
Effects of Dietary Capsaicin and Yucca schidigera Extracts as Feed Additives on Rumen Fermentation and Microflora of Beef Cattle Fed with a Moderate-Energy Diet


Abstract
:1. Introduction
2. Materials and Methods
2.1. Diets, Animals, and Experimental Design
2.2. Feeding Management
2.3. Ruminal Fluid Sample Collection and Measurement
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Statistical Analysis
3. Results
3.1. Feed Intake and Rumen Fermentation Parameters
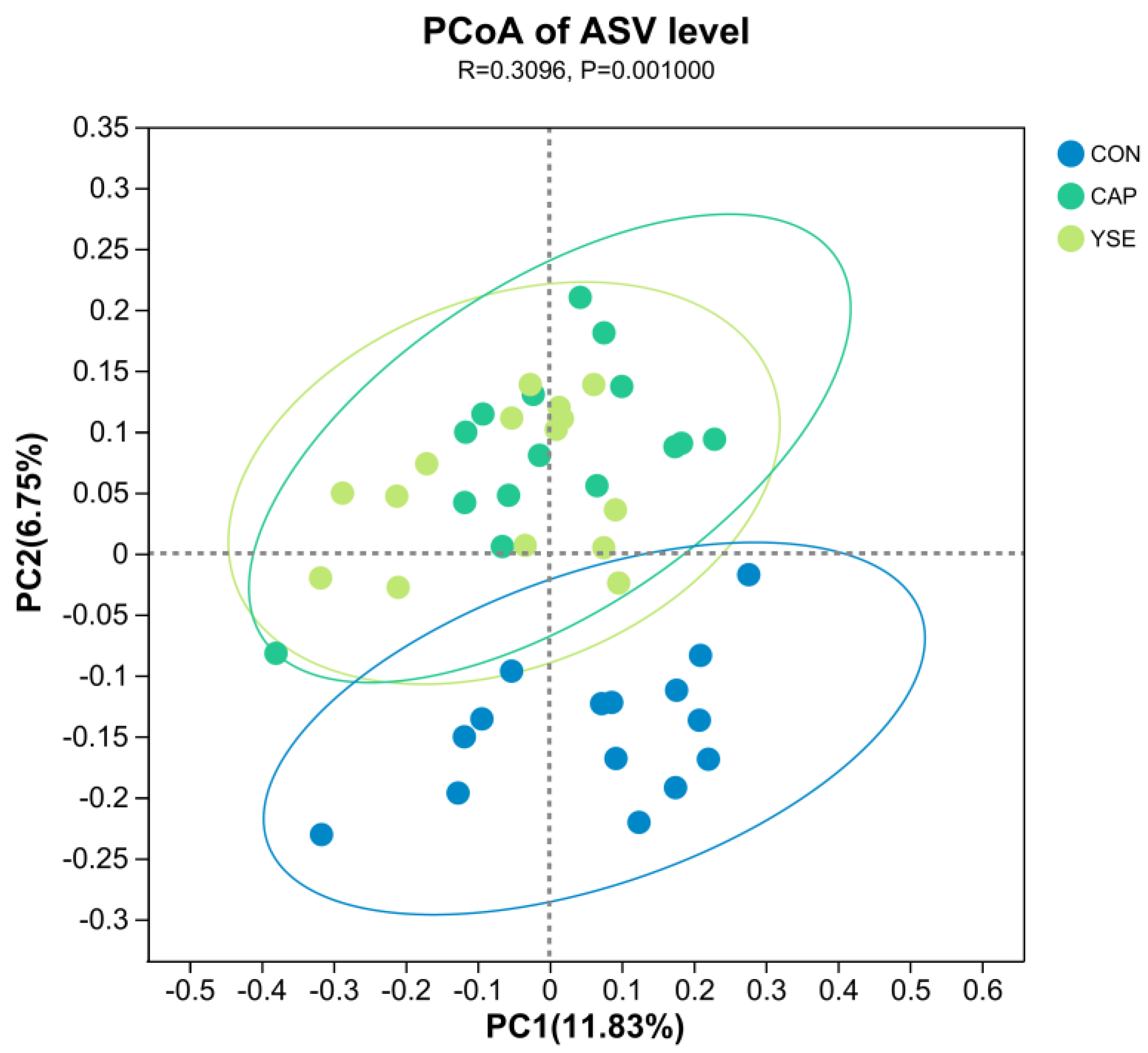
3.2. Bacterial Abundance and Diversity in the Rumen
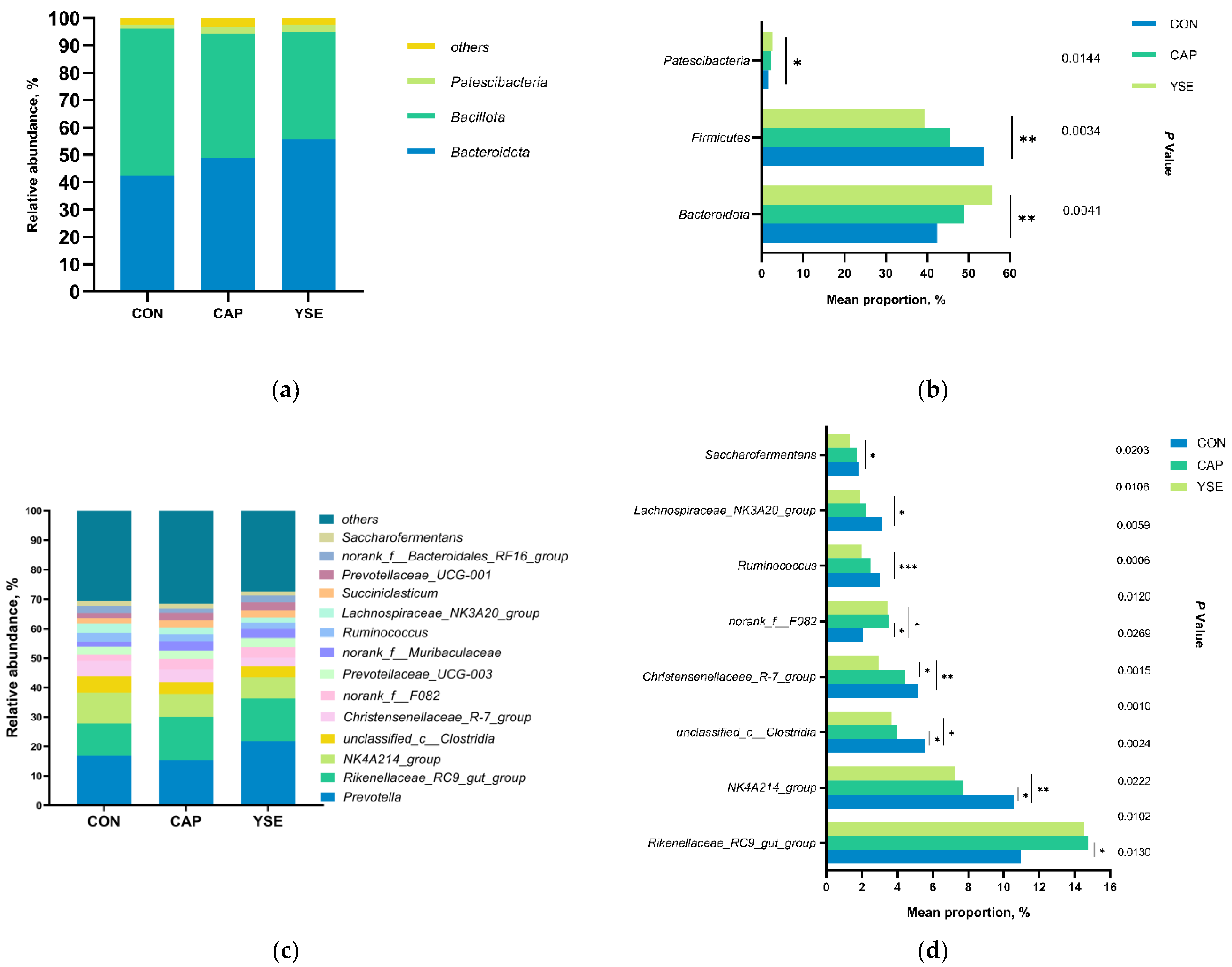
3.3. Rumen Bacterial Community Composition and Species Differences
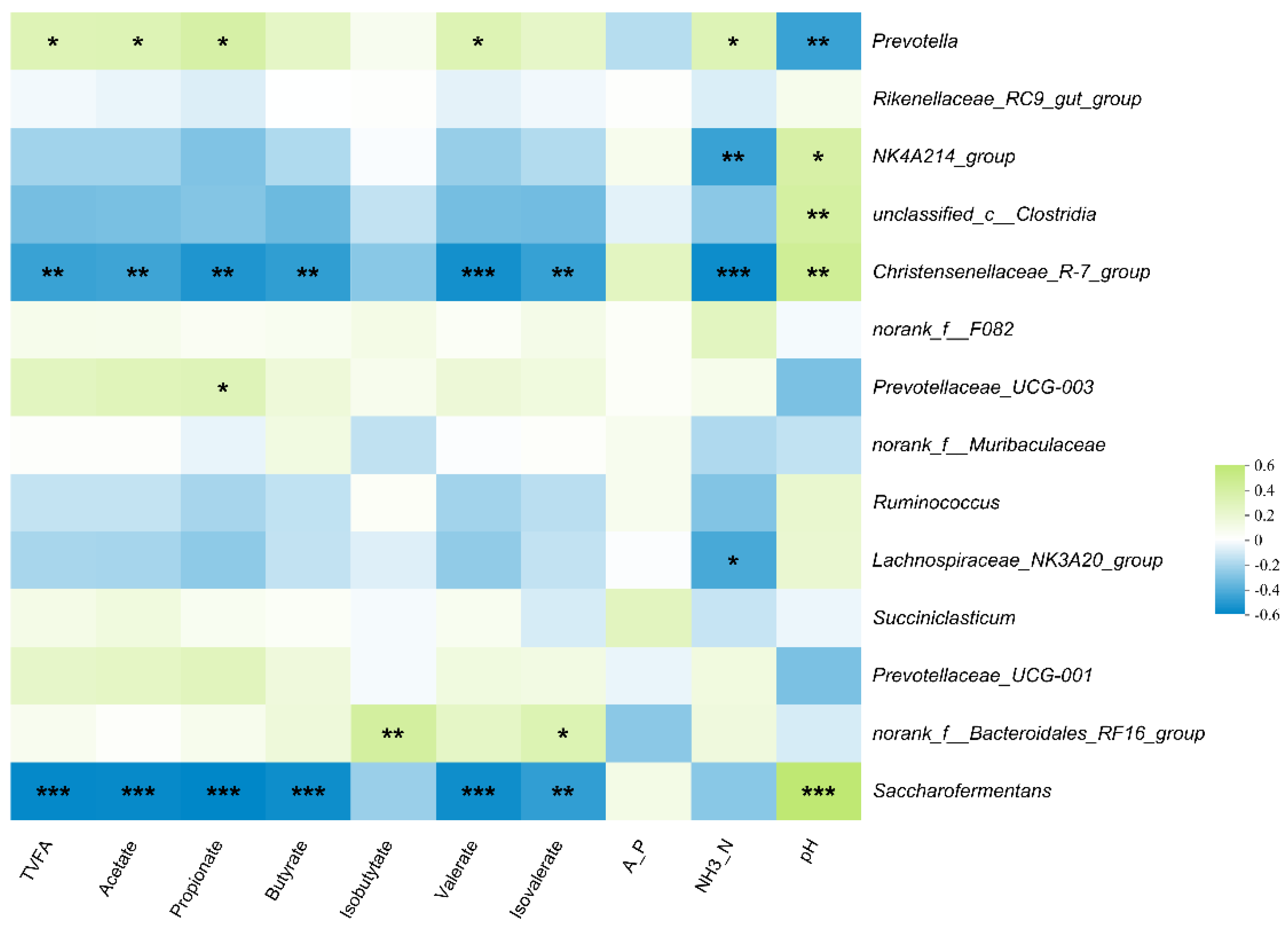
3.4. Correlation between Rumen Bacteria and Rumen Fermentation Parameters
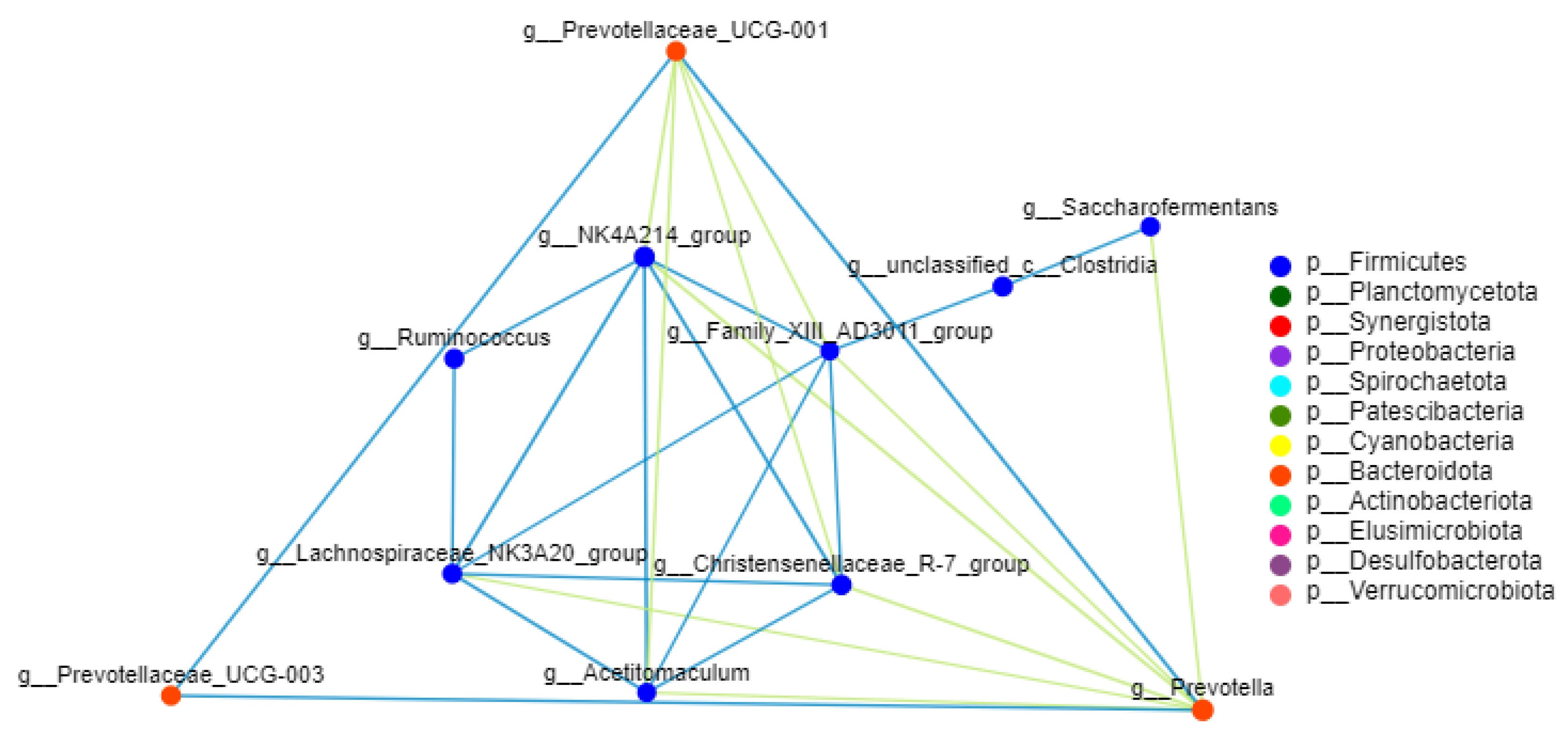
3.5. Correlation of Rumen Bacteria at the Genus Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doyle, N.; Mbandlwa, P.; Kelly, W.J.; Attwood, G.; Li, Y.; Ross, R.P.; Stanton, C.; Leahy, S. Use of Lactic Acid Bacteria to Reduce Methane Production in Ruminants, a Critical Review. Front. Microbiol. 2019, 10, 2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruninger, R.J.; Ribeiro, G.O.; Cameron, A.; McAllister, T.A. Invited review: Application of meta-omics to understand the dynamic nature of the rumen microbiome and how it responds to diet in ruminants. Animal 2019, 13, 1843–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, J.E.; Kingston-Smith, A.H.; Jimenez, H.R.; Huws, S.A.; Skøt, K.P.; Griffith, G.W.; McEwan, N.R.; Theodorou, M.K. Dynamics of initial colonization of nonconserved perennial ryegrass by anaerobic fungi in the bovine rumen. FEMS Microbiol. Ecol. 2008, 66, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Pan, J.; Kobayashi, Y.; Tanaka, K. Kinetics of in sacco fiber-attachment of representative ruminal cellulolytic bacteria monitored by competitive PCR. J. Dairy Sci. 2003, 86, 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.A.; Bae, H.D.; Jones, G.A.; Cheng, K.J. Microbial Attachment and Feed Digestion in the Rumen. J. Anim. Sci. 1994, 72, 3004–3018. [Google Scholar] [CrossRef] [Green Version]
- Flachowsky, G.; Lebzien, P. Effects of phytogenic substances on rumen fermentation and methane emissions: A proposal for a research process. Anim. Feed Sci. Technol. 2012, 176, 70–77. [Google Scholar] [CrossRef]
- Arowolo, M.A.; He, J. Use of probiotics and botanical extracts to improve ruminant production in the tropics: A review. Anim. Nutr. 2018, 4, 241–249. [Google Scholar] [CrossRef]
- Purba, R.A.P.; Paengkoum, S.; Yuangklang, C.; Paengkoum, P.; Salem, A.Z.M.; Boo, L.J. Mammary gene expressions and oxidative indicators in ruminal fluid, blood, milk, and mammary tissue of dairy goats fed a total mixed ration containing piper meal (Piper betle L.). Ital. J. Anim. Sci. 2022, 21, 129–141. [Google Scholar] [CrossRef]
- Aihaiti, K.; Li, J.; Yaermaimaiti, S.; Liu, L.; Xin, X.; Aisa, H.A. Non-volatile compounds of Hyssopus cuspidatus Boriss and their antioxidant and antimicrobial activities. Food Chem. 2022, 374, 131638. [Google Scholar] [CrossRef]
- Liu, J.-H.; Li, C.; Cao, L.; Zhang, C.-H.; Zhang, Z.-H. Cucurbitacin B regulates lung cancer cell proliferation and apoptosis via inhibiting the IL-6/STAT3 pathway through the lncRNA XIST/miR-let-7c axis. Pharm. Biol. 2022, 60, 154–162. [Google Scholar] [CrossRef]
- Jiao, P.; Hu, G.; Liang, G.; Chen, M.; An, N.; Wang, Z.; Liu, H.; Xing, H.; Xie, X. Dietary supplementation with Macleaya cordata extract inclusion affects growth performance, rumen fermentation, bacterial communities, and immune responses of weaned lambs. Anim. Feed Sci. Technol. 2021, 282, 115127. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Goci, E.; Haloci, E.; Di Stefano, A.; Chiavaroli, A.; Angelini, P.; Miha, A.; Cacciatore, I.; Marinelli, L. Evaluation of In Vitro Capsaicin Release and Antimicrobial Properties of Topical Pharmaceutical Formulation. Biomolecules 2021, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Dong, B.; Friesen, M.; Liu, S.; Zhu, C.; Yang, C. Capsaicin Attenuates Lipopolysaccharide-Induced Inflammation and Barrier Dysfunction in Intestinal Porcine Epithelial Cell Line-J2. Front. Physiol. 2021, 12, 715469. [Google Scholar] [CrossRef]
- Wang, Y.; McAllister, T.A.; Yanke, L.J.; Cheeke, P.R. Effect of steroidal saponin from Yucca schidigera extract on ruminal microbes. J. Appl. Microbiol. 2000, 88, 887–896. [Google Scholar] [CrossRef]
- Zuniga-Serrano, A.; Barrios-Garcia, H.B.; Anderson, R.C.; Hume, M.E.; Ruiz-Albarran, M.; Bautista-Martinez, Y.; Sanchez-Guerra, N.A.; Vazquez-Villanueva, J.; Infante-Rodriguez, F.; Salinas-Chavira, J. Antimicrobial and Digestive Effects of Yucca schidigera Extracts Related to Production and Environment Implications of Ruminant and Non-Ruminant Animals: A Review. Agriculture 2022, 12, 1198. [Google Scholar] [CrossRef]
- Liu, C.L.; Li, Z.Q. Effect of Levels of Yucca Schidigera Extract on Ruminal Fermentation Parameters, Digestibility of Nutrients and Growth Performance in Sheep. Adv. Mater. Res. 2011, 343–344, 655–660. [Google Scholar] [CrossRef]
- Wu, Z.; Sadik, M.; Sleiman, F.T.; Simas, J.M.; Pessarakli, M.; Huber, J.T. Influence of Yucca Extract on Ruminal Metabolism in Cows. J. Anim. Sci. 1994, 72, 1038–1042. [Google Scholar] [CrossRef]
- Rodriguez-Prado, M.; Ferret, A.; Zwieten, J.; Gonzalez, L.; Bravo, D.; Calsamiglia, S. Effects of dietary addition of capsicum extract on intake, water consumption, and rumen fermentation of fattening heifers fed a high-concentrate diet. J. Anim. Sci. 2012, 90, 1879–1884. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle, 8th ed.; The National Academies Press: Washington, DC, USA, 2016; p. 494. [Google Scholar]
- Erwin, E.S.; Marco, G.J.; Emery, E.M. Volatile Fatty Acid Analyses of Blood and Rumeen Fluid by Gas Chromatography. J. Dairy Sci. 1961, 44, 1768–1771. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated Simultaneous Determination of Ammonia and Total Amino-Acids in Ruminal Fluid and In Vitro Media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Meng, Q. Research advances in prediction models of dry matter intake in ruminants. Chin. J. Anim. Nutr. 2013, 25, 248–255. [Google Scholar]
- Noziere, P.; Glasser, F.; Sauvant, D. In vivo production and molar percentages of volatile fatty acids in the rumen: A quantitative review by an empirical approach. Animal 2011, 5, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westphalen, M.F.; Carvalho, P.H.V.; Oh, J.; Hristov, A.N.; Staniar, W.B.; Felix, T.L. Effects of feeding rumen-protected Capsicum oleoresin on growth performance, health status, and total tract digestibility of growing beef cattle. Anim. Feed Sci. Technol. 2021, 271, 114778. [Google Scholar] [CrossRef]
- Rett, B.; Cooke, R.F.; Brandao, A.P.; Ferreira, V.S.M.; Colombo, E.A.; Wiegand, J.B.; Pohler, K.G.; Rincker, M.J.; Schubach, K.M. Supplementing Yucca schidigera extract to mitigate frothy bloat in beef cattle receiving a high-concentrate diet. J. Anim. Sci. 2020, 98, skaa355. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Sen, S.; Blummel, M.; Becker, K. Effects of fractions containing saponins from Yucca schidigera, Quillaja saponaria, and Acacia auriculoformis on rumen fermentation. J. Agric. Food Chem. 1998, 46, 4324–4328. [Google Scholar] [CrossRef]
- Holtshausen, L.; Chaves, A.V.; Beauchemin, K.A.; McGinn, S.M.; McAllister, T.A.; Cheeke, P.R.; Benchaar, C. Feeding saponin-containing Yucca schidigera and Quillaja saponaria to decrease enteric methane production in dairy cows. J. Dairy Sci. 2009, 92, 2809–2821. [Google Scholar] [CrossRef] [PubMed]
- Chalermglin, P.; Hong, S.H.; Choi, Y.J.; Hwang, I.H.; Lee, S.S.; Ha, J.K. Effect of Yucca extract on in vitro fermentation by mixed ruminal microorganism. J. Anim. Sci. Technol. 2001, 43, 707–720. [Google Scholar]
- Wallace, R.J.; Arthaud, L.; Newbold, C.J. Influence of Yucca-Shidigera Extract on Ruminal Ammonia Concentrations and Ruminal Microorganisms. Appl. Environ. Microbiol. 1994, 60, 1762–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Raheem, S.M.; Farghaly, M.M.; Hassan, E.H. Effect of dietary supplementation with Yucca schidigera powder on nutrient digestibility, rumen fermentation, rumenal enzyme activities and growth performance of buffalo calves. Biol. Rhythm. Res. 2022, 53, 854–866. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Cheeke, P.R. Actual and potential applications of Yucca schidigera and Quillaja saponaria saponins in human and animal nutrition. In Saponins in Food, Feedstuffs and Medicinal Plants; Springer: Dordrecht, The Netherlands, 2000; Volume 45, pp. 241–254. [Google Scholar]
- Benchaar, C.; McAllister, T.A.; Chouinard, P.Y. Digestion, Ruminal Fermentation, Ciliate Protozoal Populations, and Milk Production from Dairy Cows Fed Cinnamaldehyde, Quebracho Condensed Tannin, or Yucca schidigera Saponin Extracts. J. Dairy Sci. 2008, 91, 4765–4777. [Google Scholar] [CrossRef] [Green Version]
- Eryavuz, A.; Dehority, B.A. Effect of Yucca schidigera extract on the concentration of rumen microorganisms in sheep. Anim. Feed. Sci. Technol. 2004, 117, 215–222. [Google Scholar] [CrossRef]
- Gumus, R.; Imik, H. Effects of Yucca schidigera powder added to lamb ration on rumen total number of protozoa and pH. Ataturk Univ. Vet. Bilim. Derg. 2016, 11, 159–165. [Google Scholar]
- Liu, Y.; Ma, T.; Chen, D.; Zhang, N.; Si, B.; Deng, K.; Tu, Y.; Diao, Q. Effects of Tea Saponin Supplementation on Nutrient Digestibility, Methanogenesis, and Ruminal Microbial Flora in Dorper Crossbred Ewe. Animals 2019, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Goodall, S.R.; Reyes, A.; Guimaraes, O.; Belk, K.; Engle, T.E. Effects of polyphenols (mixed tannins) and saponin (Yucca schidigera), with or without a direct fed microbial (DFM) on in vitro rumen fermentation. J. Anim. Sci. 2020, 98, 427. [Google Scholar] [CrossRef]
- Chaput, J.P.; Thivierge, M.C.; Tremblay, A. Propionate: Hypophagic Effects Observed in Animal Models Might be Transposed to the Human Obesity Management. Curr. Nutr. Food Sci. 2006, 2, 375–379. [Google Scholar] [CrossRef]
- Wu, S.; Cui, Z.; Chen, X.; Zheng, L.; Ren, H.; Wang, D.; Yao, J. Diet-ruminal microbiome-host crosstalk contributes to differential effects of calf starter and alfalfa hay on rumen epithelial development and pancreatic alpha-amylase activity in yak calves. J. Dairy Sci. 2021, 104, 4326–4340. [Google Scholar] [CrossRef] [PubMed]
- Allison, M.J. Biosynthesis of Amino Acids by Ruminal Microorganisms. J. Anim. Sci. 1969, 29, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Andries, J.I.; Buysse, F.X.; Debrabander, D.L.; Cottyn, B.G. Isoacids in Ruminant Nutrition—Their Role in Ruminal and Intermediary Metabolism and Possible Influences on Performances—A Review. Anim. Feed Sci. Technol. 1987, 18, 169–180. [Google Scholar] [CrossRef]
- Mohammed, R.; Brink, G.E.; Stevenson, D.M.; Neumann, A.P.; Beauchemin, K.A.; Suen, G.; Weimer, P.J. Bacterial communities in the rumen of Holstein heifers differ when fed orchardgrass as pasture vs. hay. Front. Microbiol. 2014, 5, 689. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Geng, S.; Liu, M.; Zhao, L.; Zhang, J.; Huang, S.; Ma, Q. Effects of Different Methionine Levels in Low Protein Diets on Production Performance, Reproductive System, Metabolism, and Gut Microbiota in Laying Hens. Front. Nutr. 2021, 8, 739676. [Google Scholar] [CrossRef]
- Monteiro, H.F.; Lelis, A.L.J.; Fan, P.; Agustinho, B.C.; Lobo, R.R.; Arce-Cordero, J.A.; Dai, X.; Jeong, K.C.; Faciola, A.P. Effects of lactic acid-producing bacteria as direct-fed microbials on the ruminal microbiome. J. Dairy Sci. 2022, 105, 2242–2255. [Google Scholar] [CrossRef]
- Moraïs, S.; Mizrahi, I. The Road Not Taken: The Rumen Microbiome, Functional Groups, and Community States. Trends Microbiol. 2019, 27, 538–549. [Google Scholar] [CrossRef]
- Purushe, J.; Fouts, D.E.; Morrison, M.; White, B.A.; Mackie, R.I.; Coutinho, P.M.; Henrissat, B.; Nelson, K.E. Comparative Genome Analysis of Prevotella ruminicola and Prevotella bryantii: Insights into Their Environmental Niche. Microb. Ecol. 2010, 60, 721–729. [Google Scholar] [CrossRef]
- Thomas, F.; Hehemann, J.-H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut Bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [Green Version]
- Ozbayram, E.G.; Kleinsteuber, S.; Nikolausz, M.; Ince, B.; Ince, O. Enrichment of lignocellulose-degrading microbial communities from natural and engineered methanogenic environments. Appl. Microbiol. Biotechnol. 2018, 102, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Lopes, D.R.G.; de Souza Duarte, M.; La Reau, A.J.; Chaves, I.Z.; de Oliveira Mendes, T.A.; Detmann, E.; Bento, C.B.P.; Mercadante, M.E.Z.; Bonilha, S.F.M.; Suen, G.; et al. Assessing the relationship between the rumen microbiota and feed efficiency in Nellore steers. J. Anim. Sci. Biotechnol. 2021, 12, 79. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.; Chai, S.; Yang, Y.; Wang, X.; Liu, S.; Wang, S. Dietary forage to concentrate ratios impact on yak ruminal microbiota and metabolites. Front. Microbiol. 2022, 13, 964564. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, H.; Deng, J.; Fan, D. Ginsenoside Rk3 Ameliorates Obesity-Induced Colitis by Regulating of Intestinal Flora and the TLR4/NF-kappa B Signaling Pathway in C57BL/6 Mice. J. Agric. Food Chem. 2021, 69, 3082–3093. [Google Scholar] [CrossRef]
- Shu, X.; Chen, R.; Yang, M.; Xu, J.; Gao, R.; Hu, Y.; He, X.; Zhao, C. Gynostemma pentaphyllum and Gypenoside-IV Ameliorate Metabolic Disorder and Gut Microbiota in Diet-Induced-Obese Mice. Plant Foods Hum. Nutr. 2022, 77, 367–372. [Google Scholar] [CrossRef]
- Xu, J.; Li, T.; Xia, X.; Fu, C.; Wang, X.; Zhao, Y. Dietary Ginsenoside T19 Supplementation Regulates Glucose and Lipid Metabolism via AMPK and PI3K Pathways and Its Effect on Intestinal Microbiota. J. Agric. Food Chem. 2020, 68, 14452–14462. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.-A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Lemos, L.N.; Medeiros, J.D.; Dini-Andreote, F.; Fernandes, G.R.; Varani, A.M.; Oliveira, G.; Pylro, V.S. Genomic signatures and co-occurrence patterns of the ultra-small Saccharimonadia (phylum CPR/Patescibacteria) suggest a symbiotic lifestyle. Mol. Ecol. 2019, 28, 4259–4271. [Google Scholar] [CrossRef]
- Castelle, C.J.; Banfield, J.F. Major New Microbial Groups Expand Diversity and Alter Our Understanding of the Tree of Life. Cell 2018, 172, 1181–1197. [Google Scholar] [CrossRef] [Green Version]
- Holman, D.B.; Gzyl, K.E. A meta-analysis of the bovine gastrointestinal tract microbiota. FEMS Microbiol. Ecol. 2019, 95, fiz072. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, R.; Wu, T.; Yang, Y.; He, Z.; Ma, Z.; Tan, Z.; Lin, B.; Wang, M. Comparisons of Corn Stover Silages after Fresh- or Ripe-Corn Harvested: Effects on Digestibility and Rumen Fermentation in Growing Beef Cattle. Animals 2022, 12, 1248. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Guo, L.; Chang, X.; Liu, K.; Tang, W.; Zheng, N.; Zhao, S.; Zhang, Y.; Wang, J. Effect of Whole or Ground Flaxseed Supplementation on Fatty Acid Profile, Fermentation, and Bacterial Composition in Rumen of Dairy Cows. Front. Microbiol. 2021, 12, 760528. [Google Scholar] [CrossRef] [PubMed]
- Tomazetto, G.; Hahnke, S.; Wibberg, D.; Puhler, A.; Klocke, M.; Schluter, A. Proteiniphilum saccharofermentans str. M3/6T isolated from a laboratory biogas reactor is versatile in polysaccharide and oligopeptide utilization as deduced from genome-based metabolic reconstructions. Biotechnol. Rep. 2018, 18, e00254. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Hao, Z.; Shi, H.; Li, T.; Wang, H.; Li, Q.; Zhang, Y.; Ma, Y. Metagenomic Analysis Revealed Differences in Composition and Function between Liquid-Associated and Solid-Associated Microorganisms of Sheep Rumen. Front. Microbiol. 2022, 13, 851567. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zhang, D.; Qiu, X.; Zhao, K.; Zhang, S.; Liu, C.; Lu, L.; Cui, Y.; Shi, C.; Chen, Z.; et al. 2-Hydroxy-4-(Methylthio) Butanoic Acid Isopropyl Ester Supplementation Altered Ruminal and Cecal Bacterial Composition and Improved Growth Performance of Finishing Beef Cattle. Front. Nutr. 2022, 9, 833881. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef] [Green Version]
- Evans, N.J.; Brown, J.M.; Murray, R.D.; Getty, B.; Birtles, R.J.; Hart, C.A.; Carter, S.D. Characterization of Novel Bovine Gastrointestinal Tract Treponema Isolates and Comparison with Bovine Digital Dermatitis Treponemes. Appl. Environ. Microbiol. 2011, 77, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shen, Q.; Zhong, S.; Chen, Y.; Yang, Y. Comparison of Rumen Microbiota and Serum Biochemical Indices in White Cashmere Goats Fed Ensiled or Sun-Dried Mulberry Leaves. Microorganisms 2020, 8, 981. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Y.; Cui, Z.; Yang, Y.; An, X.; Qi, J. Fermented Wheat Bran Polysaccharides Intervention Alters Rumen Bacterial Community and Promotes Rumen Development and Growth Performance in Lambs. Front. Vet. Sci. 2022, 9, 841406. [Google Scholar] [CrossRef]
- Wang, Y.; Nan, X.; Zhao, Y.; Wang, Y.; Jiang, L.; Xiong, B. Ruminal Degradation of Rumen-Protected Glucose Influences the Ruminal Microbiota and Metabolites in Early-Lactation Dairy Cows. Appl. Environ. Microbiol. 2021, 87, e01908–e01920. [Google Scholar] [CrossRef]
- Newbold, C.J.; de la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valera-Vera, E.A.; Reigada, C.; Saye, M.; Digirolamo, F.A.; Galceran, F.; Miranda, M.R.; Pereira, C.A. Effect of capsaicin on the protozoan parasite Trypanosoma cruzi. FEMS Microbiol. Lett. 2020, 367, fnaa194. [Google Scholar] [CrossRef]
- Saye, M.; Gauna, L.; Valera-Vera, E.; Reigada, C.; Miranda, M.R.; Pereira, C.A. Crystal violet structural analogues identified by in silico drug repositioning present anti-Trypanosoma cruzi activity through inhibition of proline transporter TcAAAP069. PLoS Negl. Trop. Dis. 2020, 14, 7481. [Google Scholar] [CrossRef] [PubMed]
- Valera Vera, E.A.; Saye, M.; Reigada, C.; Damasceno, F.S.; Silber, A.M.; Miranda, M.R.; Pereira, C.A. Resveratrol inhibits Trypanosoma cruzi arginine kinase and exerts a trypanocidal activity. Int. J. Biol. Macromol. 2016, 87, 498–503. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | 1–59 Days | 60–90 Days |
|---|---|---|
| Ingredient (% of DM) | ||
| Ground corn | 31.90 | 37.70 |
| Soybean meal | 9.35 | 11.05 |
| Jujube powder | 8.25 | 9.75 |
| Whole-plant corn silage | 24.75 | 19.25 |
| Corn stalker | 20.25 | 15.75 |
| Salt | 1.10 | 1.30 |
| Premix 1 | 2.20 | 2.60 |
| Calcium hydrophosphate | 1.10 | 1.30 |
| Sodium bicarbonate | 1.10 | 1.30 |
| Chemical composition | ||
| CP, % of DM | 11.33 | 11.94 |
| NDF, % of DM | 41.75 | 33.84 |
| Ca, % of DM | 0.50 | 0.52 |
| P, % of DM | 0.43 | 0.48 |
| ME 2 MJ/kg DM | 10.42 | 10.59 |
| Item | CON | CAP | YSE | SEM | p-Value |
|---|---|---|---|---|---|
| Feed intake | |||||
| Dry matter intake, kg/day | 12.12 | 12.18 | 12.10 | 0.17 | 0.98 |
| Fermentation characteristics | |||||
| pH | 6.98 a | 7.01 a | 6.80 b | 0.02 | <0.01 |
| NH3-N, mg/100 mL | 5.16 b | 4.30 c | 5.68 a | 0.11 | <0.01 |
| TVFA, mmol/L | 51.75 b | 46.47 c | 70.91 a | 2.06 | <0.01 |
| Individual VFA, % of total VFA | |||||
| Acetic acid | 70.42 a | 71.11 a | 69.19 b | 0.25 | <0.01 |
| Propionic acid | 15.81 | 15.83 | 15.96 | 0.11 | 0.83 |
| Butyric acid | 9.54 b | 9.18 b | 11.21 a | 0.22 | <0.01 |
| Isobutyric acid | 1.53 a | 1.40 b | 1.19 c | 0.03 | <0.01 |
| Valeric acid | 0.65 ab | 0.60 b | 0.68 a | 0.01 | 0.04 |
| Isovaleric acid | 2.06 a | 1.88 b | 1.77 b | 0.04 | <0.01 |
| Ac:Pr | 4.46 | 4.50 | 4.34 | 0.04 | 0.21 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | CAP | YSE | |||
| Sobs | 323.47 c | 538.60 a | 428.00 b | 15.32 | <0.01 |
| Shannon | 5.37 c | 5.89 a | 5.63 b | 0.04 | <0.01 |
| Ace | 324.58 c | 549.75 a | 434.59 b | 16.09 | <0.01 |
| Coverage | 0.9942 | 0.9992 | 0.9965 | 0.0004 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, X.; Wu, B.; Ma, J.; Cui, X.; Deng, Z.; Hu, S.; Li, W.; A, R.; Li, X.; Meng, Q.; et al. Effects of Dietary Capsaicin and Yucca schidigera Extracts as Feed Additives on Rumen Fermentation and Microflora of Beef Cattle Fed with a Moderate-Energy Diet. Fermentation 2023, 9, 30. https://doi.org/10.3390/fermentation9010030
Yi X, Wu B, Ma J, Cui X, Deng Z, Hu S, Li W, A R, Li X, Meng Q, et al. Effects of Dietary Capsaicin and Yucca schidigera Extracts as Feed Additives on Rumen Fermentation and Microflora of Beef Cattle Fed with a Moderate-Energy Diet. Fermentation. 2023; 9(1):30. https://doi.org/10.3390/fermentation9010030
Chicago/Turabian StyleYi, Xin, Baoyun Wu, Jinglei Ma, Xiaojing Cui, Ziqi Deng, Sanlong Hu, Wei Li, Runa A, Xiang Li, Qingxiang Meng, and et al. 2023. "Effects of Dietary Capsaicin and Yucca schidigera Extracts as Feed Additives on Rumen Fermentation and Microflora of Beef Cattle Fed with a Moderate-Energy Diet" Fermentation 9, no. 1: 30. https://doi.org/10.3390/fermentation9010030