
UDP-Glycosyltransferases in Edible Fungi: Function, Structure, and Catalytic Mechanism
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. UDP-Glycosyltransferases
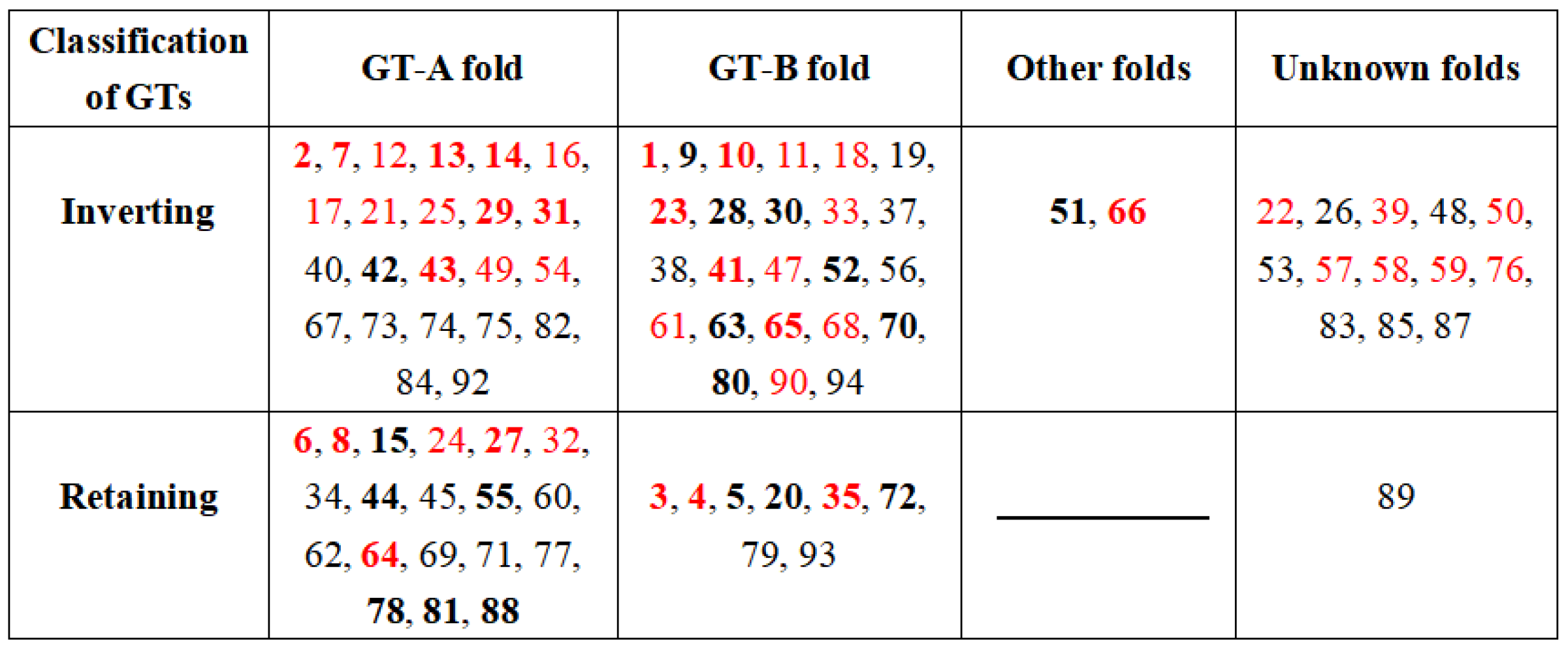
2.1. Donor Recognition and Specificity of UGTs
2.2. Broad Substrate Selection
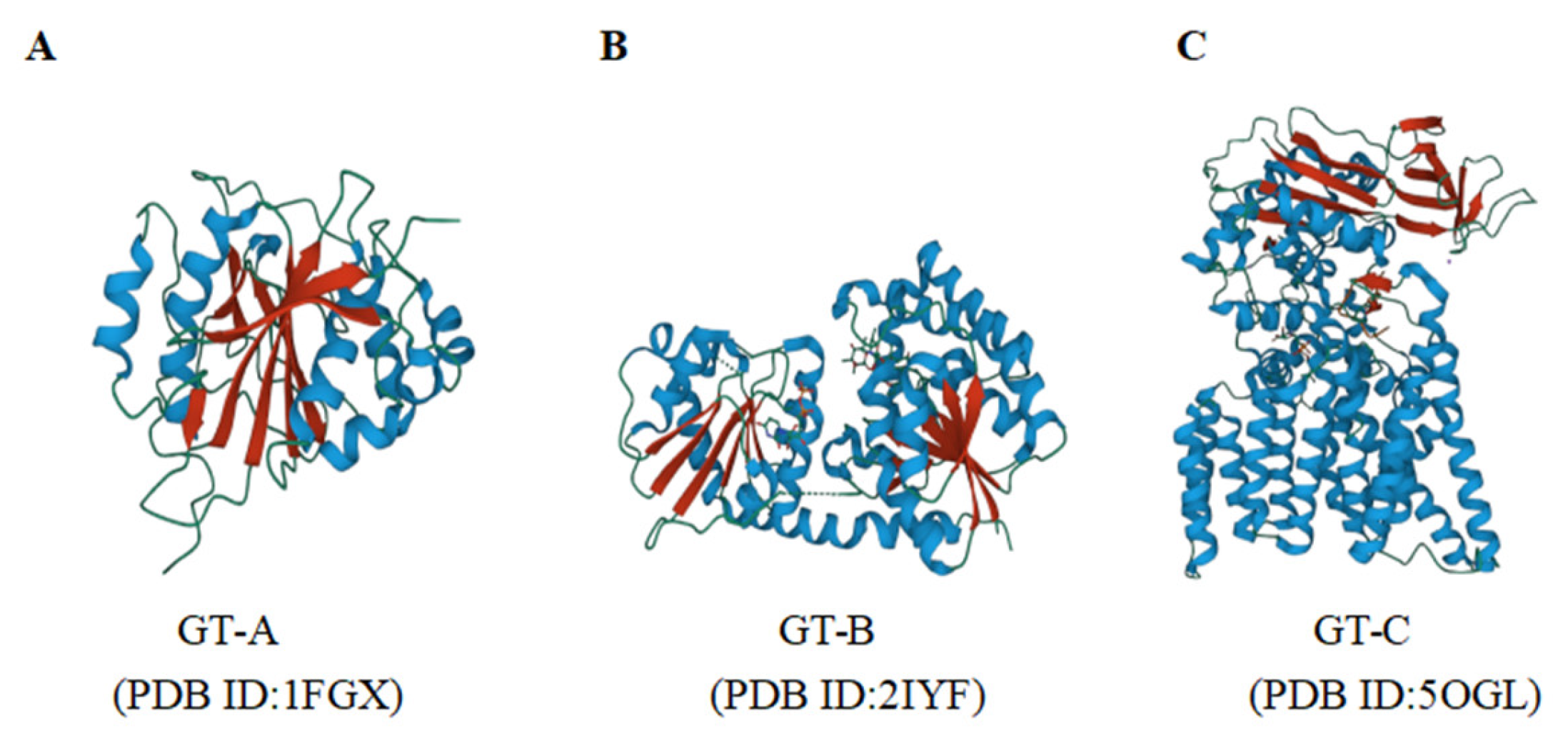
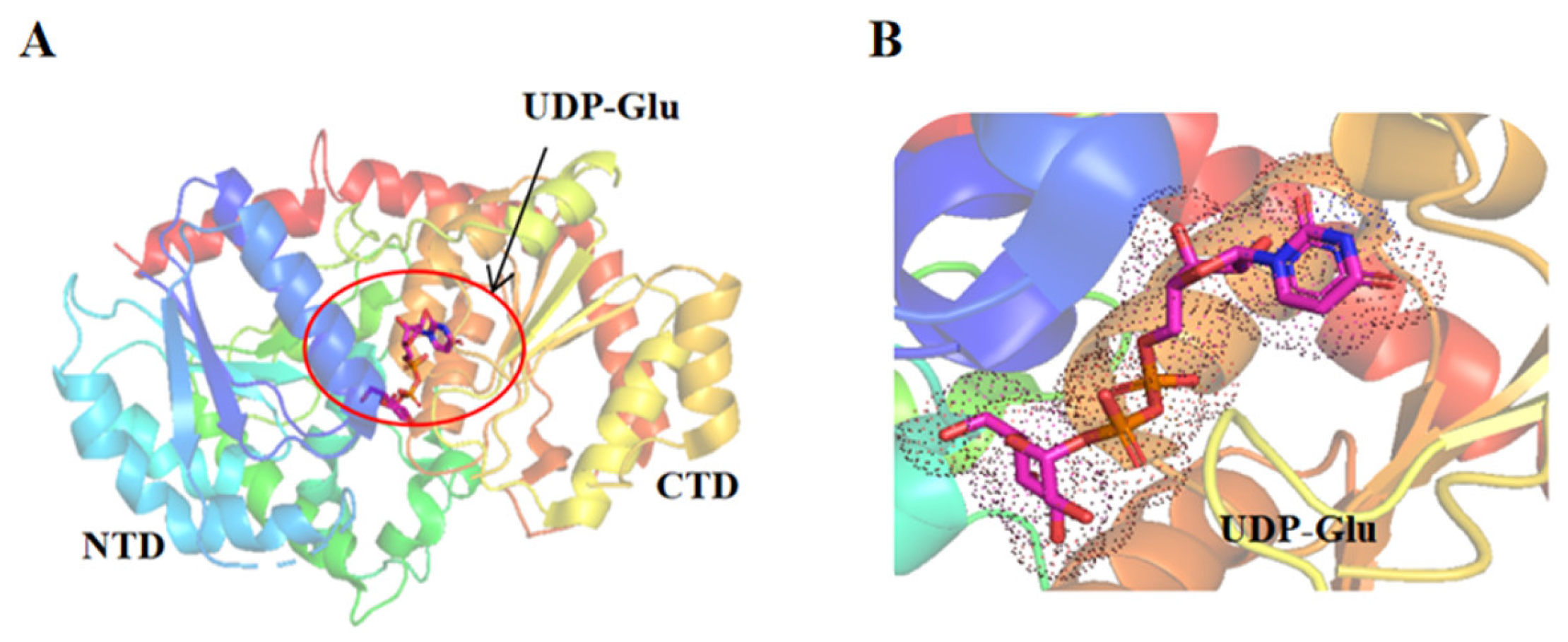
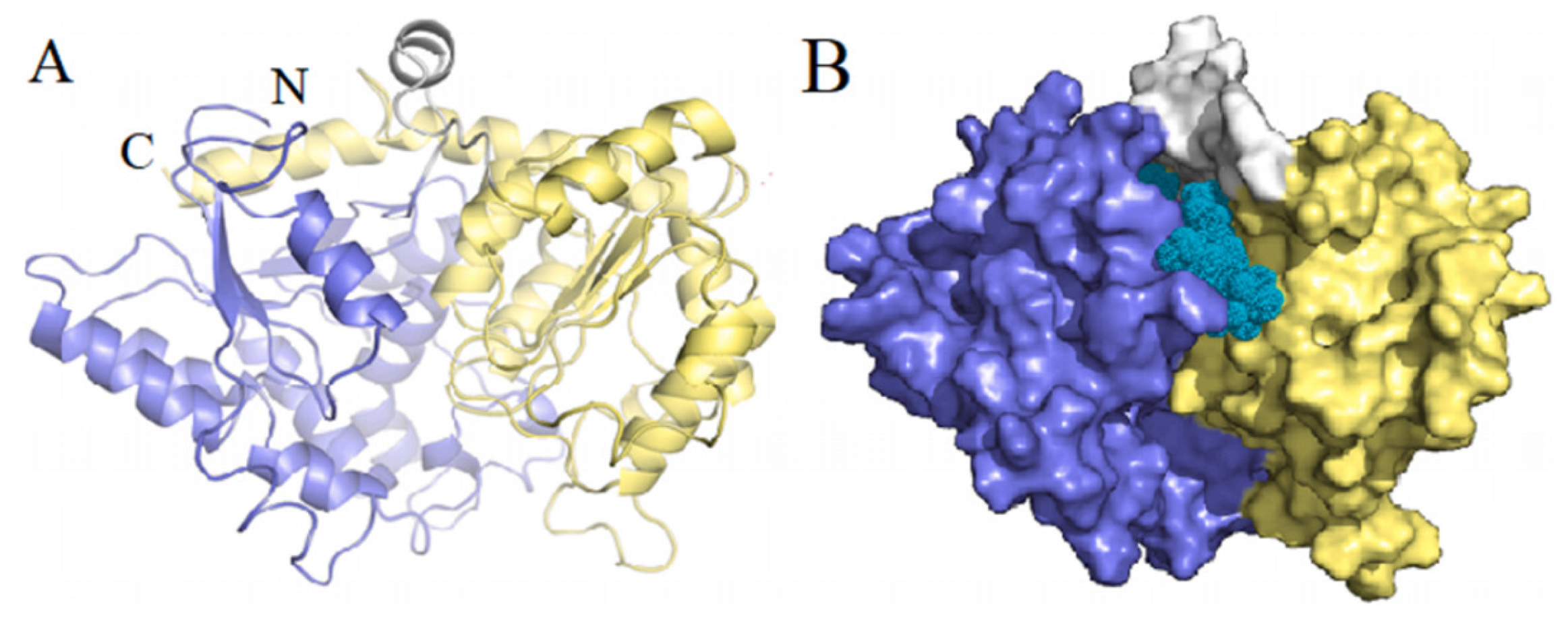
2.3. Structure and Function of UGTs
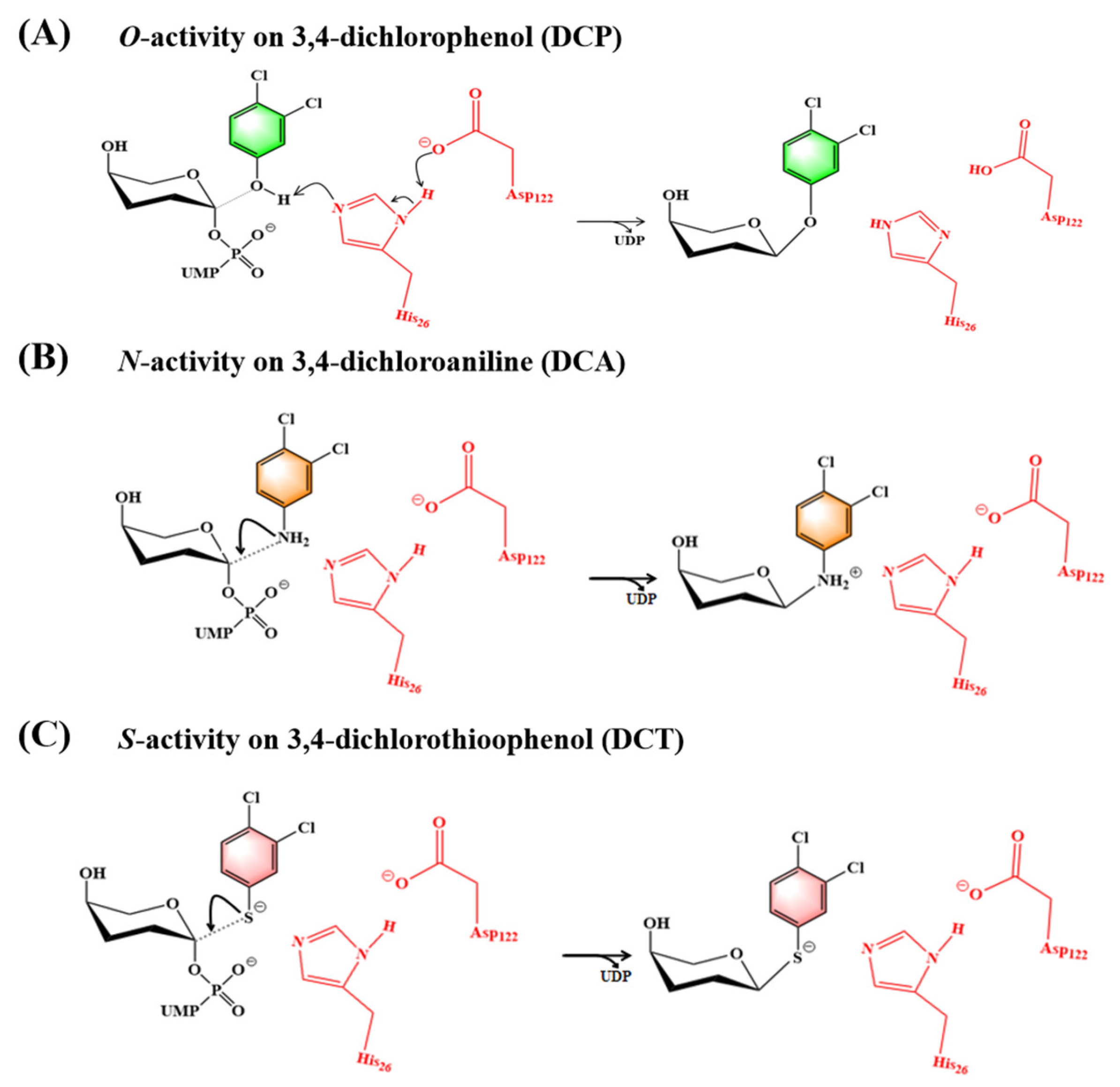
2.4. Catalytic Mechanism
3. Recent Research Status of UDP-Glycosyltransferases
3.1. UDP-Glycosyltransferase in Plants
3.2. UDP-Glycosyltransferases in Bacteria
3.3. UDP-Glycosyltransferase in Yeasts
3.4. UDP-Glycosyltransferases in Edible Fungi
3.4.1. UDP-Glycosyltransferase in Cordyceps militaris
3.4.2. UDP-Glycosyltransferase in Ganoderma lucidum
3.4.3. UDP-Glycosyltransferase in Grifola frondosa
3.4.4. UDP-Glycosyltransferase in Sparassis crispa
3.5. UDP-Glycosyltransferase in Other Fungi
4. Conclusions and Future Perspectives
Funding
Conflicts of Interest
References
- Mackenzie, P.I.; Owens, I.S.; Burchell, B.; Bock, K.W.; Bairoch, A.; Bélanger, A.; Fournel-Gigleux, S.; Green, M.; Hum, D.W.; Iyanagi, T.; et al. The UDP glycosyltransferase gene superfamily: Recommended nomenclature update based on evolutionary divergence. Pharmacogenetics 1997, 7, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.; Isayenkova, J.; Lim, E.K.; Poppenberger, B. Glycosyltransferases: Managers of small molecules. Curr. Opin. Plant Biol. 2005, 8, 254–263. [Google Scholar] [CrossRef]
- Marschall, E.; Cryle, M.J.; Tailhades, J. Biological, chemical, and biochemical strategies for modifying glycopeptide antibiotics. J. Biol. Chem. 2019, 294, 18769–18783. [Google Scholar] [CrossRef] [PubMed]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, functions, and mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, Z.; Zhang, L.; Wang, J.; Wu, C. Glycosyltransferase GT1 family: Phylogenetic distribution, substrates coverage, and representative structural features. Comput. Struct. Biotechnol. J. 2020, 18, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Singh, S.; Phillips Jr, G.N.; Thorson, J.S. Glycosyltransferase structural biology and its role in the design of catalysts for glycosylation. Curr. Opin. Biotechnol. 2011, 22, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Bohl, H.; Bai, L.; Li, H. Recent Progress in Structural Studies on the GT-C Superfamily of Protein Glycosyltransferases. In Macromolecular Protein Complexes III: Structure and Function; Springer: Cham, Switzerland, 2021; pp. 259–271. [Google Scholar]
- Gloster, T.M. Advances in understanding glycosyltransferases from a structural perspective. Curr. Opin. Struct. Biol. 2014, 28, 131–141. [Google Scholar] [CrossRef]
- Gantt, R.W.; Peltier-Pain, P.; Singh, S.; Zhou, M.; Thorson, J.S. Broadening the scope of glycosyltransferase-catalyzed sugar nucleotide synthesis. Proc. Natl. Acad. Sci. USA 2013, 110, 7648–7653. [Google Scholar] [CrossRef]
- Feng, Y.; Yao, M.; Wang, Y.; Ding, M.; Zha, J.; Xiao, W.; Yuan, Y. Advances in engineering UDP-sugar supply for recombinant biosynthesis of glycosides in microbes. Biotechnol. Adv. 2020, 41, 107538. [Google Scholar] [CrossRef]
- Liang, D.M.; Liu, J.H.; Wu, H.; Wang, B.B.; Zhu, H.J.; Qiao, J.J. Glycosyltransferases: Mechanisms and applications in natural product development. Chem. Soc. Rev. 2015, 44, 8350–8374. [Google Scholar] [CrossRef]
- Yu, H.; Takeuchi, M.; LeBarron, J.; Kantharia, J.; London, E.; Bakker, H.; Haltiwanger, R.; Li, H.; Takeuchi, H. Notch-modifying xylosyltransferase structures support an SNi-like retaining mechanism. Nat. Chem. Biol. 2015, 11, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Breton, C.; Fournel-Gigleux, S.; Palcic, M.M. Recent structures, evolution and mechanisms of glycosyltransferases. Curr. Opin. Struct. Biol. 2012, 22, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Paquette, S.; Møller, B.L.; Bak, S. On the origin of family 1 plant glycosyltransferases. Phytochemistry 2003, 62, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Teze, D.; Coines, J.; Fredslund, F.; Dubey, K.D.; Bidart, G.N.; Adams, P.D.; Dueber, J.E.; Svensson, B.; Rovira, C.; Welner, D.H. O-/N-/S-Specificity in Glycosyltransferase Catalysis: From Mechanistic Understanding to Engineering. ACS Catal. 2021, 11, 1810–1815. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Hanada, K. An evolutionary view of functional diversity in family 1 glycosyltransferases. Plant J. 2011, 66, 182–193. [Google Scholar] [CrossRef]
- Campbell, J.A.; Davies, G.J.; Bulone, V.; Henrissat, B. A classification of nucleotide-diphospho-sugar glycosyltransferases based on amino acid sequence similarities. Biochem. J. 1997, 326 Pt 3, 929. [Google Scholar] [CrossRef]
- Qin, J.; Sun, C.; Zhang, M.; Wang, Y. Classification, function and evolution of plant UDP-glycosyltransferase. Genom. Appl. Biol. 2018, 37, 440–450. [Google Scholar]
- Wen, C.; Huang, W.; Zhu, X.L.; Li, X.S.; Zhang, F.; Jiang, R.W. UGT74AN1, a permissive glycosyltransferase from Asclepias curassavica for the regiospecific steroid 3-O-glycosylation. Org. Lett. 2018, 20, 534–537. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.; Mu, S.; Shang, N.; Liu, C.; Zhu, Y.; Cai, Y.; Liu, P.; Lin, J.; Liu, W.; et al. Efficient O-glycosylation of triterpenes enabled by protein engineering of plant glycosyltransferase UGT74AC1. ACS Catal. 2020, 10, 3629–3639. [Google Scholar] [CrossRef]
- Takenaka, Y.; Kato, K.; Ogawa-Ohnishi, M.; Tsuruhama, K.; Kajiura, H.; Yagyu, K.; Takeda, A.; Takeda, Y.; Kunieda, T.; Hara-Nishimura, I.; et al. Pectin RG-I rhamnosyltransferases represent a novel plant-specific glycosyltransferase family. Nat. Plants 2018, 4, 669–676. [Google Scholar] [CrossRef]
- He, J.B.; Zhao, P.; Hu, Z.M.; Liu, S.; Kuang, Y.; Zhang, M.; Li, B.; Yun, C.H.; Qiao, X.; Ye, M. Molecular and Structural Characterization of a Promiscuous C-Glycosyltransferase from Trollius chinensis. Angew. Chem. 2019, 131, 11637–11644. [Google Scholar]
- Cai, X.; Taguchi, T.; Wang, H.; Yuki, M.; Tanaka, M.; Gong, K.; Xu, J.; Zhao, Y.; Ichinose, K.; Li, A. Identification of a C-Glycosyltransferase Involved in Medermycin Biosynthesis. ACS Chem. Biol. 2021, 16, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, L.; Jiang, X.; Diao, X.T.; Li, S.; Li, D.D.; Zhang, Z.; Fang, J.Q.; Tang, Y.J.; Wu, D.L.; et al. Docking-guided rational engineering of a macrolide glycosyltransferase glycodiversifies epothilone B. Commun. Biol. 2022, 5, 100. [Google Scholar] [CrossRef] [PubMed]
- El Qaidi, S.; Zhu, C.; McDonald, P.; Roy, A.; Maity, P.K.; Rane, D.; Perera, C.; Hardwidge, P.R. High-throughput screening for bacterial glycosyltransferase inhibitors. Front. Cell. Infect. Microbiol. 2018, 8, 435. [Google Scholar] [CrossRef]
- Kattke, M.D.; Gosschalk, J.E.; Martinez, O.E.; Kumar, G.; Gale, R.T.; Cascio, D.; Sawaya, M.R.; Philips, M.; Brown, E.D.; Clubb, R.T. Structure and mechanism of TagA, a novel membrane-associated glycosyltransferase that produces wall teichoic acids in pathogenic bacteria. PLoS Pathog. 2019, 15, e1007723. [Google Scholar] [CrossRef]
- Asano, M. Various biological functions of carbohydrate chains learned from glycosyltransferase-deficient mice. Exp. Anim. 2020, 69, 261–268. [Google Scholar] [CrossRef]
- Pal, S.; Verma, J.; Mallick, S.; Rastogi, S.K.; Kumar, A.; Ghosh, A.S. Absence of the glycosyltransferase WcaJ in Klebsiella pneumoniae ATCC13883 affects biofilm formation, increases polymyxin resistance and reduces murine macrophage activation. Microbiology 2019, 165, 891–904. [Google Scholar] [CrossRef]
- Jacquin, L.; Reader, S.M.; Boniface, A.; Mateluna, J.; Patalas, I.; Pérez-Jvostov, F.; Hendry, A.P. Parallel and nonparallel behavioural evolution in response to parasitism and predation in Trinidadian guppies. J. Evol. Biol. 2016, 29, 1406–1422. [Google Scholar] [CrossRef]
- Yu, J.; Hu, F.; Dossa, K.; Wang, Z.; Ke, T. Genome-wide analysis of UDP-glycosyltransferase super family in Brassica rapa and Brassica oleracea reveals its evolutionary history and functional characterization. BMC Genom. 2017, 18, 474. [Google Scholar] [CrossRef]
- Li, Y.; Li, P.; Wang, Y.; Dong, R.; Yu, H.; Hou, B. Genome-wide identification and phylogenetic analysis of Family-1 UDP glycosyltransferases in maize (Zea mays). Planta 2014, 239, 1265–1279. [Google Scholar] [CrossRef]
- Nair, P.C.; Meech, R.; Mackenzie, P.I.; McKinnon, R.A.; Miners, J.O. Insights into the UDP-sugar selectivities of human UDP-glycosyltransferases (UGT): A molecular modeling perspective. Drug Metab. Rev. 2015, 47, 335–345. [Google Scholar] [PubMed]
- Miley, M.J.; Zielinska, A.K.; Keenan, J.E.; Bratton, S.M.; Radominska-Pandya, A.; Redinbo, M.R. Crystal structure of the cofactor-binding domain of the human phase II drug-metabolism enzyme UDP-glucuronosyltransferase 2B7. J. Mol. Biol. 2007, 369, 498–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, N.; Elliot, D.J.; Lewis, B.C.; Burns, K.; Johnston, M.R.; Mackenzie, P.I.; Miners, J.O. Morphine glucuronidation and glucosidation represent complementary metabolic pathways that are both catalyzed by UDP-glucuronosyltransferase 2B7: Kinetic, inhibition, and molecular modeling studies. J. Pharmacol. Exp. Ther. 2014, 349, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.M.; Nawaz, M.A.; Shah, Z.H.; Ludwig-Müller, J.; Chung, G.; Ahmad, M.Q.; Ynag, S.H.; Lee, S.I. Comparative genomic and transcriptomic analyses of Family-1 UDP glycosyltransferase in three Brassica species and Arabidopsis indicates stress-responsive regulation. Sci. Rep. 2018, 8, 1875. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, A.; Horikawa, M.; Fukui, Y.; Fukuchi-Mizutani, M.; Iuchi-Okada, A.; Ishiguro, M.; Kiso, Y.; Nakayama, T.; Ono, E. Local differentiation of sugar donor specificity of flavonoid glycosyltransferase in Lamiales. Plant Cell 2009, 21, 1556–1572. [Google Scholar] [CrossRef] [PubMed]
- Osmani, S.A.; Bak, S.; Imberty, A.; Olsen, C.E.; Møller, B.L. Catalytic key amino acids and UDP-sugar donor specificity of a plant glucuronosyltransferase, UGT94B1: Molecular modeling substantiated by site-specific mutagenesis and biochemical analyses. Plant Physiol. 2008, 148, 1295–1308. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, X.; Hu, Y.; Wang, Y.; Sun, B.; Chen, T.; Luo, Y.; Zhang, Y.; Li, M.; Liu, Z.; et al. Broaden the sugar donor selectivity of blackberry glycosyltransferase UGT78H2 through residual substitutions. Int. J. Biol. Macromol. 2021, 166, 277–287. [Google Scholar] [CrossRef]
- Na, L.; Li, R.; Chen, X. Recent progress in synthesis of carbohydrates with sugar nucleotide-dependent glycosyltransferases. Curr. Opin. Chem. Biol. 2021, 61, 81–95. [Google Scholar] [CrossRef]
- Wei, T.; Zhao, C.; Quareshy, M.; Wu, N.; Huang, S.; Zhao, Y.; Yang, P.; Mao, D.; Chen, Y. A Glycolipid Glycosyltransferase with Broad Substrate Specificity from the Marine Bacterium “Candidatus Pelagibacter sp.” Strain HTCC7211. Appl. Environ. Microbiol. 2021, 87, e00326-21. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.; Xu, Z.; Li, M.; Chen, K.; Zhang, Y.; Hu, Z.; Zhang, M.; Zhang, Z.; Qiao, X.; et al. Highly promiscuous flavonoid 3-O-glycosyltransferase from Scutellaria baicalensis. Org. Lett. 2019, 21, 2241–2245. [Google Scholar] [CrossRef]
- Meech, R.; Mubarokah, N.; Shivasami, A.; Rogers, A.; Nair, P.C.; Hu, D.G.; Mckinnon, R.A.; Mackenzie, P.I. A novel function for UDP glycosyltransferase 8: Galactosidation of bile acids. Mol. Pharmacol. 2015, 87, 442–450. [Google Scholar] [CrossRef]
- Liang, Y.Y.; Zan, X.Y.; Sun, L.; Fu, X.; Cui, F.J.; Tan, M.; Shao, Z.Y.; Sun, W.J. A uridine diphosphate-glycosyltransferase GFUGT88A1 derived from edible mushroom Grifola frondosa extends oligosaccharide chains. Process Biochem. 2022, 112, 80–91. [Google Scholar] [CrossRef]
- Xie, L.; Cao, Y.; Zhao, Z.; Ren, C.; Xing, M.; Wu, B.; Zhang, B.; Xu, C.; Chen, K.; Li, X. Involvement of MdUGT75B1 and MdUGT71B1 in flavonol galactoside/glucoside biosynthesis in apple fruit. Food Chem. 2020, 312, 126124. [Google Scholar] [CrossRef] [PubMed]
- Moremen, K.W.; Haltiwanger, R.S. Emerging structural insights into glycosyltransferase-mediated synthesis of glycans. Nat. Chem. Biol. 2019, 15, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Hamza, A.; Zhan, C.G.; Thorson, J.S. Assessing the regioselectivity of OleD-catalyzed glycosylation with a diverse set of acceptors. J. Nat. Prod. 2013, 76, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, Z.; Li, Z.F.; Chen, Q.; Li, Y.Y.; Gong, Y.; Yue, X.J.; Sheng, D.H.; Zhang, Y.M.; Wu, C.S.; et al. Phylogeny-guided characterization of glycosyltransferases for epothilone glycosylation. Microb. Biotechnol. 2019, 12, 763–774. [Google Scholar] [CrossRef]
- Chen, K.; Hu, Z.M.; Song, W.; Wang, Z.L.; He, J.B.; Shi, X.M.; Cui, Q.H.; Qiao, X.; Ye, M. Diversity of O-glycosyltransferases contributes to the biosynthesis of flavonoid and triterpenoid glycosides in Glycyrrhiza uralensis. ACS Synth. Biol. 2019, 8, 1858–1866. [Google Scholar] [CrossRef]
- Brandt, W.; Schulze, E.; Liberman-Aloni, R.; Bartelt, R.; Pienkny, S.; Carmeli-Weissberg, M.; Frydman, A.; Eyal, Y. Structural modeling of two plant UDP-dependent sugar-sugar glycosyltransferases reveals a conserved glutamic acid residue that is a hallmark for sugar acceptor recognition. J. Struct. Biol. 2021, 213, 107777. [Google Scholar] [CrossRef]
- Wilson, A.E.; Tian, L. Phylogenomic analysis of UDP-dependent glycosyltransferases provides insights into the evolutionary landscape of glycosylation in plant metabolism. Plant J. 2019, 100, 1273–1288. [Google Scholar] [CrossRef]
- Modolo, L.V.; Li, L.; Pan, H.; Blount, J.W.; Dixon, R.A.; Wang, X. Crystal structures of glycosyltransferase UGT78G1 reveal the molecular basis for glycosylation and deglycosylation of (iso) flavonoids. J. Mol. Biol. 2009, 392, 1292–1302. [Google Scholar] [CrossRef]
- George Thompson, A.M.; Iancu, C.V.; Neet, K.E.; Dean, J.V.; Choe, J.Y. Differences in salicylic acid glucose conjugations by UGT74F1 and UGT74F2 from Arabidopsis thaliana. Sci. Rep. 2017, 7, 46629. [Google Scholar] [CrossRef] [PubMed]
- Zong, G.; Fei, S.; Liu, X.; Li, J.; Gao, Y.; Yang, X.; Wang, X.; Shen, Y. Crystal structures of rhamnosyltransferase UGT89C1 from Arabidopsis thaliana reveal the molecular basis of sugar donor specificity for UDP-β-l-rhamnose and rhamnosylation mechanism. Plant J. 2019, 99, 257–269. [Google Scholar] [PubMed]
- Hiromoto, T.; Honjo, E.; Noda, N.; Tamada, T.; Kazuma, K.; Suzuki, M.; Blaber, M.; Kuroki, R. Structural basis for acceptor-substrate recognition of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase from Clitoria ternatea. Protein Sci. 2015, 24, 395–407. [Google Scholar] [CrossRef]
- Wetterhorn, K.M.; Newmister, S.A.; Caniza, R.K.; Busman, M.; McCormick, S.P.; Berthiller, F.; Adam, G.; Rayment, I. Crystal structure of Os79 (Os04g0206600) from Oryza sativa: A UDP-glucosyltransferase involved in the detoxification of deoxynivalenol. Biochemistry 2016, 55, 6175–6186. [Google Scholar] [CrossRef]
- Bolam, D.N.; Roberts, S.; Proctor, M.R.; Turkenburg, J.P.; Dodson, E.J.; Martinez-Fleites, C.; Yang, M.; Davies, G.J.; Gilbert, H.J. The crystal structure of two macrolide glycosyltransferases provides a blueprint for host cell antibiotic immunity. Proc. Natl. Acad. Sci. USA 2007, 104, 5336–5341. [Google Scholar] [CrossRef]
- Smith, A.D.; Page, B.D.; Collier, A.C.; Coughtrie, M.W. Homology modeling of human uridine-5′-diphosphate-glucuronosyltransferase 1A6 reveals insights into factors influencing substrate and cosubstrate binding. ACS Omega 2020, 5, 6872–6887. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, L.; Qu, W.; Wu, F.; Hu, M.; Xie, W.; Liu, Z.; Wang, C. Insight into tartrate inhibition patterns in vitro and in vivo based on cocrystal structure with UDP-glucuronosyltransferase 2B15. Biochem. Pharmacol. 2020, 172, 113753. [Google Scholar] [CrossRef] [PubMed]
- Moremen, K.W.; Ramiah, A.; Stuart, M.; Steel, J.; Meng, L.; Forouhar, F.; Moniz, H.A.; Gahlay, G.; Gao, Z.; Chapla, D.; et al. Expression system for structural and functional studies of human glycosylation enzymes. Nat. Chem. Biol. 2018, 14, 156–162. [Google Scholar] [CrossRef]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Žídek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly accurate protein structure prediction for the human proteome. Nature 2021, 596, 590–596. [Google Scholar] [CrossRef]
- Pennisi, E. Protein structure prediction now easier, faster. Science 2021, 373, 262–263. [Google Scholar] [CrossRef]
- Yan, Y.; Mo, T.; Huang, W.; Xu, X.; Tian, W.; Wang, Y.; Song, Y.; Li, J.; Shi, S.; Liu, X.; et al. Glycosylation of Aromatic Glycosides by a Promiscuous Glycosyltransferase UGT71BD1 from Cistanche tubulosa. J. Nat. Prod. 2022, 85, 1826–1836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, S.; Yang, J.; Kang, C.; Huang, L.; Guo, L. Glycosylation of plant secondary metabolites: Regulating from chaos to harmony. Environ. Exp. Bot. 2022, 194, 104703. [Google Scholar] [CrossRef]
- Krupicka, M.; Tvaroska, I. Hybrid quantum mechanical/molecular mechanical investigation of the β-1, 4-galactosyltransferase-I mechanism. J. Phys. Chem. B 2009, 113, 11314–11319. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; He, Y.; Jiang, R.; Deng, Z.; Long, F. Functional and Structural Dissection of a Plant Steroid 3-O-Glycosyltransferase Facilitated the Engineering Enhancement of Sugar Donor Promiscuity. ACS Catal. 2022, 12, 2927–2937. [Google Scholar] [CrossRef]
- Wilson, A.E.; Feng, X.; Ono, N.N.; Holland, D.; Amir, R.; Tian, L. Characterization of a UGT84 family glycosyltransferase provides new insights into substrate binding and reactivity of galloylglucose ester-forming UGTs. Biochemistry 2017, 56, 6389–6400. [Google Scholar] [CrossRef]
- Brazier-Hicks, M.; Offen, W.A.; Gershater, M.C.; Revett, T.J.; Lim, E.K.; Bowles, D.J.; Davies, G.J.; Edwards, R. Characterization and engineering of the bifunctional N-and O-glucosyltransferase involved in xenobiotic metabolism in plants. Proc. Natl. Acad. Sci. USA 2007, 104, 20238–20243. [Google Scholar] [CrossRef]
- Zhang, J.J.; Cao, W.; Xi, P.; Li, L.; Qiao, S.; Luo, H.; Zhang, J.; Liu, X.; Du, N. S-Glycosylation of Fluensulfone in Tomatoes: An Important Way of Fluensulfone Metabolism. J. Agric. Food Chem. 2021, 69, 12974–12984. [Google Scholar] [CrossRef]
- Wang, H.; Oman, T.J.; Zhang, R.; Garcia De Gonzalo, C.V.; Zhang, Q.; Van Der Donk, W.A. The glycosyltransferase involved in thurandacin biosynthesis catalyzes both O-and S-glycosylation. J. Am. Chem. Soc. 2014, 136, 84–87. [Google Scholar] [CrossRef]
- Bashyal, P.; Thapa, S.B.; Kim, T.S.; Pandey, R.P.; Sohng, J.K. Exploring the nucleophilic N-and S-glycosylation capacity of Bacillus licheniformis YjiC enzyme. J. Microbiol. Biotechnol. 2020, 30, 1092–1096. [Google Scholar] [CrossRef]
- Sasaki, N.; Nishizaki, Y.; Yamada, E.; Tatsuzawa, F.; Nakatsuka, T.; Takahashi, H.; Nishihara, M. Identification of the glucosyltransferase that mediates direct flavone C-glucosylation in Gentiana triflora. FEBS Lett. 2015, 589, 182–187. [Google Scholar] [CrossRef]
- Wang, X.; Li, C.; Zhou, C.; Li, J.; Zhang, Y. Molecular characterization of the C-glucosylation for puerarin biosynthesis in Pueraria lobata. Plant J. 2017, 90, 535–546. [Google Scholar] [CrossRef]
- Liu, M.; Wang, D.; Li, Y.; Li, X.; Zong, G.; Fei, S.; Yang, X.; Lin, J.; Wang, X.; Shen, Y. Crystal structures of the C-glycosyltransferase UGT708C1 from buckwheat provide insights into the mechanism of C-glycosylation. Plant Cell 2020, 32, 2917–2931. [Google Scholar] [CrossRef]
- Ito, T.; Fujimoto, S.; Suito, F.; Shimosaka, M.; Taguchi, G. C-Glycosyltransferases catalyzing the formation of di-C-glucosyl flavonoids in citrus plants. Plant J. 2017, 91, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Fan, S.; Chen, R.; Xie, K.; Yin, S.; Sun, L.; Liu, J.; Yang, L.; Kong, J.; Yang, Z.; et al. Probing and engineering key residues for bis-C-glycosylation and promiscuity of a C-glycosyltransferase. ACS Catal. 2018, 8, 4917–4927. [Google Scholar] [CrossRef]
- Dai, L.; Hu, Y.; Chen, C.C.; Ma, L.; Guo, R.T. Flavonoid C-Glycosyltransferases: Function, Evolutionary Relationship, Catalytic Mechanism and Protein Engineering. ChemBioEng Rev. 2021, 8, 15–26. [Google Scholar] [CrossRef]
- Chen, D.; Chen, R.; Wang, R.; Li, J.; Xie, K.; Bian, C.; Sun, L.; Zhang, Z.; Liu, J.; Yang, L.; et al. Probing the catalytic promiscuity of a regio-and stereospecific C-glycosyltransferase from Mangifera indica. Angew. Chem. 2015, 127, 12869–12873. [Google Scholar] [CrossRef]
- Yi, S.; Kuang, T.; Miao, Y.; Xu, Y.; Wang, Z.; Dong, L.B.; Tan, N. Discovery and characterization of four glycosyltransferases involved in anthraquinone glycoside biosynthesis in Rubia yunnanensis. Org. Chem. Front. 2020, 7, 2442–2448. [Google Scholar] [CrossRef]
- Aoi, Y.; Hira, H.; Hayakawa, Y.; Liu, H.; Fukui, K.; Dai, X.; Tanaka, K.; Hayashi, K.; Zhao, Y.; Kasahara, H. UDP-glucosyltransferase UGT84B1 regulates the levels of indole-3-acetic acid and phenylacetic acid in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 532, 244–250. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, W.; You, W.; Di, Z.; Wang, M.; Zhou, H.; Yuan, S.; Wong, N.; Xiao, Y. Discovery of Arabidopsis UGT73C1 as a steviol-catalyzing UDP-glycosyltransferase with chemical probes. Chem. Commun. 2018, 54, 7179–7182. [Google Scholar] [CrossRef]
- Šmehilová, M.; Dobrůšková, J.; Novák, O.; Takáč, T.; Galuszka, P. Cytokinin-specific glycosyltransferases possess different roles in cytokinin homeostasis maintenance. Front. Plant Sci. 2016, 7, 1264. [Google Scholar] [CrossRef]
- Pandey, R.P.; Parajuli, P.; Sohng, J.K. Metabolic engineering of glycosylated polyketide biosynthesis. Emerg. Top. Life Sci. 2018, 2, 389–403. [Google Scholar] [PubMed]
- Xie, K.; Chen, R.; Li, J.; Wang, R.; Chen, D.; Dou, X.; Dai, J. Exploring the catalytic promiscuity of a new glycosyltransferase from Carthamus tinctorius. Org. Lett. 2014, 16, 4874–4877. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Hayase, H.; Nakayama, A.; Yamaguchi, I.; Asami, T.; Nakajima, M. Identification and characterization of an Ipomoea nil glucosyltransferase which metabolizes some phytohormones. Biochem. Biophys. Res. Commun. 2007, 361, 980–986. [Google Scholar] [CrossRef]
- Maharjan, R.; Fukuda, Y.; Shimomura, N.; Nakayama, T.; Okimoto, Y.; Kawakami, K.; Nakayama, T.; Hamada, H.; Inoue, T.; Ozaki, S. An Ambidextrous Polyphenol Glycosyltransferase Pa GT2 from Phytolacca americana. Biochemistry 2020, 59, 2551–2561. [Google Scholar] [CrossRef]
- Shao, H.; He, X.; Achnine, L.; Blount, J.; Dixon, R.; Wang, X. Crystal structures of a multifunctional triterpene/flavonoid glycosyltransferase from Medicago truncatula. Plant Cell 2005, 17, 3141–3154. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Zhang, Z.M.; Zhong, L.; Fan, J.; Li, M.; Ma, Y.; Zhou, Y.; Zhang, W.; Guo, B.; Chen, B. Directed Evolution of a Plant Glycosyltransferase for Chemo-and Regioselective Glycosylation of Pharmaceutically Significant Flavonoids. ACS Catal. 2021, 11, 14781–14790. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Son, Y.O.; Wang, X.; Divya, S.P.; Joseph, B.; Hitron, J.A.; Wang, L.; Kim, D.; Yin, Y.; Roy, R.V.; et al. Cyanidin-3-glucoside inhibits UVB-induced oxidative damage and inflammation by regulating MAP kinase and NF-κB signaling pathways in SKH-1 hairless mice skin. Toxicol. Appl. Pharmacol. 2014, 280, 127–137. [Google Scholar] [CrossRef]
- Dai, L.; Li, J.; Yao, P.; Zhu, Y.; Men, Y.; Zeng, Y.; Yang, J.G.; Sun, Y. Exploiting the aglycon promiscuity of glycosyltransferase Bs-YjiC from Bacillus subtilis and its application in synthesis of glycosides. J. Biotechnol. 2017, 248, 69–76. [Google Scholar] [CrossRef]
- Chen, K.; He, J.; Hu, Z.; Song, W.; Yu, L.; Li, K.; Qiao, X.; Ye, M. Enzymatic glycosylation of oleanane-type triterpenoids. J. Asian Nat. Prod. Res. 2018, 20, 615–623. [Google Scholar] [CrossRef]
- Li, K.; Feng, J.; Kuang, Y.; Song, W.; Zhang, M.; Ji, S.; Qiao, X.; Ye, M. Enzymatic Synthesis of Bufadienolide O-Glycosides as Potent Antitumor Agents Using a Microbial Glycosyltransferase. Adv. Synth. Catal. 2017, 359, 3765–3772. [Google Scholar] [CrossRef]
- Liang, H.; Hu, Z.; Zhang, T.; Gong, T.; Chen, J.; Zhu, P.; Li, Y.; Yang, J. Production of a bioactive unnatural ginsenoside by metabolically engineered yeasts based on a new UDP-glycosyltransferase from Bacillus subtilis. Metab. Eng. 2017, 44, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Thierbach, S.; Sartor, P.; Yücel, O.; Fetzner, S. Efficient modification of the Pseudomonas aeruginosa toxin 2-heptyl-1-hydroxyquinolin-4-one by three Bacillus glycosyltransferases with broad substrate ranges. J. Biotechnol. 2020, 308, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Bashyal, P.; Parajuli, P.; Yamaguchi, T.; Sohng, J.K. Two trifunctional leloir glycosyltransferases as biocatalysts for natural products glycodiversification. Org. Lett. 2019, 21, 8058–8064. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.Y.; Van Minh, N.; Choi, J.M.; Hwang, J.Y.; Seo, S.T.; Lee, S.K.; Kim, W.G. Enzymatic synthesis of avermectin B1a glycosides for the effective prevention of the pine wood nematode Bursaphelenchus xylophilus. Appl. Microbiol. Biotechnol. 2018, 102, 2155–2165. [Google Scholar] [CrossRef]
- Pandey, R.P.; Chu, L.L.; Kim, T.S.; Sohng, J.K. Bioconversion of tetracycline antibiotics to novel glucoside derivatives by single-vessel multienzymatic glycosylation. J. Microbiol. Biotechnol. 2018, 28, 298–304. [Google Scholar] [CrossRef]
- Pu, X.; He, C.; Yang, Y.; Wang, W.; Hu, K.; Xu, Z.; Song, J. In vivo production of five crocins in the engineered Escherichia coli. ACS Synth. Biol. 2020, 9, 1160–1168. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Y.; Feng, Y. Structural dissection of sterol glycosyltransferase UGT51 from Saccharomyces cerevisiae for substrate specificity. J. Struct. Biol. 2018, 204, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.L.; Montecillo, J.A.V.; Bae, H. Recent advances in the metabolic engineering of yeasts for ginsenoside biosynthesis. Front. Bioeng. Biotechnol. 2020, 8, 139. [Google Scholar] [CrossRef]
- Wang, C.; Liwei, M.; Park, J.B.; Jeong, S.H.; Wei, G.; Wang, Y.; Kim, S.W. Microbial platform for terpenoid production: Escherichia coli and yeast. Front. Microbiol. 2018, 9, 2460. [Google Scholar] [CrossRef]
- Wang, P.; Wei, W.; Ye, W.; Li, X.; Zhao, W.; Yang, C.; Li, C.; Yan, X.; Zhou, Z. Synthesizing ginsenoside Rh2 in Saccharomyces cerevisiae cell factory at high-efficiency. Cell Discov. 2019, 5, 5. [Google Scholar] [CrossRef]
- Yuan, Q.H.; Xie, F.; Tan, J.; Yuan, Y.; Mei, H.; Zheng, Y.R. Extraction, structure and pharmacological effects of the polysaccharides from Cordyceps sinensis: A review. J. Funct. Foods 2022, 89, 104909. [Google Scholar] [CrossRef]
- Felix, G.; Regenass, M.; Boller, T. Specific perception of subnanomolar concentrations of chitin fragments by tomato cells: Induction of extracellular alkalinization, changes in protein phosphorylation, and establishment of a refractory state. Plant J. Cell Mol. Biol. 2010, 4, 307–316. [Google Scholar] [CrossRef]
- Tomazett, P.K.; Félix, C.R.; Lenzi, H.L.; de Paula Faria, F.; de Almeida Soares, C.M.; Pereira, M. 1,3-β-D-Glucan synthase of Paracoccidioides brasiliensis: Recombinant protein, expression and cytolocalization in the yeast and mycelium phases. Fungal Biol. 2010, 114, 809–816. [Google Scholar] [CrossRef]
- Arellano, M.; Duran, A.; Perez, P. Rho 1 GTPase activates the (1, 3) beta-D-glucan synthase and is involved in Schizosaccharomyces pombe morphogenesis. EMBO J. 1996, 15, 4584–4591. [Google Scholar] [CrossRef]
- Okada, H.; Abe, M.; Asakawa-Minemura, M.; Hirata, A.; Qadota, H.; Morishita, K.; Ohnuki, S.; Nogami, S.; Ohya, Y. Multiple functional domains of the yeast l, 3-β-glucan synthase subunit Fks1p revealed by quantitative phenotypic analysis of temperature-sensitive mutants. Genetics 2010, 184, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Ujita, M.; Katsuno, Y.; Suzuki, K.; Sugiyama, K.; Takeda, E.; Hara, A.; Yokoyama, E. Molecular cloning and sequence analysis of the β-1,3-glucan synthase catalytic subunit gene from a medicinal fungus, Cordyceps militaris. Mycoscience 2006, 47, 98–105. [Google Scholar] [CrossRef]
- Fu, X.; Zan, X.Y.; Sun, L.; Tan, M.; Cui, F.J.; Liang, Y.Y.; Meng, L.J.; Sun, W.J. Functional Characterization and Structural Basis of the β-1,3-Glucan Synthase CMGLS from Mushroom Cordyceps militaris. J. Agric. Food Chem. 2022, 70, 8725–8737. [Google Scholar] [CrossRef]
- Liu, D.; Gong, J.; Dai, W.; Kang, X.; Huang, Z.; Zhang, H.M.; Liu, W.; Liu, L.; Ma, J.; Xia, Z.; et al. The genome of Ganderma lucidum provide insights into triterpense biosynthesis and wood degradation. PLoS ONE 2012, 7, e36146. [Google Scholar]
- Achnine, L.; Huhman, D.V.; Farag, M.A.; Sumner, L.W.; Blount, J.W.; Dixon, R.A. Genomics-based selection and functional characterization of triterpene glycosyltransferases from the model legume Medicago truncatula. Plant J. 2005, 41, 875–887. [Google Scholar] [CrossRef]
- Chen, S.; Xu, J.; Liu, C.; Zhu, Y.; Nelson, D.R.; Zhou, S.; Li, C.; Wang, L.; Guo, X.; Sun, Y.; et al. Genome sequence of the model medicinal mushroom Ganoderma lucidum. Nat. Commun. 2012, 3, 913. [Google Scholar] [CrossRef]
- Borchers, A.; Stern, J.; Hackman, R.; Keen, C.; Gershwin, M. Mushrooms, tumors, and immunity. Proc. Soc. Exp. Biol. Med. 1999, 221, 281–293. [Google Scholar] [PubMed]
- Tao, T.L.; Cui, F.J.; Chen, X.X.; Sun, W.J.; Huang, D.M.; Zhang, J.S.; Yang, Y.; Wu, D.; Liu, W.M. Improved mycelia and polysaccharide production of Grifola frondosa by controlling morphology with microparticle Talc. Microb. Cell Fact. 2018, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, F.; Wu, X.; Tao, T.; Zan, X.; Sun, W.; Mu, D.; Yang, Y.; Wu, D. Functions of a glucan synthase gene GFGLS in mycelial growth and polysaccharide production of Grifola frondosa. J. Agric. Food Chem. 2019, 67, 8875–8883. [Google Scholar] [CrossRef]
- Zan, X.; Wu, X.; Cui, F.; Zhu, H.; Sun, W.; Jiang, L.; Tao, T.; Zhao, X. UDP-glucose pyrophosphorylase gene affects mycelia growth and polysaccharide synthesis of Grifola frondosa. Int. J. Biol. Macromol. 2020, 20, 33580–33587. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Li, X.F.; Zan, X.Y.; Fu, X.; Cui, F.J.; Zhu, H.A.; Sun, W.J.; Tao, T.L. The β-1,3-glucan synthase gene GFGLS2 plays major roles in mycelial growth and polysaccharide synthesis in Grifola frondosa. Appl. Microbiol. Biotechnol. 2022, 106, 563–578. [Google Scholar] [CrossRef]
- Yang, Y.H.; Kang, H.W.; Ro, H.S. Cloning and molecular characterization of β-1, 3-glucan synthase from Sparassis crispa. Mycobiology 2014, 42, 167–173. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, P.; Cui, Y.; Li, K.; Qiao, X.; Zhang, Y.T.; Li, S.M.; Cox, R.J.; Wu, B.; Ye, M.; et al. Regio- and stereospecificO-glycosylation of phenolic compounds catalyzed by a fungal glycosyltransferase from Mucor hiemalis. Adv. Synth. Catal. 2017, 359, 995–1006. [Google Scholar] [CrossRef]
- Xie, K.; Dou, X.; Chen, R.; Chen, D.; Fang, C.; Xiao, Z.; Dai, J. Two novel fungal phenolic UDP glycosyltransferases from Absidia coerulea and Rhizopus japonicus. Appl. Environ. Microbiol. 2017, 83, e03103-16. [Google Scholar] [CrossRef]
- Xie, L.N. Exploration and application of the glycosyltransferase-methyltransferase biosynthesis module of B. bassiana. Chin. Acad. Agric. Sci. 2021. [Google Scholar] [CrossRef]
- Schuma, B.; Alfaro, J.; Evans, S. Glycosyltransferase structure and function. Top. Curr. Chem. 2007, 271, 217–257. [Google Scholar]
- Wilson, I.B.; Breton, C.; Imberty, A.; Tvaroška, I. Molecular basis for the biosynthesis of oligo-and polysaccharides. Glycoscience 2008, 11, 2265–2323. [Google Scholar]
- Chen, X.; Wang, Q.Q.; Liu, N.N.; Liu, G.L.; Chi, Z.; Chi, Z.M. A glycosyltransferase gene responsible for pullulan biosynthesis in Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2017, 95, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Engel, J.; Schmalhorst, P.S.; Dörk-Bousset, T.; Ferrieres, V.; Routier, F.H. A single UDP-galactofuranose transporter is required for galactofuranosylation in Aspergillus fumigatus. J. Biol. Chem. 2009, 284, 33859–33868. [Google Scholar] [CrossRef]
- Schmalhorst, P.S.; Krappmann, S.; Vervecken, W.; Rohde, M.; Muller, M.; Braus, G.H.; Contreras, R.; Braun, A.; Bakker, H.; Routier, F.H. Contribution of galactofuranose to the virulence of the opportunistic pathogen Aspergillus fumigatus. Eukaryot. Cell 2008, 7, 1268–1277. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; O’Donnell, H.; Routier, F.H.; Tiralongo, J.; Haselhorst, T. Glycobiology of Human Fungal Pathogens: New Avenues for Drug Development. Cells 2019, 8, 1348. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Liang, Y.; Cui, F.; Wang, Y.; Sun, L.; Zan, X.; Sun, W. UDP-Glycosyltransferases in Edible Fungi: Function, Structure, and Catalytic Mechanism. Fermentation 2023, 9, 164. https://doi.org/10.3390/fermentation9020164
Yang Y, Liang Y, Cui F, Wang Y, Sun L, Zan X, Sun W. UDP-Glycosyltransferases in Edible Fungi: Function, Structure, and Catalytic Mechanism. Fermentation. 2023; 9(2):164. https://doi.org/10.3390/fermentation9020164
Chicago/Turabian StyleYang, Yumeng, Yingying Liang, Fengjie Cui, Yongli Wang, Lei Sun, Xinyi Zan, and Wenjing Sun. 2023. "UDP-Glycosyltransferases in Edible Fungi: Function, Structure, and Catalytic Mechanism" Fermentation 9, no. 2: 164. https://doi.org/10.3390/fermentation9020164