
Okara Waste as a Substrate for the Microalgae Phaeodactylum tricornutum Enhances the Production of Algal Biomass, Fucoxanthin, and Polyunsaturated Fatty Acids
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Culture
2.2. Determination of Algal Growth
2.3. Preparation of Growth Media
2.4. Nutritional Analysis of Fermented Okara Media
2.5. Quantification of Fucoxanthin and Chlorophyll a Production from P. tricornutum
2.6. Lipid Extraction and Yield Measurement
2.7. Fatty Acid Analysis
2.8. Intracellular Metabolite Analysis
2.9. Statistical Analysis
3. Results and Discussion
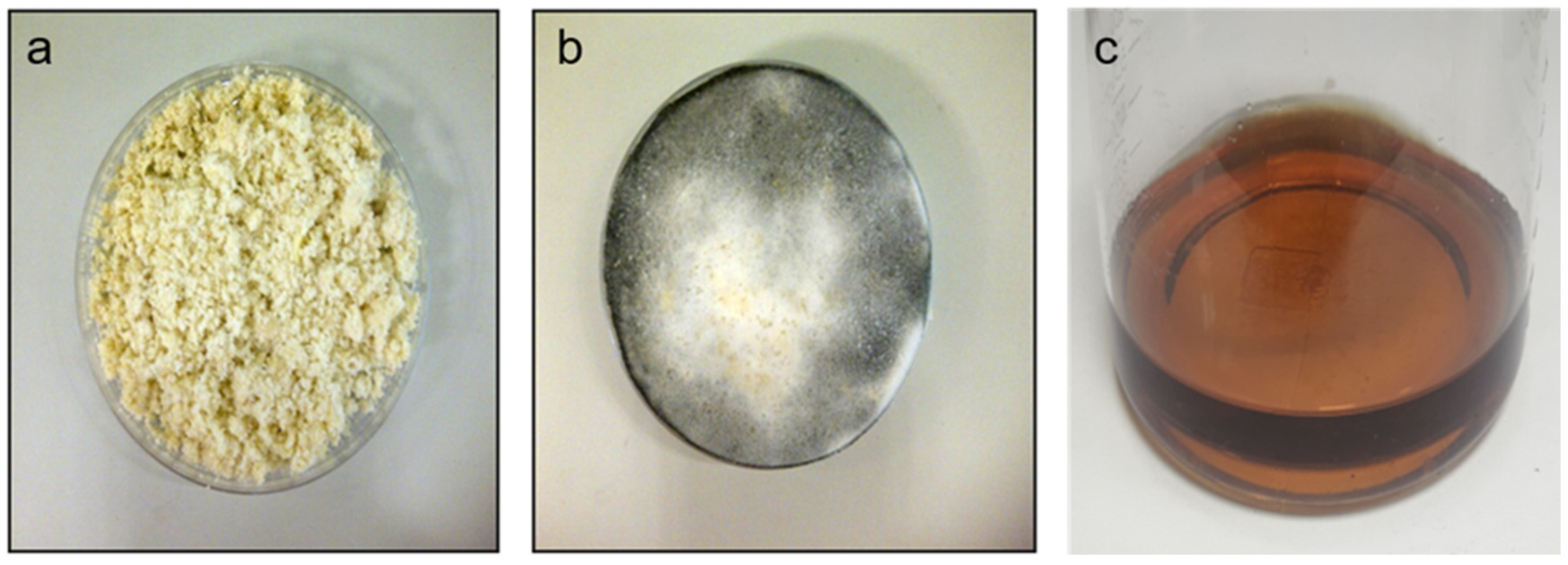
3.1. Okara Fermentation and Media Preparation
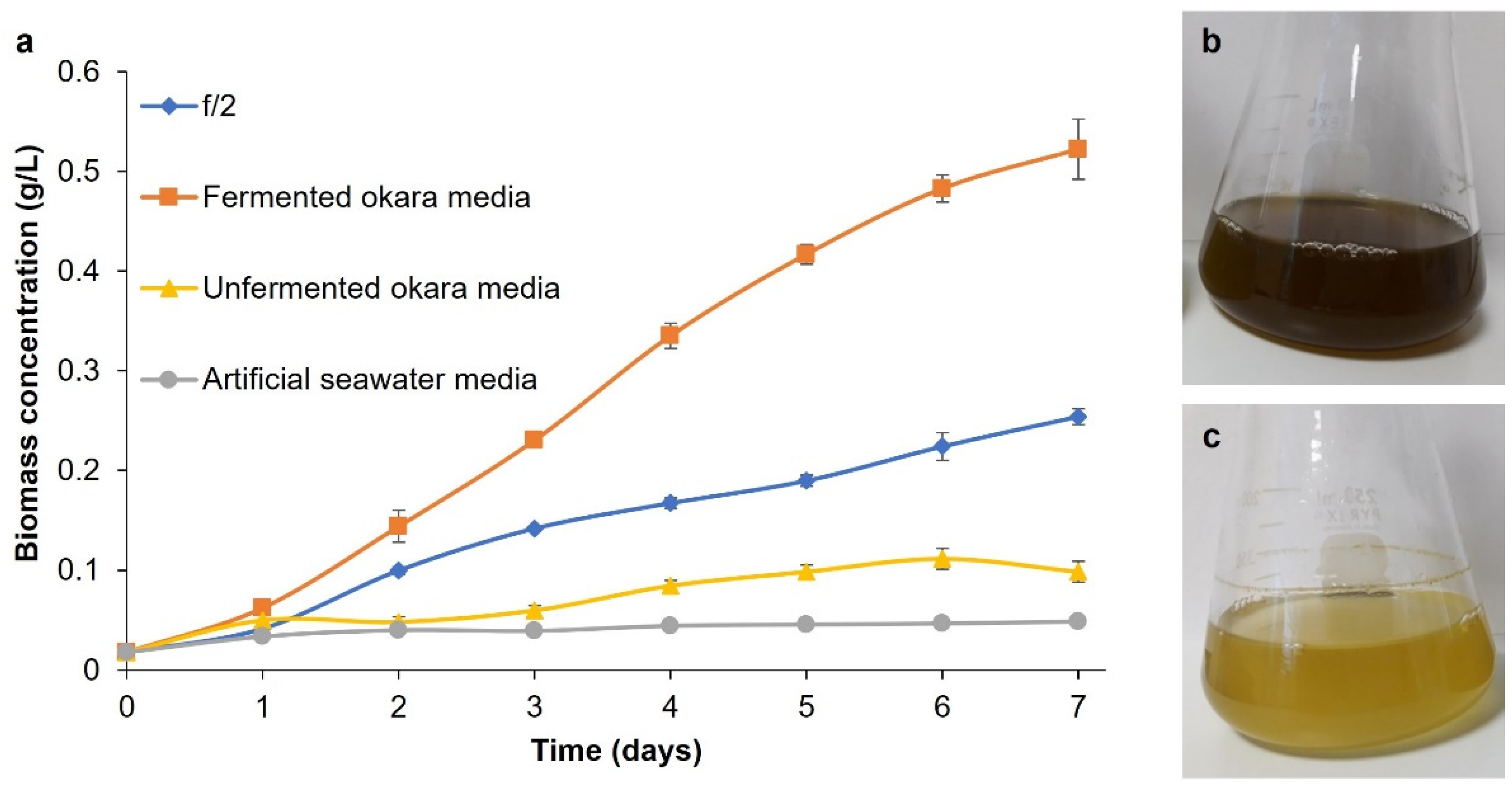
3.2. Growth of P. tricornutum in Fermented Okara Media
3.3. Nutrient Composition in Fermented Okara Media
3.4. Fucoxanthin Production from P. tricornutum Cells in Fermented Okara Culture
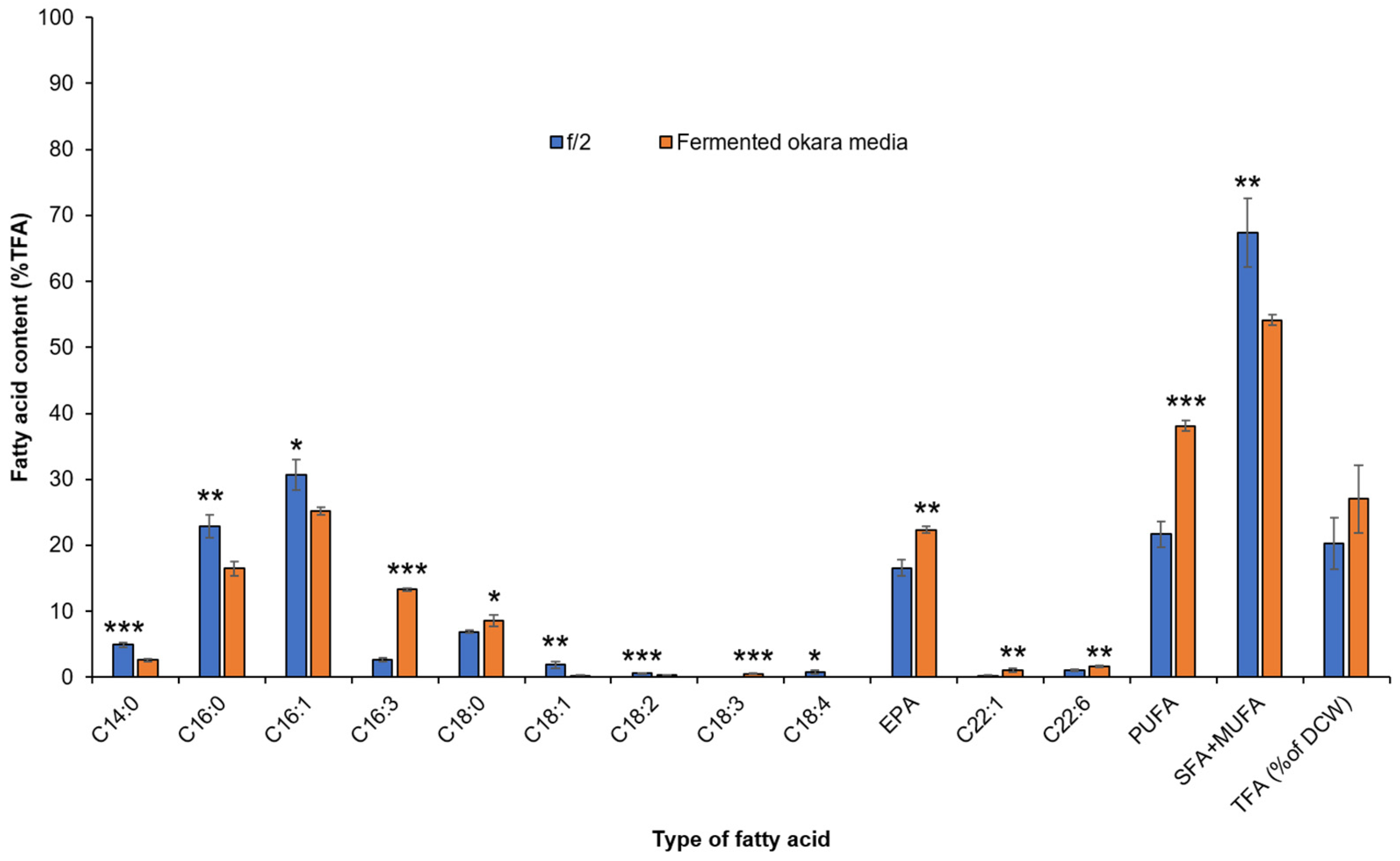
3.5. Fatty Acid Production from P. tricornutum Cells in Fermented Okara Culture
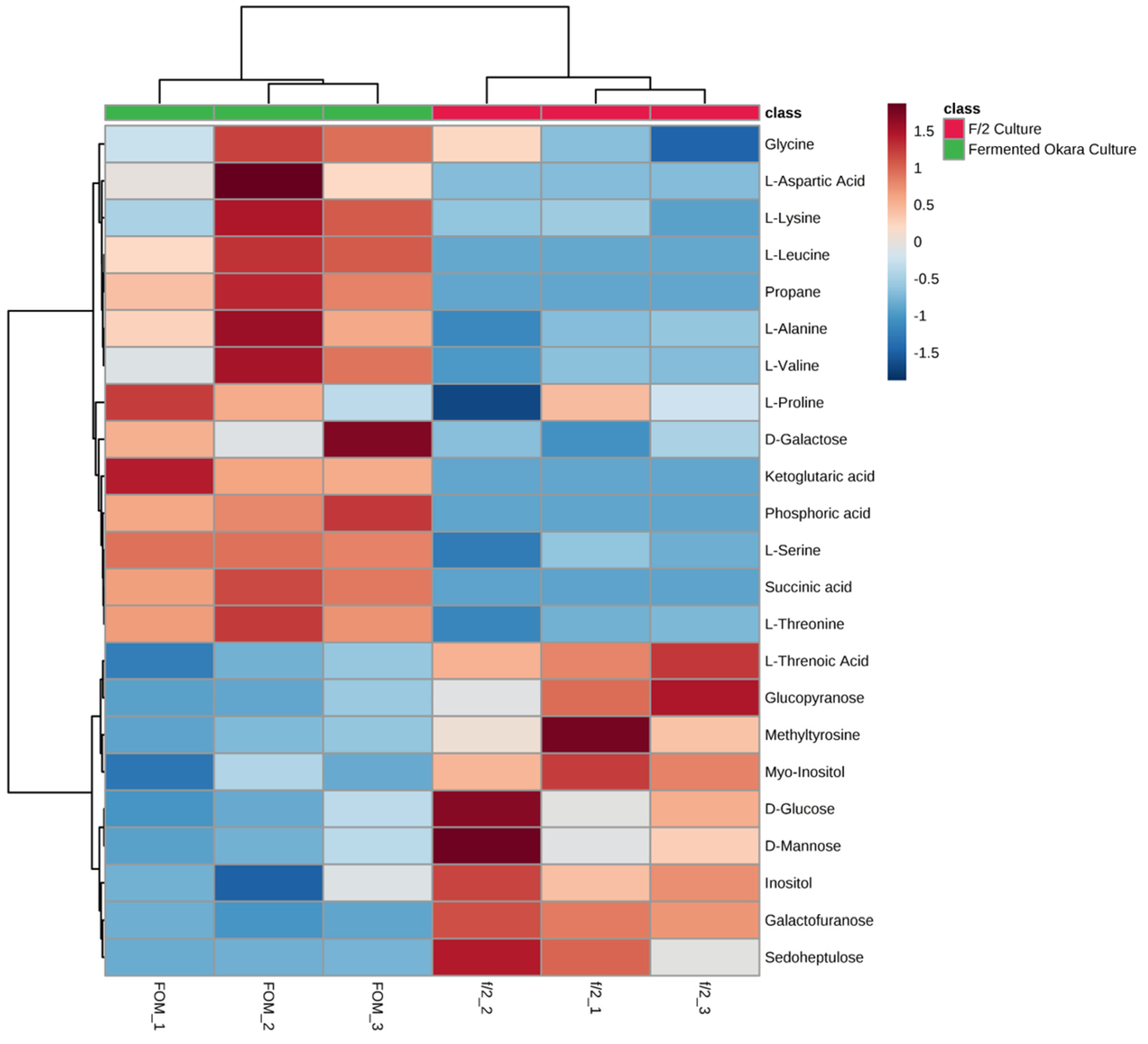
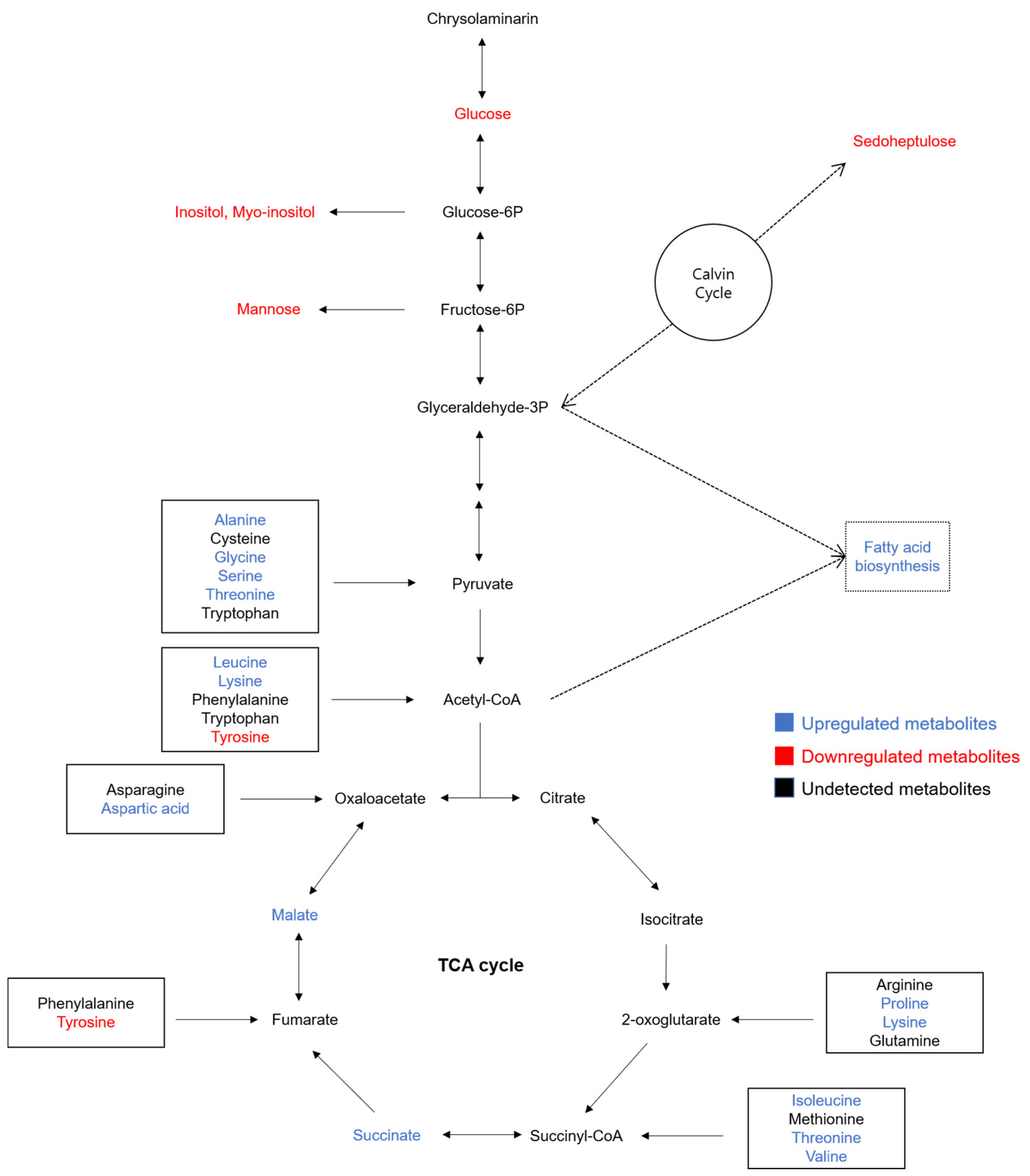
3.6. Metabolomic Profiling of the P. tricornutum Cells in Fermented Okara Culture
3.6.1. Remodeling of N Metabolism
3.6.2. Reprogramming of Carbon Allocation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, B.; Qiao, M.; Lu, F. Composition, Nutrition, and Utilization of Okara (Soybean Residue). Food Rev. Int. 2012, 28, 231–252. [Google Scholar] [CrossRef]
- van der Riet, W.B.; Wight, A.W.; Cilliers, J.J.L.; Datel, J.M. Food Chemical Investigation of Tofu and Its Byproduct Okara. Food Chem. 1989, 34, 193–202. [Google Scholar] [CrossRef]
- Colletti, A.; Attrovio, A.; Boffa, L.; Mantegna, S.; Cravotto, G. Valorisation of By-Products from Soybean (Glycine max (L.) Merr.) Processing. Molecules 2020, 25, 2129. [Google Scholar] [CrossRef]
- O’Toole, D.K. Characteristics and Use of Okara, the Soybean Residue from Soy Milk Production—A Review. J. Agric. Food Chem. 1999, 47, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Kasai, N.; Murata, A.; Inui, H.; Sakamoto, T.; Kahn, R.I. Enzymatic High Digestion of Soybean Milk Residue (Okara). J. Agric. Food Chem. 2004, 52, 5709–5716. [Google Scholar] [CrossRef]
- Yogo, T. Influence of Dried Okara-Tempeh on the Composition and Metabolites of Fecal Microbiota in Dogs. Int. J. Appl. Res. Vet. Med. 2011, 9, 176. [Google Scholar]
- Enari, T.-M.; Markkanen, P. Production of Cellulolytic Enzymes by Fungi. In Advances in Biochemical Engineering; Springer: Berlin/Heidelberg, Germany, 2005; Volume 5, pp. 1–24. [Google Scholar] [CrossRef]
- Lee, J.J.L.; Cooray, S.T.; Mark, R.; Chen, W.N. Effect of Sequential Twin Screw Extrusion and Fungal Pretreatment to Release Soluble Nutrients from Soybean Residue for Carotenoid Production. J. Sci. Food Agric. 2019, 99, 2646–2650. [Google Scholar] [CrossRef]
- Gupta, S.; Lee, J.J.L.; Chen, W.N. Analysis of Improved Nutritional Composition of Potential Functional Food (Okara) after Probiotic Solid-State Fermentation. J. Agric. Food Chem. 2018, 66, 5373–5381. [Google Scholar] [CrossRef]
- Chu, W. Biotechnological Applications of Microalgae. In Biotechnological Applications of Microalgae; CRC Press: Boca Raton, FL, USA, 2013; pp. 24–37. [Google Scholar] [CrossRef] [Green Version]
- Norsker, N.H.; Barbosa, M.J.; Vermuë, M.H.; Wijffels, R.H. Microalgal Production—A Close Look at the Economics. Biotechnol. Adv. 2011, 29, 24–27. [Google Scholar] [CrossRef]
- Bhatnagar, A.; Chinnasamy, S.; Singh, M.; Das, K.C. Renewable Biomass Production by Mixotrophic Algae in the Presence of Various Carbon Sources and Wastewaters. Appl. Energy 2011, 88, 3425–3431. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and Lipid Productivities of Chlorella Vulgaris under Autotrophic, Heterotrophic and Mixotrophic Growth Conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, W.; Hu, B.; Min, M.; Chen, P.; Ruan, R.R. Effect of Light Intensity on Algal Biomass Accumulation and Biodiesel Production for Mixotrophic Strains Chlorella Kessleri and Chlorella Protothecoide Cultivated in Highly Concentrated Municipal Wastewater. Biotechnol. Bioeng. 2012, 109, 2222–2229. [Google Scholar] [CrossRef] [PubMed]
- Cerón Garcí, M.C.; Fernández Sevilla, J.M.; Acién Fernández, F.G.; Molina Grima, E.; García Camacho, F. Mixotrophic Growth of Phaeodactylum tricornutum on Glycerol: Growth Rate and Fatty Acid Profile. J. Appl. Phycol. 2000, 12, 239–248. [Google Scholar] [CrossRef]
- Cerón García, M.C.; Sánchez Mirón, A.; Fernández Sevilla, J.M.; Molina Grima, E.; García Camacho, F. Mixotrophic Growth of the Microalga Phaeodactylum tricornutum: Influence of Different Nitrogen and Organic Carbon Sources on Productivity and Biomass Composition. Process. Biochem. 2005, 40, 297–305. [Google Scholar] [CrossRef]
- Wang, X.; Balamurugan, S.; Liu, S.F.; Zhang, M.M.; Yang, W.D.; Liu, J.S.; Li, H.Y.; Lin, C.S.K. Enhanced Polyunsaturated Fatty Acid Production Using Food Wastes and Biofuels Byproducts by an Evolved Strain of Phaeodactylum tricornutum. Bioresour. Technol. 2020, 296, 122351. [Google Scholar] [CrossRef]
- Pleissner, D.; Lam, W.C.; Sun, Z.; Lin, C.S.K. Food Waste as Nutrient Source in Heterotrophic Microalgae Cultivation. Bioresour. Technol. 2013, 137, 139–146. [Google Scholar] [CrossRef]
- Lopez-Huertas, E. Health Effects of Oleic Acid and Long Chain Omega-3 Fatty Acids (EPA and DHA) Enriched Milks. A Review of Intervention Studies. Pharmacol. Res. 2010, 61, 200–207. [Google Scholar] [CrossRef]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical Scavenging and Singlet Oxygen Quenching Activity of Marine Carotenoid Fucoxanthin and Its Metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from Edible Seaweed, Undaria Pinnatifida, Shows Antiobesity Effect through UCP1 Expression in White Adipose Tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef]
- Woo, M.N.; Jeon, S.M.; Kim, H.J.; Lee, M.K.; Shin, S.K.; Shin, Y.C.; Park, Y.B.; Choi, M.S. Fucoxanthin Supplementation Improves Plasma and Hepatic Lipid Metabolism and Blood Glucose Concentration in High-Fat Fed C57BL/6N Mice. Chem. Biol. Interact. 2010, 186, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Neumann, U.; Derwenskus, F.; Flister, V.F.; Schmid-Staiger, U.; Hirth, T.; Bischoff, S.C. Fucoxanthin, a Carotenoid Derived from Phaeodactylum tricornutum Exerts Antiproliferative and Antioxidant Activities in Vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, H.; Kamo, Y.; Machmudah, S.; Wahyudiono; Goto, M. Extraction of Fucoxanthin from Raw Macroalgae Excluding Drying and Cell Wall Disruption by Liquefied Dimethyl Ether. Mar. Drugs 2014, 12, 2383–2396. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Si, X.; Yuan, Z.; Xu, X.; Li, G. Isolation of Fucoxanthin from Edible Brown Algae by Microwave-Assisted Extraction Coupled with High-Speed Countercurrent Chromatography. J. Sep. Sci. 2012, 35, 2313–2317. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, C.H. A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Cerón García, M.C.; García Camacho, F.; Sánchez Mirón, A.; Fernández Sevilla, J.M.; Chisti, Y.; Molina Grima, E. Mixotrophic Production of Marine Microalga Phaeodactylum tricornutum on Various Carbon Sources. J. Microbiol. Biotechnol. 2006, 16, 689–694. [Google Scholar]
- Guillard Robert, R.L. Culture of Marine Invertebrates; Smith, W.L., Chanley, M.H., Eds.; Springer: New York, NY, USA, 1975. [Google Scholar]
- Uçkun Kiran, E.; Salakkam, A.; Trzcinski, A.P.; Bakir, U.; Webb, C. Enhancing the Value of Nitrogen from Rapeseed Meal for Microbial Oil Production. Enzym. Microb. Technol. 2012, 50, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Kirk, P.L. Kjeldahl Method for Total Nitrogen. Anal. Chem. 1950, 22, 354–358. [Google Scholar] [CrossRef]
- Kirk, S.R.; Ronald, S. Pearson’s Composition and Analysis of Foods; Longman Group Ltd.: Harlow, UK, 1991. [Google Scholar]
- Kim, J.; Lyu, X.M.; Lee, J.J.L.; Zhao, G.; Chin, S.F.; Yang, L.; Chen, W.N. Metabolomics Analysis of Pseudomonas Chlororaphis JK12 Algicidal Activity under Aerobic and Micro-Aerobic Culture Condition. AMB Express 2018, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Mok, W.K.; Tan, Y.X.; Lee, J.; Kim, J.; Chen, W.N. A Metabolomic Approach to Understand the Solid-State Fermentation of Okara Using Bacillus Subtilis WX-17 for Enhanced Nutritional Profile. AMB Express 2019, 9, 60. [Google Scholar] [CrossRef]
- Cooray, S.T.; Chen, W.N. Valorization of Brewer’s Spent Grain Using Fungi Solid-State Fermentation to Enhance Nutritional Value. J. Funct. Foods 2018, 42, 85–94. [Google Scholar] [CrossRef]
- Cooray, S.T.; Lee, J.J.L.; Chen, W.N. Evaluation of Brewers’ Spent Grain as a Novel Media for Yeast Growth. AMB Express 2017, 7, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yodsuwan, N.; Sawayama, S.; Sirisansaneeyakul, S. Effect of Nitrogen Concentration on Growth, Lipid Production and Fatty Acid Profiles of the Marine Diatom Phaeodactylum tricornutum. Agric. Nat. Resour. 2017, 51, 190–197. [Google Scholar] [CrossRef]
- Chauton, M.S.; Olsen, Y.; Vadstein, O. Biomass Production from the Microalga Phaeodactylum tricornutum: Nutrient Stress and Chemical Composition in Exponential Fed-Batch Cultures. Biomass Bioenergy 2013, 58, 87–94. [Google Scholar] [CrossRef]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An Investigation into the Effect of Culture Conditions on Fucoxanthin Production Using the Marine Microalgae Phaeodactylum tricornutum. Algal Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella Aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef]
- Guo, B.; Liu, B.; Yang, B.; Sun, P.; Lu, X.; Liu, J.; Chen, F. Screening of Diatom Strains and Characterization of Cyclotella Cryptica as a Potential Fucoxanthin Producer. Mar. Drugs 2016, 14, 125. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Loredo, A.; Benavides, J.; Rito-Palomares, M. Growth Kinetics and Fucoxanthin Production of Phaeodactylum Tricornutum and Isochrysis Galbana Cultures at Different Light and Agitation Conditions. J. Appl. Phycol. 2016, 28, 849–860. [Google Scholar] [CrossRef]
- Owens, T.G. Light-Harvesting Function in the Diatom Phaeodactylum tricornutum. Plant Physiol. 1986, 80, 739–746. [Google Scholar] [CrossRef] [Green Version]
- Molina Grima, E.; Sánchez Pérez, J.A.; García Camacho, F.; Fernández Sevilla, J.M.; Acién Fernández, F.G. Effect of Growth Rate on the Eicosapentaenoic Acid and Docosahexaenoic Acid Content of Isochrysis Galbana in Chemostat Culture. Appl. Microbiol. Biotechnol. 1994, 41, 23–27. [Google Scholar] [CrossRef]
- Hodgson, P.A.; Henderson, R.J.; Sargent, J.R.; Leftley, J.W. Patterns of Variation in the Lipid Class and Fatty Acid Composition of Nannochloropsis Oculata (Eustigmatophyceae) during Batch Culture—I. The Growth Cycle. J. Appl. Phycol. 1991, 3, 169–181. [Google Scholar] [CrossRef]
- Qiao, H.; Cong, C.; Sun, C.; Li, B.; Wang, J.; Zhang, L. Effect of Culture Conditions on Growth, Fatty Acid Composition and DHA/EPA Ratio of Phaeodactylum tricornutum. Aquaculture 2016, 452, 311–317. [Google Scholar] [CrossRef]
- Alipanah, L.; Rohloff, J.; Winge, P.; Bones, A.M.; Brembu, T. Whole-Cell Response to Nitrogen Deprivation in the Diatom Phaeodactylum tricornutum. J. Exp. Bot. 2015, 66, 6281–6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanova, V.; Fortunato, A.E.; Singh, D.; Bo, D.D.; Conte, M.; Obata, T.; Jouhet, J.; Fernie, A.R.; Marechal, E.; Falciatore, A.; et al. Investigating Mixotrophic Metabolism in the Model Diatom Phaeodactylum tricornutum. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160404. [Google Scholar] [CrossRef] [Green Version]
- Remmers, I.M.; D’Adamo, S.; Martens, D.E.; de Vos, R.C.H.; Mumm, R.; America, A.H.P.; Cordewener, J.H.G.; Bakker, L.V.; Peters, S.A.; Wijffels, R.H.; et al. Orchestration of Transcriptome, Proteome and Metabolome in the Diatom Phaeodactylum tricornutum during Nitrogen Limitation. Algal Res. 2018, 35, 33–49. [Google Scholar] [CrossRef]
- Granum, E.; Myklestad, S.M. A Simple Combined Method for Determination of β-1,3-Glucan and Cell Wall Polysaccharides in Diatoms. Hydrobiologia 2002, 477, 155–161. [Google Scholar] [CrossRef]
- Popko, J.; Herrfurth, C.; Feussner, K.; Ischebeck, T.; Iven, T.; Haslam, R.; Hamilton, M.; Sayanova, O.; Napier, J.; Khozin-Goldberg, I.; et al. Metabolome Analysis Reveals Betaine Lipids as Major Source for Triglyceride Formation, and the Accumulation of Sedoheptulose during Nitrogen-Starvation of Phaeodactylum tricornutum. PLoS ONE 2016, 11, e0164673. [Google Scholar] [CrossRef] [Green Version]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum Genome Reveals the Evolutionary History of Diatom Genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, A.; Maheswari, U.; Dorrell, R.G.; Vieira, F.R.J.; Maumus, F.; Kustka, A.; McCarthy, J.; Allen, A.E.; Kersey, P.; Bowler, C.; et al. Integrative Analysis of Large Scale Transcriptome Data Draws a Comprehensive Landscape of Phaeodactylum tricornutum Genome and Evolutionary Origin of Diatoms. Sci. Rep. 2018, 8, 4834. [Google Scholar] [CrossRef] [Green Version]
- Eilers, U.; Bikoulis, A.; Breitenbach, J.; Büchel, C.; Sandmann, G. Limitations in the Biosynthesis of Fucoxanthin as Targets for Genetic Engineering in Phaeodactylum tricornutum. J. Appl. Phycol. 2016, 28, 123–129. [Google Scholar] [CrossRef]
- Zaslavskaia, L.A.; Lippmeier, J.C.; Shih, C.; Ehrhardt, D.; Grossman, A.R.; Apt, K.E. Trophic Conversion of an Obligate Photoautotrophic Organism through Metabolic Engineering. Science 2001, 292, 2073–2075. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined Production of Fucoxanthin and EPA from Two Diatom Strains Phaeodactylum Tricornutum and Cylindrotheca Fusiformis Cultures. Bioprocess Biosyst. Eng. 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chen, A.; Zhang, W.; Li, A.; Zhang, C. Co-Production of Lipids, Eicosapentaenoic Acid, Fucoxanthin, and Chrysolaminarin by Phaeodactylum Tricornutum Cultured in a Flat-Plate Photobioreactor under Varying Nitrogen Conditions. J. Ocean. Univ. China 2017, 16, 916–924. [Google Scholar] [CrossRef]
- Yang, R.; Wei, D. Improving Fucoxanthin Production in Mixotrophic Culture of Marine Diatom Phaeodactylum Tricornutum by LED Light Shift and Nitrogen Supplementation. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Li, T.; Wang, G.; Dai, S.; He, H.; Xiang, W. A Comparative Analysis of Fatty Acid Composition and Fucoxanthin Content in Six Phaeodactylum Tricornutum Strains from Diff Erent Origins. Chin. J. Oceanol. Limnol. 2016, 34, 391–398. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Fermented Okara Media | F/2 a | Fold Change |
|---|---|---|---|
| Carbohydrates (mg L−1) | 300 | 0 | N.A. |
| Nitrogen (mg L−1) | 150 | 12.7 | 12 |
| Phosphorus (mg L−1) | 14 | 1.1 | 12 |
| Carbon (wt.%) | 2.09 | 0.15 | 14 |
| Parameters | Fermented Okara Media | F/2 Media | Fold Change |
|---|---|---|---|
| Chlorophyll a content (mg g−1) | 76.5 ± 5.6 | 23.2 ± 6.2 | 3 |
| Fucoxanthin content (mg g−1) | 15.3 ± 0.6 | 7.3 ± 1.8 | 2 |
| Fucoxanthin yield (mg L−1) | 8.2 ± 0.3 | 1.7 ± 0.3 | 5 |
| Fatty Acid Yield (mg L−1) | Growth Media | ||
|---|---|---|---|
| Fermented Okara Media | F/2 Media | Fold Change | |
| C14:0 | 3.66 ± 0.46 | 3.32 ± 0.19 | 1.1 |
| C16:0 | 23.32 ± 5.78 | 15.62 ± 2.24 | 1.5 |
| C16:1 | 35.41 ± 6.27 | 20.93 ± 3.24 | 1.7 |
| C16:3 | 18.67 ± 3.32 | 1.83 ± 0.11 | 10.2 |
| C18:0 | 12.19 ± 3.40 | 4.69 ± 0.37 | 2.6 |
| C18:1 | 0.28 ± 0.24 | 1.28 ± 0.44 | 0.2 |
| C18:2 | 0.45 ± 0.05 | 0.39 ± 0.04 | 1.2 |
| C18:3 | 0.70 ± 0.05 | N.D. | - |
| C18:4 | N.D. | 0.53 ± 0.25 | - |
| C20:5 | 31.36 ± 5.38 | 11.26 ± 0.88 | 2.8 |
| C22:1 | 1.45 ± 0.10 | 0.12 ± 0.12 | 12.1 |
| C22:6 | 2.30 ± 0.49 | 0.70 ± 0.05 | 3.3 |
| SFA | 39.17 ± 9.64 | 23.63 ± 2.76 | 1.7 |
| MUFA | 37.14 ± 6.11 | 22.33 ± 3.78 | 1.7 |
| PUFA | 52.78 ± 9.19 | 14.71 ± 1.05 | 3.6 |
| SFA+MUFA | 76.31 ± 15.59 | 45.96 ± 6.55 | 1.7 |
| TFAC | 140.69 ± 27.00 | 68.09 ± 6.71 | 2.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Lee, J.; Voo, A.Y.H.; Tan, Y.X.; Mok, W.K.; Li, A.Z.; Chen, W.N. Okara Waste as a Substrate for the Microalgae Phaeodactylum tricornutum Enhances the Production of Algal Biomass, Fucoxanthin, and Polyunsaturated Fatty Acids. Fermentation 2023, 9, 31. https://doi.org/10.3390/fermentation9010031
Kim J, Lee J, Voo AYH, Tan YX, Mok WK, Li AZ, Chen WN. Okara Waste as a Substrate for the Microalgae Phaeodactylum tricornutum Enhances the Production of Algal Biomass, Fucoxanthin, and Polyunsaturated Fatty Acids. Fermentation. 2023; 9(1):31. https://doi.org/10.3390/fermentation9010031
Chicago/Turabian StyleKim, Jaejung, Jaslyn Lee, Amanda Ying Hui Voo, Yong Xing Tan, Wai Kit Mok, Aaron Zongwei Li, and Wei Ning Chen. 2023. "Okara Waste as a Substrate for the Microalgae Phaeodactylum tricornutum Enhances the Production of Algal Biomass, Fucoxanthin, and Polyunsaturated Fatty Acids" Fermentation 9, no. 1: 31. https://doi.org/10.3390/fermentation9010031