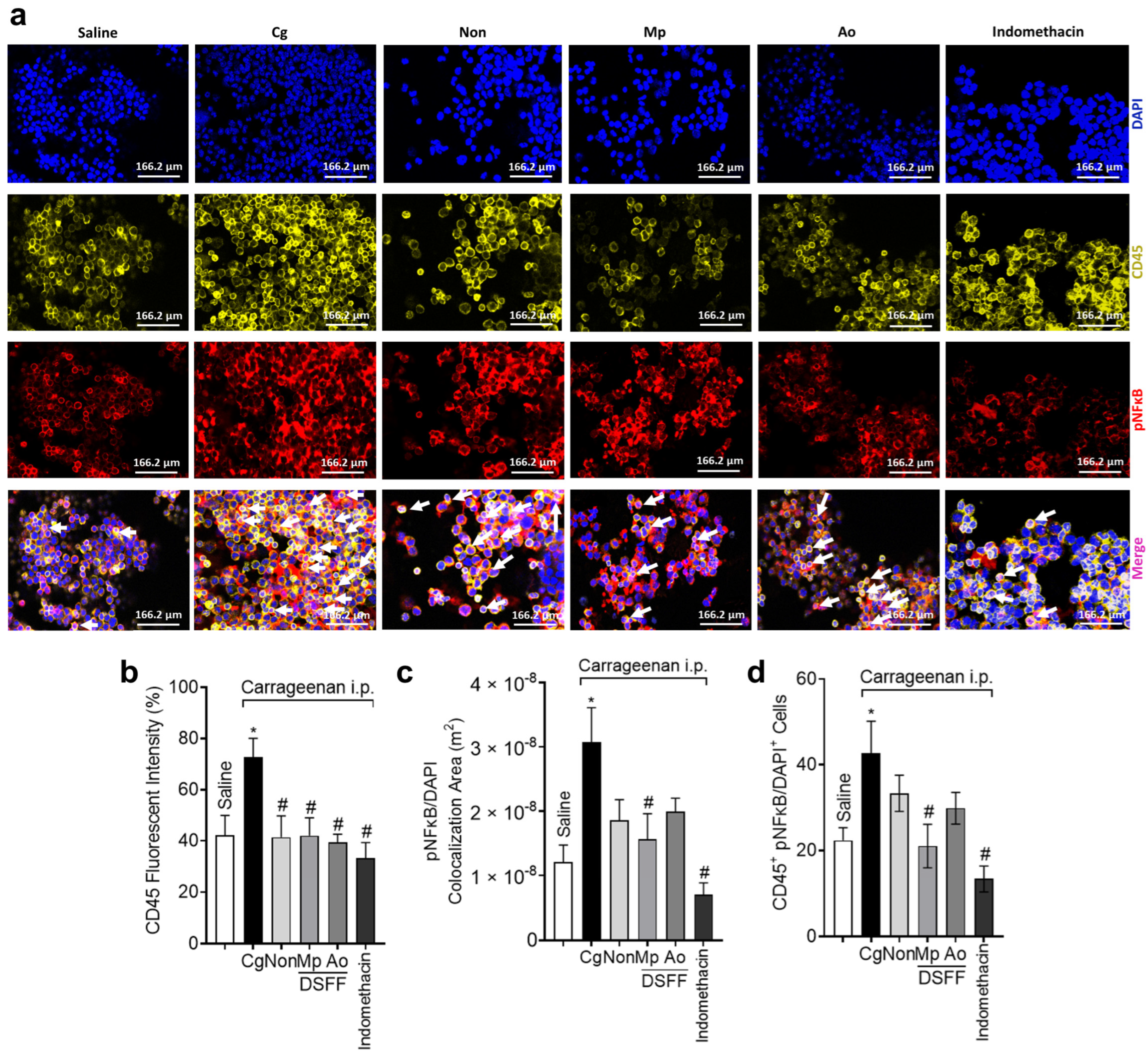
3.4. DSF-Non, DSFF-Mp, and DSFF-Ao Inhibited Leukocyte Recruitment and Differentially Modulated NFκB Activation in Peritoneal Leukocytes Recruited by Carrageenan
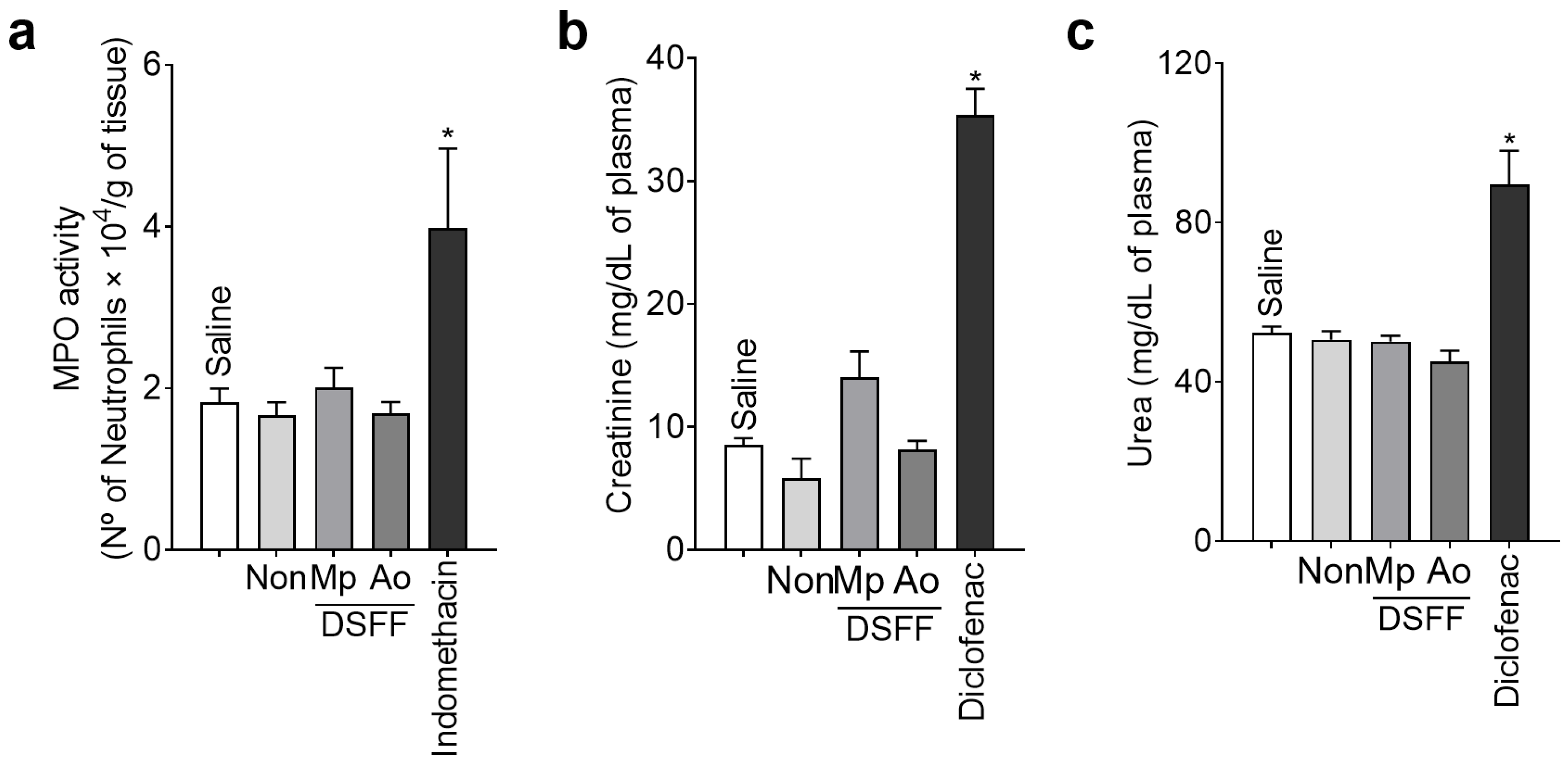
3.5. DSF-Non, DSFF-Mp, and DSFF-Ao Extracts Did Not Induce Stomach or Kidney Toxicity
4. Discussion
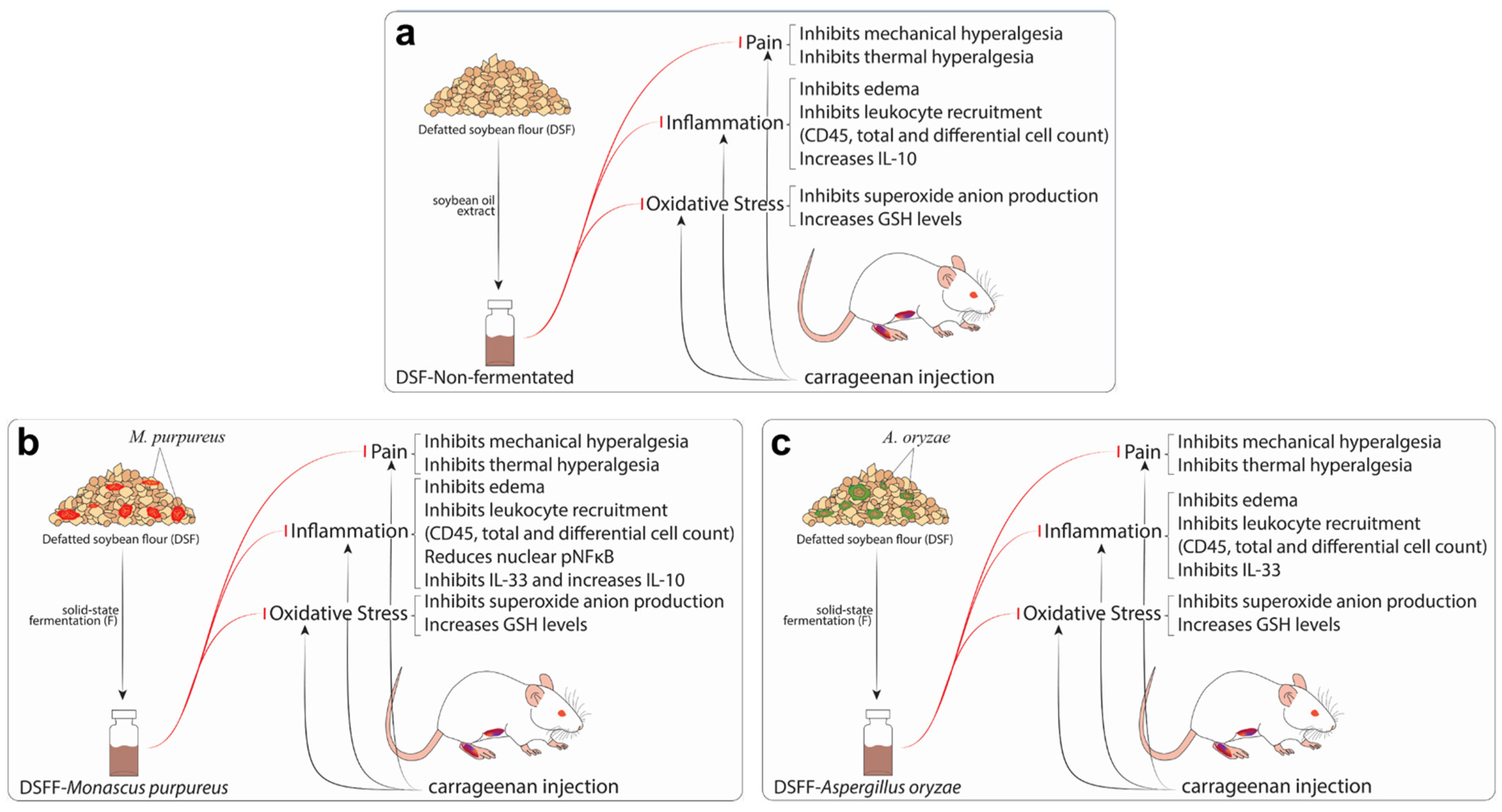
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Sugano, M. (Ed.) Soy in Health and Disease Prevention, 1st ed.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Villalobos Mdel, C.; Serradilla, M.J.; Martin, A.; Ordiales, E.; Ruiz-Moyano, S.; Cordoba Mde, G. Antioxidant and antimicrobial activity of natural phenolic extract from defatted soybean flour by-product for stone fruit postharvest application. J. Sci. Food Agric. 2016, 96, 2116–2124. [Google Scholar] [CrossRef] [PubMed]
- Genovese, M.I.; Barbosa, A.C.; Pinto Mda, S.; Lajolo, F.M. Commercial soy protein ingredients as isoflavone sources for functional foods. Plant Foods Hum. Nutr. 2007, 62, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Izumi, T.; Piskula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Faraj, A.; Vasanthan, T. Soybean Isoflavones: Effects of Processing and Health Benefits. Food Rev. Int. 2004, 20, 51–75. [Google Scholar] [CrossRef]
- Handa, C.L.; Couto, U.R.; Vicensoti, A.H.; Georgetti, S.R.; Ida, E.I. Optimisation of soy flour fermentation parameters to produce beta-glucosidase for bioconversion into aglycones. Food Chem. 2014, 152, 56–65. [Google Scholar] [CrossRef]
- Pyo, Y.H.; Seong, K.S. Hypolipidemic effects of Monascus-fermented soybean extracts in rats fed a high-fat and -cholesterol diet. J. Agric. Food Chem. 2009, 57, 8617–8622. [Google Scholar] [CrossRef]
- Chiang, S.S.; Chang, S.P.; Pan, T.M. Osteoprotective effect of Monascus-fermented dioscorea in ovariectomized rat model of postmenopausal osteoporosis. J. Agric. Food Chem. 2011, 59, 9150–9157. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Chen, J.; Li, X.-L.; Yi, K.; Ye, Y.; Liu, G.; Wang, S.-F.; Hu, H.-L.; Zou, L.; Wang, Z.-G. Dynamic changes in antioxidant activity and biochemical composition of tartary buckwheat leaves during Aspergillus niger fermentation. J. Funct. Foods 2017, 32, 375–381. [Google Scholar] [CrossRef]
- Da Silva, L.; Celeghini, R.; Chang, Y. Effect of the fermentation of whole soybean flour on the conversion of isoflavones from glycosides to aglycones. Food Chem. 2011, 128, 640–644. [Google Scholar] [CrossRef]
- Handa, C.L.; de Lima, F.S.; Guelfi, M.F.; Georgetti, S.R.; Ida, E.I. Multi-response optimisation of the extraction solvent system for phenolics and antioxidant activities from fermented soy flour using a simplex-centroid design. Food Chem. 2016, 197, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Peirs, C.; Seal, R.P. Targeting Toll-like receptors to treat chronic pain. Nat. Med. 2015, 21, 1251–1252. [Google Scholar] [CrossRef] [PubMed]
- Verri, W.A., Jr.; Cunha, T.M.; Parada, C.A.; Poole, S.; Cunha, F.Q.; Ferreira, S.H. Hypernociceptive role of cytokines and chemokines: Targets for analgesic drug development? Pharmacol. Ther. 2006, 112, 116–138. [Google Scholar] [CrossRef]
- Maioli, N.A.; Zarpelon, A.C.; Mizokami, S.S.; Calixto-Campos, C.; Guazelli, C.F.; Hohmann, M.S.; Pinho-Ribeiro, F.A.; Carvalho, T.T.; Manchope, M.F.; Ferraz, C.R.; et al. The superoxide anion donor, potassium superoxide, induces pain and inflammation in mice through production of reactive oxygen species and cyclooxygenase-2. Braz. J. Med. Biol. Res. 2015, 48, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Khattab, M.M. TEMPOL, a membrane-permeable radical scavenger, attenuates peroxynitrite- and superoxide anion-enhanced carrageenan-induced paw edema and hyperalgesia: A key role for superoxide anion. Eur. J. Pharmacol. 2006, 548, 167–173. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Porreca, F.; Cuzzocrea, S.; Galen, K.; Lightfoot, R.; Masini, E.; Muscoli, C.; Mollace, V.; Ndengele, M.; Ischiropoulos, H.; et al. A newly identified role for superoxide in inflammatory pain. J. Pharmacol. Exp. Ther. 2004, 309, 869–878. [Google Scholar] [CrossRef]
- Zarpelon, A.C.; Cunha, T.M.; Alves-Filho, J.C.; Pinto, L.G.; Ferreira, S.H.; McInnes, I.B.; Xu, D.; Liew, F.Y.; Cunha, F.Q.; Verri, W.A., Jr. IL-33/ST2 signalling contributes to carrageenin-induced innate inflammation and inflammatory pain: Role of cytokines, endothelin-1 and prostaglandin E2. Br. J. Pharmacol. 2013, 169, 90–101. [Google Scholar] [CrossRef]
- Verri, W.A., Jr.; Guerrero, A.T.; Fukada, S.Y.; Valerio, D.A.; Cunha, T.M.; Xu, D.; Ferreira, S.H.; Liew, F.Y.; Cunha, F.Q. IL-33 mediates antigen-induced cutaneous and articular hypernociception in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 2723–2728. [Google Scholar] [CrossRef]
- Zarpelon, A.C.; Rodrigues, F.C.; Lopes, A.H.; Souza, G.R.; Carvalho, T.T.; Pinto, L.G.; Xu, D.; Ferreira, S.H.; Alves-Filho, J.C.; McInnes, I.B.; et al. Spinal cord oligodendrocyte-derived alarmin IL-33 mediates neuropathic pain. FASEB J. 2016, 30, 54–65. [Google Scholar] [CrossRef]
- Liu, B.; Tai, Y.; Achanta, S.; Kaelberer, M.M.; Caceres, A.I.; Shao, X.; Fang, J.; Jordt, S.E. IL-33/ST2 signaling excites sensory neurons and mediates itch response in a mouse model of poison ivy contact allergy. Proc. Natl. Acad. Sci. USA 2016, 113, E7572–E7579. [Google Scholar] [CrossRef]
- Verri, W.A., Jr.; Souto, F.O.; Vieira, S.M.; Almeida, S.C.; Fukada, S.Y.; Xu, D.; Alves-Filho, J.C.; Cunha, T.M.; Guerrero, A.T.; Mattos-Guimaraes, R.B.; et al. IL-33 induces neutrophil migration in rheumatoid arthritis and is a target of anti-TNF therapy. Ann. Rheum. Dis. 2010, 69, 1697–1703. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed]
- Poole, S.; Cunha, F.Q.; Selkirk, S.; Lorenzetti, B.B.; Ferreira, S.H. Cytokine-mediated inflammatory hyperalgesia limited by interleukin-10. Br. J. Pharmacol. 1995, 115, 684–688. [Google Scholar] [CrossRef]
- Keeble, J.E.; Bodkin, J.V.; Liang, L.; Wodarski, R.; Davies, M.; Fernandes, E.S.; Coelho Cde, F.; Russell, F.; Graepel, R.; Muscara, M.N.; et al. Hydrogen peroxide is a novel mediator of inflammatory hyperalgesia, acting via transient receptor potential vanilloid 1-dependent and independent mechanisms. Pain 2009, 141, 135–142. [Google Scholar] [CrossRef]
- Ghanem, K.Z.; Mahran, M.Z.; Ramadan, M.M.; Ghanem, H.Z.; Fadel, M.; Mahmoud, M.H. A comparative study on flavour components and therapeutic properties of unfermented and fermented defatted soybean meal extract. Sci. Rep. 2020, 10, 5998. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kang, S.A. Antioxidant and Anti-Diabetic Activities of Soybean Fermented with Monascus. Korean J. Food Nutr. 2021, 34, 187–195. [Google Scholar]
- Kumazawa, S.; Taniguchi, M.; Suzuki, Y.; Shimura, M.; Kwon, M.S.; Nakayama, T. Antioxidant activity of polyphenols in carob pods. J. Agric. Food Chem. 2002, 50, 373–377. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Fattori, V.; Manchope, M.F.; Mizokami, S.S.; Casagrande, R.; Verri, W.A., Jr. Naringenin reduces inflammatory pain in mice. Neuropharmacology 2016, 105, 508–519. [Google Scholar] [CrossRef]
- Mizokami, S.S.; Hohmann, M.S.; Staurengo-Ferrari, L.; Carvalho, T.T.; Zarpelon, A.C.; Possebon, M.I.; de Souza, A.R.; Veneziani, R.C.; Arakawa, N.S.; Casagrande, R.; et al. Pimaradienoic Acid Inhibits Carrageenan-Induced Inflammatory Leukocyte Recruitment and Edema in Mice: Inhibition of Oxidative Stress, Nitric Oxide and Cytokine Production. PLoS ONE 2016, 11, e0149656. [Google Scholar] [CrossRef]
- Zucoloto, A.Z.; Manchope, M.F.; Staurengo-Ferrari, L.; Alves-Filho, J.C.; Cunha, T.M.; Antunes, M.M.; Menezes, G.B.; Cunha, F.Q.; Casagrande, R.; Verri, W.A., Jr. Probucol attenuates overt pain-like behavior and carrageenan-induced inflammatory hyperalgesia and leukocyte recruitment by inhibiting NF-small ka, CyrillicB activation and cytokine production without antioxidant effects. Inflamm. Res. 2017, 66, 591–602. [Google Scholar] [CrossRef]
- Handa, C.L.; de Lima, F.S.; Guelfi, M.F.G.; Fernandes, M.D.S.; Georgetti, S.R.; Ida, E.I. Parameters of the fermentation of soybean flour by Monascus purpureus or Aspergillus oryzae on the production of bioactive compounds and antioxidant activity. Food Chem. 2019, 271, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.P.; Bui, T.; Muller, W.A.; Butin-Israeli, V.; Sumagin, R. In vivo imaging reveals unique neutrophil transendothelial migration patterns in inflamed intestines. Mucosal Immunol. 2018, 11, 1571–1581. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Caiado, F.; Pietras, E.M.; Manz, M.G. Inflammation as a regulator of hematopoietic stem cell function in disease, aging, and clonal selection. J. Exp. Med. 2021, 218, e20201541. [Google Scholar] [CrossRef]
- Ungprasert, P.; Cheungpasitporn, W.; Crowson, C.S.; Matteson, E.L. Individual non-steroidal anti-inflammatory drugs and risk of acute kidney injury: A systematic review and meta-analysis of observational studies. Eur. J. Intern. Med. 2015, 26, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Labianca, R.; Sarzi-Puttini, P.; Zuccaro, S.M.; Cherubino, P.; Vellucci, R.; Fornasari, D. Adverse effects associated with non-opioid and opioid treatment in patients with chronic pain. Clin. Drug Investig. 2012, 32 (Suppl. S1), 53–63. [Google Scholar] [CrossRef]
- Patil, K.R.; Mahajan, U.B.; Unger, B.S.; Goyal, S.N.; Belemkar, S.; Surana, S.J.; Ojha, S.; Patil, C.R. Animal Models of Inflammation for Screening of Anti-inflammatory Drugs: Implications for the Discovery and Development of Phytopharmaceuticals. Int. J. Mol. Sci. 2019, 20, 4367. [Google Scholar] [CrossRef]
- Panthong, A.; Norkaew, P.; Kanjanapothi, D.; Taesotikul, T.; Anantachoke, N.; Reutrakul, V. Anti-inflammatory, analgesic and antipyretic activities of the extract of gamboge from Garcinia hanburyi Hook f. J. Ethnopharmacol. 2007, 111, 335–340. [Google Scholar] [CrossRef]
- Tsai, D.S.; Huang, M.H.; Tsai, J.C.; Chang, Y.S.; Chiu, Y.J.; Lin, Y.C.; Wu, L.Y.; Peng, W.H. Analgesic and Anti-Inflammatory Activities of Rosa taiwanensis Nakai in Mice. J. Med. Food 2015, 18, 592–600. [Google Scholar] [CrossRef]
- Lopes, A.H.; Silva, R.L.; Fonseca, M.D.; Gomes, F.I.; Maganin, A.G.; Ribeiro, L.S.; Marques, L.M.M.; Cunha, F.Q.; Alves-Filho, J.C.; Zamboni, D.S.; et al. Molecular basis of carrageenan-induced cytokines production in macrophages. Cell Commun. Signal. 2020, 18, 141. [Google Scholar] [CrossRef]
- Cunha, T.M.; Verri, W.A., Jr.; Schivo, I.R.; Napimoga, M.H.; Parada, C.A.; Poole, S.; Teixeira, M.M.; Ferreira, S.H.; Cunha, F.Q. Crucial role of neutrophils in the development of mechanical inflammatory hypernociception. J. Leukoc. Biol. 2008, 83, 824–832. [Google Scholar] [CrossRef]
- Guerrero, A.T.; Verri, W.A., Jr.; Cunha, T.M.; Silva, T.A.; Schivo, I.R.; Dal-Secco, D.; Canetti, C.; Rocha, F.A.; Parada, C.A.; Cunha, F.Q.; et al. Involvement of LTB4 in zymosan-induced joint nociception in mice: Participation of neutrophils and PGE2. J. Leukoc. Biol. 2008, 83, 122–130. [Google Scholar] [CrossRef]
- Qian, Y.; Guan, T.; Huang, M.; Cao, L.; Li, Y.; Cheng, H.; Jin, H.; Yu, D. Neuroprotection by the soy isoflavone, genistein, via inhibition of mitochondria-dependent apoptosis pathways and reactive oxygen induced-NF-kappaB activation in a cerebral ischemia mouse model. Neurochem. Int. 2012, 60, 759–767. [Google Scholar] [CrossRef]
- Khan, A.Q.; Khan, R.; Rehman, M.U.; Lateef, A.; Tahir, M.; Ali, F.; Sultana, S. Soy isoflavones (daidzein & genistein) inhibit 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced cutaneous inflammation via modulation of COX-2 and NF-kappaB in Swiss albino mice. Toxicology 2012, 302, 266–274. [Google Scholar] [CrossRef]
- Choi, E.J.; Kim, G.H. The antioxidant activity of daidzein metabolites, Odesmethylangolensin and equol, in HepG2 cells. Mol. Med. Rep. 2014, 9, 328–332. [Google Scholar] [CrossRef]
- McCue, P.; Shetty, K. Role of Carbohydrate-Cleaving Enzymes in Phenolic Antioxidant Mobilization from Whole Soybean Fermented with Rhizopus oligosporus. Food Biotechnol. 2003, 17, 27–37. [Google Scholar] [CrossRef]
- Pahl, H.L. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Choi, Y.S.; Park, J.A.; Kim, J.; Rho, S.S.; Park, H.; Kim, Y.M.; Kwon, Y.G. Nuclear IL-33 is a transcriptional regulator of NF-kappaB p65 and induces endothelial cell activation. Biochem. Biophys. Res. Commun. 2012, 421, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Pinto, S.M.; Subbannayya, Y.; Rex, D.A.B.; Raju, R.; Chatterjee, O.; Advani, J.; Radhakrishnan, A.; Keshava Prasad, T.S.; Wani, M.R.; Pandey, A. A network map of IL-33 signaling pathway. J. Cell Commun. Signal. 2018, 12, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Driessler, F.; Venstrom, K.; Sabat, R.; Asadullah, K.; Schottelius, A.J. Molecular mechanisms of interleukin-10-mediated inhibition of NF-kappaB activity: A role for p50. Clin. Exp. Immunol. 2004, 135, 64–73. [Google Scholar] [CrossRef]
- Wang, P.; Wu, P.; Siegel, M.I.; Egan, R.W.; Billah, M.M. Interleukin (IL)-10 inhibits nuclear factor kappa B (NF kappa B) activation in human monocytes. IL-10 and IL-4 suppress cytokine synthesis by different mechanisms. J. Biol. Chem. 1995, 270, 9558–9563. [Google Scholar] [CrossRef]
- Chen, S.; Chen, B.; Wen, Z.; Huang, Z.; Ye, L. IL-33/ST2-mediated inflammation in macrophages is directly abrogated by IL-10 during rheumatoid arthritis. Oncotarget 2017, 8, 32407–32418. [Google Scholar] [CrossRef]
- Siqueira Mietto, B.; Kroner, A.; Girolami, E.I.; Santos-Nogueira, E.; Zhang, J.; David, S. Role of IL-10 in Resolution of Inflammation and Functional Recovery after Peripheral Nerve Injury. J. Neurosci. 2015, 35, 16431–16442. [Google Scholar] [CrossRef]
- Harden, L.M.; Rummel, C.; Luheshi, G.N.; Poole, S.; Gerstberger, R.; Roth, J. Interleukin-10 modulates the synthesis of inflammatory mediators in the sensory circumventricular organs: Implications for the regulation of fever and sickness behaviors. J. Neuroinflamm. 2013, 10, 22. [Google Scholar] [CrossRef]
- Sun, L.; Guo, R.F.; Newstead, M.W.; Standiford, T.J.; Macariola, D.R.; Shanley, T.P. Effect of IL-10 on neutrophil recruitment and survival after Pseudomonas aeruginosa challenge. Am. J. Respir. Cell Mol. Biol. 2009, 41, 76–84. [Google Scholar] [CrossRef]
- Latorre, E.; Matheus, N.; Layunta, E.; Alcalde, A.I.; Mesonero, J.E. IL-10 counteracts proinflammatory mediator evoked oxidative stress in Caco-2 cells. Mediat. Inflamm. 2014, 2014, 982639. [Google Scholar] [CrossRef]
- Kaur, K.; Sharma, A.K.; Dhingra, S.; Singal, P.K. Interplay of TNF-alpha and IL-10 in regulating oxidative stress in isolated adult cardiac myocytes. J. Mol. Cell. Cardiol. 2006, 41, 1023–1030. [Google Scholar] [CrossRef]
- Gregory, N.S.; Harris, A.L.; Robinson, C.R.; Dougherty, P.M.; Fuchs, P.N.; Sluka, K.A. An overview of animal models of pain: Disease models and outcome measures. J. Pain 2013, 14, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, M.M.; Davoli-Ferreira, M.; Santa-Cecilia, F.; Guimaraes, R.M.; Oliveira, F.F.B.; Kusuda, R.; Ferreira, D.W.; Alves-Filho, J.C.; Cunha, F.Q.; Cunha, T.M. IL-27 Counteracts Neuropathic Pain Development Through Induction of IL-10. Front. Immunol. 2019, 10, 3059. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.R.; Rasquel-Oliveira, F.S.; Borghi, S.M.; Franciosi, A.; Carvalho, T.T.; Saraiva-Santos, T.; Artero, N.A.; Casagrande, R.; Verri, W.A.J. Interlinking interleukin-33 (IL-33), neuroinflammation and neuropathic pain. In The Neurobiology, Physiology, and Psychology of Pain; Rajendram, R., Patel, V.B., Preedy, V.R., Martin, C.R., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 171–181. [Google Scholar]
- Jones, M.R.; Urits, I.; Wolf, J.; Corrigan, D.; Colburn, L.; Peterson, E.; Williamson, A.; Viswanath, O. Drug-Induced Peripheral Neuropathy: A Narrative Review. Curr. Clin. Pharmacol. 2020, 15, 38–48. [Google Scholar] [CrossRef]
- Shin, S.A.; Joo, B.J.; Lee, J.S.; Ryu, G.; Han, M.; Kim, W.Y.; Park, H.H.; Lee, J.H.; Lee, C.S. Phytochemicals as Anti-Inflammatory Agents in Animal Models of Prevalent Inflammatory Diseases. Molecules 2020, 25, 5932. [Google Scholar] [CrossRef] [PubMed]
- Schilrreff, P.; Alexiev, U. Chronic Inflammation in Non-Healing Skin Wounds and Promising Natural Bioactive Compounds Treatment. Int. J. Mol. Sci. 2022, 23, 4928. [Google Scholar] [CrossRef]
- Patil, T.; Soni, A.; Acharya, S. A brief review on in vivo models for Gouty Arthritis. Metabol. Open 2021, 11, 100100. [Google Scholar] [CrossRef]
- Jang, C.H.; Oh, J.; Lim, J.S.; Kim, H.J.; Kim, J.S. Fermented Soy Products: Beneficial Potential in Neurodegenerative Diseases. Foods 2021, 10, 636. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DSF-Non | Mean | SD |
|---|---|---|
| Total phenolic content | 7972 | 0.168 |
| Daidzin | 25,489 | 0.056 |
| Glycitin | 5946 | 0.557 |
| Genistin | 32,684 | 0.375 |
| Malonyl genistin | 6930 | 0.230 |
| Dadzein | 7257 | 0.905 |
| Genistein | 3738 | 0.009 |
| Total isoflavones | 82,043 | 2132 |
| DSFF-Mp | Mean | SD |
|---|---|---|
| Total phenolic content | 10,366 | 0.296 |
| Malonyl daidzin | 4595 | 0.132 |
| Malonyl genistin | 7481 | 0.255 |
| Acetyl genistin | 0.7287 | 0.100 |
| Dadzein | 30,420 | 0.968 |
| Genistein | 32,815 | 0.050 |
| Total isoflavones | 76,040 | 1505 |
| DSFF-Ao | Mean | SD |
|---|---|---|
| Total phenolic content | 31,132 | 0.328 |
| Dadzein | 25,130 | 0.473 |
| Genistein | 13,196 | 0.257 |
| Total isoflavones | 38,326 | 0.731 |
| DSFF-Mp | Mean | SD |
|---|---|---|
| Glucose | 1.21 | 0.02 |
| Fructose | 5.96 | 0.20 |
| Sucrose | 334.01 | 19.95 |
| Raffinose | 9.49 | 0.39 |
| Stachyose | 8.08 | 0.26 |
| DSFF-Ao | Mean | SD |
|---|---|---|
| Galactose | 11.88 | 2.10 |
| Glucose | 10.95 | 0.93 |
| Fructose | 3.38 | 0.65 |
| Sucrose | 29.70 | 0.25 |
| Raffinose | 1.23 | 0.32 |
| Stachyose | 7.50 | 0.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manchope, M.F.; Bertozzi, M.M.; Borghi, S.M.; Handa, C.L.; Queiroz-Cancian, M.A.; Ferraz, C.R.; Mizokami, S.S.; Badaró-Garcia, S.; Andrade, K.C.; Zaninelli, T.H.; et al. Fermented (By Monascus purpureus or Aspergillus oryzae) and Non-Fermented Defatted Soybean Flour Extracts: Biological Insight and Mechanism Differences in Inflammatory Pain and Peritonitis. Fermentation 2023, 9, 167. https://doi.org/10.3390/fermentation9020167
Manchope MF, Bertozzi MM, Borghi SM, Handa CL, Queiroz-Cancian MA, Ferraz CR, Mizokami SS, Badaró-Garcia S, Andrade KC, Zaninelli TH, et al. Fermented (By Monascus purpureus or Aspergillus oryzae) and Non-Fermented Defatted Soybean Flour Extracts: Biological Insight and Mechanism Differences in Inflammatory Pain and Peritonitis. Fermentation. 2023; 9(2):167. https://doi.org/10.3390/fermentation9020167
Chicago/Turabian StyleManchope, Marília F., Mariana M. Bertozzi, Sergio M. Borghi, Cíntia L. Handa, Mariana A. Queiroz-Cancian, Camila R. Ferraz, Sandra S. Mizokami, Stephanie Badaró-Garcia, Ketlem C. Andrade, Tiago H. Zaninelli, and et al. 2023. "Fermented (By Monascus purpureus or Aspergillus oryzae) and Non-Fermented Defatted Soybean Flour Extracts: Biological Insight and Mechanism Differences in Inflammatory Pain and Peritonitis" Fermentation 9, no. 2: 167. https://doi.org/10.3390/fermentation9020167