
Non-Saccharomyces Are Also Forming the Veil of Flor in Sherry Wines

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains Origin and Culture Media
2.2. Physiological Characterisation
2.2.1. Fermentation and Assimilation of Different Carbon and Nitrogen Sources
2.2.2. Ethanol Resistance
2.2.3. Determination of Extracellular Enzymatic Activities
β-Glucosidase Activity
Cellulase Activity
Protease Activity
Lipolytic Activity
Urease Activity
2.3. Biofilm-Forming Characterisation
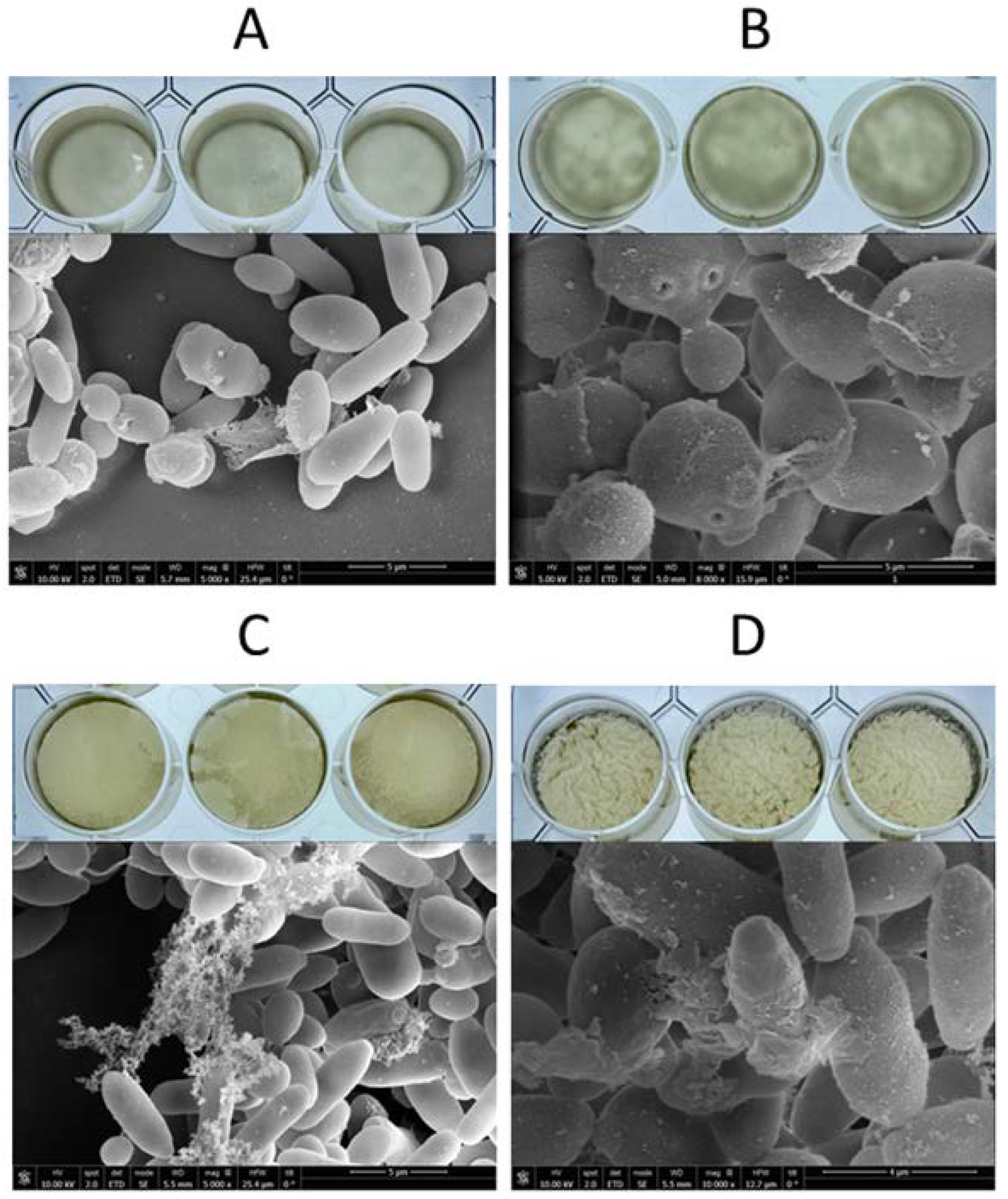
2.4. Scanning Electron Microscopy (SEM)
2.5. Statistical Analysis
3. Results
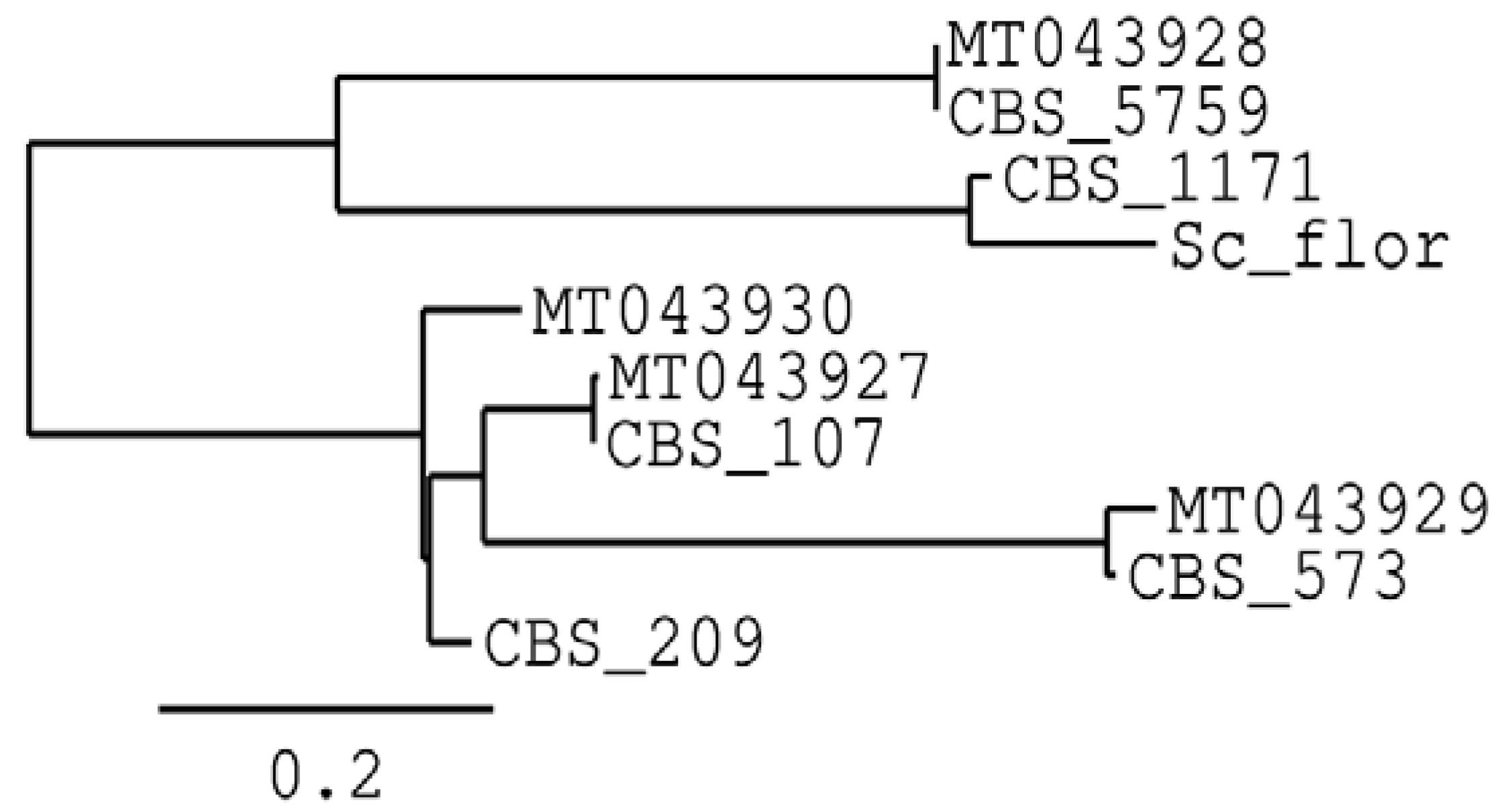
3.1. Characterisation of Non-Saccharomyces Strains
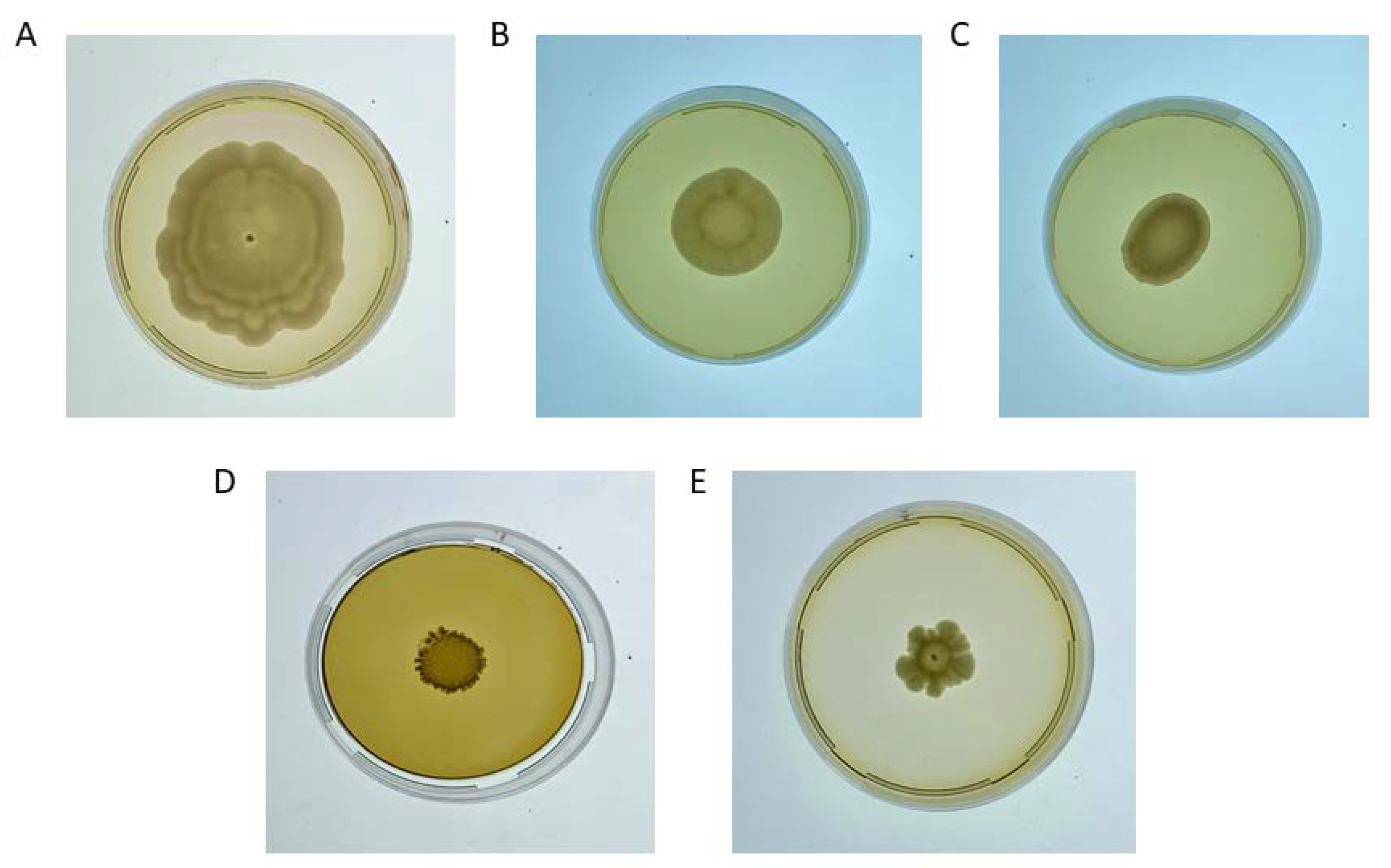
3.2. Biofilm-Forming Evaluation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cordero-Bueso, G.; Ruiz-Muñoz, M.; González-Moreno, M.; Chirino, S.; Bernal-Grande, M.C.; Cantoral, J.M. The microbial diversity of sherry wines. Fermentation 2018, 4, 19. [Google Scholar] [CrossRef]
- Martínez, P.; Pérez Rodríguez, L.; Benítez, T. Velum formation by flor yeasts isolated from sherry wine. Am. J. Enol. Vitic. 1997, 48, 55–62. [Google Scholar]
- Mesa, J.J.; Infante, J.J.; Rebordinos, L.; Cantoral, J.M. Characterization of yeasts involved in the biological ageing of sherry wines. LWT-Food Sci. Technol. 1999, 32, 114–120. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A. El metabolismo de las levaduras de flor. In Levaduras Vínicas: Funcionalidad Y Uso En Bodega; Mundi-Prensa: Madrid, Spain, 1997; pp. 173–189. ISBN 8471146851. [Google Scholar]
- Martínez, P.; Pérez Rodríguez, L.; Benítez, T. Evolution of flor yeast population during the biological aging of fino sherry wine. Am. J. Enol. Vitic. 1997, 48, 160–168. [Google Scholar]
- Esteve-Zarzoso, B.; Peris-Torán, M.J.; García-Maiquez, E.; Uruburu, F.; Querol, A. Yeast Population dynamics during the fermentation and biological aging of sherry wines. Appl. Environ. Microbiol. 2001, 67, 2056–2061. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces Cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef]
- Perpetuini, G.; Tittarelli, F.; Schirone, M.; di Gianvito, P.; Corsetti, A.; Arfelli, G.; Suzzi, G.; Tofalo, R. Adhesion properties and surface hydrophobicity of Pichia manshurica strains isolated from organic wines. LWT-Food Sci. Technol. 2018, 87, 385–392. [Google Scholar] [CrossRef]
- Saez, J.S.; Lopes, C.A.; Kirs, V.E.; Sangorrín, M. Production of volatile phenols by Pichia manshurica and Pichia membranifaciens isolated from spoiled wines and cellar environment in patagonia. Food Microbiol. 2011, 28, 503–509. [Google Scholar] [CrossRef]
- Morata, A.; Arroyo, T.; Bañuelos, M.A.; Blanco, P.; Briones, A.; Cantoral, J.M.; Castrillo, D.; Cordero-Bueso, G.; del Fresno, J.M.; Escott, C.; et al. Wine yeast selection in the iberian peninsula: Saccharomyces and non-Saccharomyces as drivers of innovation in Spanish and Portuguese wine industries. Crit. Rev. Food Sci. Nutr. 2022, 1, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Scansani, S.; van Wyk, N.; Nader, K.B.; Beisert, B.; Brezina, S.; Fritsch, S.; Semmler, H.; Pasch, L.; Pretorius, I.S.; von Wallbrunn, C.; et al. The film-forming Pichia spp. in a winemaker’s toolbox: A simple isolation procedure and their performance in a mixed-culture fermentation of Vitis Vinifera L. cv. Gewürztraminer must. Int. J. Food Microbiol. 2022, 365, 109549. [Google Scholar] [CrossRef] [PubMed]
- Loira, I.; Morata, A.; Bañuelos, M.A.; Suárez-Lepe, J.A. Isolation, selection, and identification techniques for non-Saccharomyces yeasts of oenological interest. In Biotechnological Progress and Beverage Consumption; Academic Press: Cambridge, MA, USA, 2020; pp. 467–508. [Google Scholar] [CrossRef]
- Comitini, F.; Ingeniis, J.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia anomala and Kluyveromyces wickerhamii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microbiol. Lett. 2004, 238, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; San Mauro, M.; Bravo, E.; Marquina, D. PMKT2, a new killer toxin from Pichia membranifaciens, and its promising biotechnological properties for control of the spoilage yeast Brettanomyces bruxellensis. Microbiology 2009, 155, 624–634. [Google Scholar] [CrossRef]
- Leça, J.M.; Pereira, V.; Miranda, A.; Vilchez, J.L.; Malfeito-Ferreira, M.; Marques, J.C. Impact of indigenous non-Saccharomyces yeasts isolated from Madeira Island vineyards on the formation of ethyl carbamate in the aging of fortified wines. Processes 2021, 9, 799. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, Y.; Wang, D.; Xu, Y. Immobilized Rhodotorula mucilaginosa: A novel urethanase-producing strain for degrading ethyl carbamate. Appl. Biochem. Biotechnol. 2013, 171, 2220–2232. [Google Scholar] [CrossRef]
- Fanning, S.; Mitchell, A.P. Fungal biofilms. PLoS Pathog. 2012, 8, e1002585. [Google Scholar] [CrossRef]
- Chi, M.; Li, G.; Liu, Y.; Liu, G.; Li, M.; Zhang, X.; Sun, Z.; Sui, Y.; Liu, J. Increase in antioxidant enzyme activity, stress tolerance and biocontrol efficacy of Pichia kudriavzevii with the transition from a yeast-like to biofilm morphology. Biol. Control 2015, 90, 113–119. [Google Scholar] [CrossRef]
- Di Maro, E.; Ercoloni, D.; Coppola, S. Yeast dynamics during pontaneous wine fermentation of the Catalanesca grape. Int. J. Food Microbiol. 2007, 117, 201–210. [Google Scholar] [CrossRef]
- Ruiz-Muñoz, M.; Cordero-Bueso, G.; Benítez-Trujillo, F.; Martínez, S.; Pérez, F.; Cantoral, J.M. Rethinking about flor yeast diversity and its dynamic in the “criaderas and soleras” biological aging system. Food Microbiol. 2020, 92, 103553. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S RRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large Subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Granchi, L.; Guerrini, S.; Mangani, S.; Romaniello, R.; Vincenzini, M.; Romano, P. Diversity of Saccharomyces cerevisiae strains isolated from two italian wine-producing regions. Front. Microbiol. 2016, 7, 1018. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. Yeasts: A Taxonomic Study, 5th ed; Kurtzman, C., Fell, J.W., Boekhout, T., Eds.; Elvesier: Saint Louis, MO, USA, 2011; ISBN 9780080931272. [Google Scholar]
- Ruiz-Muñoz, M.; Bernal-Grande, M.; Cordero-Bueso, G.; González, M.; Hughes-Herrera, D.; Cantoral, J. A microtiter plate assay as a reliable method to assure the identification and classification of the veil-forming yeasts during sherry wines ageing. Fermentation 2017, 3, 58. [Google Scholar] [CrossRef]
- Aranda, A.; Querol, A.; del Olmo, M. Correlation between acetaldehyde and ethanol resistance and expression of HSP genes in yeast strains isolated during the biological aging of sherry wines. Arch. Microbiol. 2002, 177, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Gaensly, F.; Agustini, B.C.; da Silva, G.A.; Picheth, G.; Bonfim, T.M.B. Autochthonous yeasts with β-glucosidase activity increase resveratrol concentration during the alcoholic fermentation of Vitis labrusca grape must. J. Funct. Foods 2015, 19, 288–295. [Google Scholar] [CrossRef]
- Mangunwardoyo, W.; Sjamsuridzal, W.; Oetari, A. Screening cellulose activity of yeast isolated from soil, sediment and water river from Taman Nasional Gunung Halimun, West Java, Indonesia. Malays. J. Microbiol. 2011, 7, 210–216. [Google Scholar] [CrossRef]
- Sierra, G. A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie van Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef]
- Moreno-García, J.; García-Martínez, T.; Moreno, J.; Millán, M.C.; Mauricio, J.C. A proteomic and metabolomic approach for understanding the role of the flor yeast mithocondria in the velum formation. Int. J. Food Microbiol. 2014, 172, 21–29. [Google Scholar] [CrossRef]
- Silva-Dias, A.; Miranda, I.M.; Branco, J.; Monteiro-Soares, M.; Pina-Vaz, C.; Rodrigues, A.G. Adhesion, biofilm formation, cell surface hydrophobicity, and antifungal planktonic susceptibility: Relationship among Candida spp. Front. Microbiol. 2015, 6, 205. [Google Scholar] [CrossRef]
- Zara, G.; Farbo, M.G.; Multineddu, C.; Migheli, Q.; Budroni, M.; Zara, S.; Mannazzu, I. Exploring the biodiversity of red yeasts for in vitro and in vivo phenotypes relevant to agri-food-related processes. Fermentation 2021, 7, 2. [Google Scholar] [CrossRef]
- Hovnanyan, K.; Martuyan, S.; Pepoyan, A.; Navasardyan, L.; Trchounian, A. Transmission and scanning electron microscopy of contacts between bacterial and yeast cells in biofilms on different surfaces. Open Access Libr. J. 2015, 02, 68321. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny. fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Castrillón, M.; Mendes, S.D.C.; Inostroza-Ponta, M.; Valente, P. (GTG)5 MSP-PCR fingerprinting as a technique for discrimination of wine associated yeasts? PLoS ONE 2014, 9, e105870. [Google Scholar] [CrossRef]
- Malfeito-Ferreira, M. Yeasts and wine off-flavours: A technological perspective. Ann. Microbiol. 2011, 61, 95–102. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Challenges of the non-conventional yeast Wickerhamomyces anomalus in winemaking. Fermentation 2018, 4, 68. [Google Scholar] [CrossRef]
- Lin, M.M.H.; Boss, P.K.; Walker, M.E.; Sumby, K.M.; Grbin, P.R.; Jiranek, V. Evaluation of indigenous non-Saccharomyces yeasts isolated from a South Australian vineyard for their potential as wine starter cultures. Int. J. Food Microbiol. 2020, 312, 108373. [Google Scholar] [CrossRef]
- Maturano, Y.P.; Rodríguez Assaf, L.A.; Toro, M.E.; Nally, M.C.; Vallejo, M.; de Figueroa, L.I.C.; Combina, M.; Vazquez, F. Multi-enzyme production by pure and mixed cultures of Saccharomyces and non-Saccharomyces yeasts during wine fermentation. Int. J. Food Microbiol. 2012, 155, 43–50. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Rodríguez-Gómez, F.; Barrio, E.; Querol, A.; Garrido-Fernández, A.; Arroyo-Lopez, F.N. Exploring the yeast biodiversity of green table olive industrial fermentations for technological applications. Int. J. Food Microbiol. 2011, 147, 89–96. [Google Scholar] [CrossRef]
- Baffi, M.A.; Tobal, T.; Lago, J.H.G.; Boscolo, M.; Gomes, E.; Da-Silva, R. Wine aroma improvement using a β-Glucosidase preparation from Aureobasidium pullulans. Appl. Biochem. Biotechnol. 2013, 169, 493–501. [Google Scholar] [CrossRef]
- Mateo, J.J.; Peris, L.; Ibañez, C.; Maicas, S. Characterization of glycolytic activities from non-Saccharomyces yeasts isolated from bobal musts. J. Ind. Microbiol. Biotechnol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Claus, H.; Mojsov, K. Enzymes for wine fermentation: Current and perspective applications. Fermentation 2018, 4, 52. [Google Scholar] [CrossRef]
- Madrigal, T.; Maicas, S.; Mateo Tolosa, J.J. Glucose and ethanol tolerant enzymes produced by Pichia (Wickerhamomyces) isolates from enological ecosystems. Am. J. Enol. Vitic. 2013, 64, 126–133. [Google Scholar] [CrossRef]
- Coi, A.L.; Bigey, F.; Mallet, S.; Marsit, S.; Zara, G.; Gladieux, P.; Galeote, V.; Budroni, M.; Dequin, S.; Legras, J.L. Genomic signatures of adaptation to wine biological ageing conditions in biofilm-forming flor yeasts. Mol. Ecol. 2017, 26, 2150–2166. [Google Scholar] [CrossRef] [PubMed]
- Eldarov, M.A.; Beletsky, A.V.; Tanashchuk, T.N.; Kishkovskaya, S.A.; Ravin, N.V.; Mardanov, A.V. Whole-genome analysis of three yeast strains used for production of sherry-like wines revealed genetic traits specific to flor yeasts. Front. Microbiol. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 14812. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J.; Basehoar-Powers, E. Phylogenetic relationships among species of Pichia, Issatchenkia and Williopsis determined from multigene sequence analysis, and the proposal of Barnettozyma gen. nov., Lindnera gen. nov. and Wickerhamomyces gen. nov. FEMS Yeast Res. 2008, 8, 939–954. [Google Scholar] [CrossRef]
- Douglass, A.P.; Offei, B.; Braun-Galleani, S.; Coughlan, A.Y.; Martos, A.A.R.; Ortiz-Merino, R.A.; Byrne, K.P.; Wolfe, K.H. Population genomics shows no distinction between pathogenic Candida krusei and environmental Pichia kudriavzevii: One species, four names. PLoS Pathog. 2018, 14, e1007138. [Google Scholar] [CrossRef]
- Carbonero-Pacheco, J.; Moreno-García, J.; Moreno, J.; García-Martínez, T.; Mauricio, J.C. revealing the yeast diversity of the flor biofilm microbiota in sherry wines through internal transcribed spacer-metabarcoding and matrix-assisted laser desorption/ionization time of flight mass spectrometry. Front. Microbiol. 2022, 12, 825756. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| P. manshurica | P. membranifaciens | P. kudriavzevii | W. anomalus | ||
|---|---|---|---|---|---|
| Fermentation | Glucose | + | + | − | + |
| Galactose | + | − | + | + | |
| Maltose | − | − | + | − | |
| Sucrose | + | − | − | + | |
| Assimilation | Glucose | + | + | + | − |
| Ethanol | + | − | + | − | |
| Glycerol | + | + | + | + | |
| Citrate | + | − | + | − | |
| Nitrite | + | − | + | − | |
| Urea | + | − | − | − | |
| Extracellular activities | Lipolytic | + | − | + | + |
| Proteolytic | + | − | − | − | |
| Cellulolytic | − | + | − | + | |
| Urease | + | − | − | − | |
| β-glucosidase | − | − | + | + |
| Biofilm Formation (Days) | Hydrophobicity (%) | Ethanol Resistance (%) | Adhesion (ΔAbs570) | |
|---|---|---|---|---|
| P. manshurica | 7 ± 0.22 c | 93 ± 1.25 b | 17.00 ± 0.25 a | 6.07 ± 0.23 c |
| P. membranifaciens | 13 ± 0.42 a | 96 ± 3.20 a | 15.50 ± 0.55 b | 10.32 ± 0.18 a |
| P. kudriavzevii | 11 ± 0.31 b | 94 ± 2.20 ab | 16.00 ± 0.60 ab | 8.72 ± 0.13 b |
| W. anomalus | 9 ± 0.55 bc | 88 ± 2.65 c | 16.50 ± 0.20 a | 8.82 ± 0.09 b |
| S. cerevisiaeflor | 12 ± 0.34 b | 95 ± 1.85 a | 16.00 ± 0.30 ab | 7.78 ± 0.11 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Muñoz, M.; Hernández-Fernández, M.; Cordero-Bueso, G.; Martínez-Verdugo, S.; Pérez, F.; Cantoral, J.M. Non-Saccharomyces Are Also Forming the Veil of Flor in Sherry Wines. Fermentation 2022, 8, 456. https://doi.org/10.3390/fermentation8090456
Ruiz-Muñoz M, Hernández-Fernández M, Cordero-Bueso G, Martínez-Verdugo S, Pérez F, Cantoral JM. Non-Saccharomyces Are Also Forming the Veil of Flor in Sherry Wines. Fermentation. 2022; 8(9):456. https://doi.org/10.3390/fermentation8090456
Chicago/Turabian StyleRuiz-Muñoz, Marina, María Hernández-Fernández, Gustavo Cordero-Bueso, Sergio Martínez-Verdugo, Fernando Pérez, and Jesús Manuel Cantoral. 2022. "Non-Saccharomyces Are Also Forming the Veil of Flor in Sherry Wines" Fermentation 8, no. 9: 456. https://doi.org/10.3390/fermentation8090456