1. Introduction
Modern consumers demand food products that are healthy and safe. Fortification with natural bioactive compounds is a means to enhance the nutritional and health benefits of foods and beverages. For instance, milk is commonly fermented with lactic acid bacteria to produce wholesome, shelf-stable, probiotic-rich foods such as yogurt, cheese, and sour milk [
1,
2]. The consumption of probiotic products provides health benefits such as treatment of diarrhea, alleviation of lactose intolerance, and enhancement of immunity [
3]. Traditionally, probiotics were commonly recognized to be derived from dairy-based products. In recent times, non-dairy foods have been used to isolate potential probiotics [
4].
Bacillus subtilis, a spore-foaming non-lactic acid producer, has also been used as a probiotic in fermented foods and supplements [
5].
Bacillus spp. are valuable microorganisms for the production of legume-based fermented foods. They can also generate valuable bioactive compounds such as poly-γ-glutamic acid (γ-PGA), hydrolytic enzymes, peptides, and probiotics [
6,
7].
Bacillus spp. are used to produce traditional Korean fermented foods like chungkookjang, characterized by its strong smell and sticky consistency. Nevertheless, fermentation of milk with
Bacillus spp. is uncommon because of the unpleasant odor of the resulting products.
γ-PGA is an anionic, naturally occurring homo-polyamide composed of d- and l-glutamic acid units connected by amide linkages between α-amino and γ-carboxylic acid groups. γ-PGA is water soluble, biodegradable, edible, and non-toxic to humans and the environment. This biopolymer is one of the main components of the mucilage produced by
Bacillus strains, such as the sticky substance found in chungkookjang. The Korea Food and Drug Administration certifies γ-PGA as a functional ingredient for boosting immunity and calcium absorption in humans [
8].
Therefore, potential applications of γ-PGA and its derivatives in food, cosmetics, and medicines are of interest [
9]. Lactic acid bacteria are considered to be probiotics and are widely used in food fermentation because of their contribution to human health; they also have a generally recognized as safe (GRAS) status. Lactic acid fermentation enhances the safety, organoleptic attributes, nutritional value, and health benefits of foods [
10]. Some foods produced by lactic fermentation are yoghurt, kefir, sauerkraut, and kimchi. Lactic acid bacteria such as
Lactobacillus brevis,
L. plantarum,
L. delbrueckii,
L. paracasei, and
Lactococcus lactis show glutamate decarboxylase activity, which is needed for the production of gamma-aminobutyric acid (GABA) [
11].
Lactobacillus brevis, as a functional dairy starter, has been used for manufacturing novel fermented dairy foods with high GABA content [
12].
GABA has numerous roles in physiological processes, including neurotransmission as well as hypotensive, diuretic, and tranquilizer effects. Natural and conventional treatments such as mechanical and cold stimulation encourage GABA biosynthesis. The addition of exogenous glutamic acid or gibberellins increases GABA production [
13,
14]. Biosynthesis of GABA may be much more promising than chemical synthesis since it involves a simple bioreaction process with high catalytic efficiency. The biosynthesis of GABA depends on the bioconversion of glutamate to GABA, which is catalyzed by glutamate decarboxylase (GAD) [
15].
In this study, a novel co-fermentation using Bacillus subtilis HA and Lactiplantibacillus plantarum EJ2014 was optimized to produce milk fortified with bioactive compounds and improved flavor. Fermentation with B. subtilis in the presence of milk containing MSG aimed to produce γ-PGA and peptides. The second lactic acid fermentation with Lactiplantibacillus plantarum was performed to produce GABA and stabilize the co-fermented milk. It is expected that this study will provide valuable information regarding milk fortification by stepwise co-fermentation, which can be used to develop healthy fermented dairy products for the market.
2. Materials and Methods
2.1. Materials
Monosodium-ʟ-glutamate (MSG) and glucose were obtained from CJ Cheil Jedang Corporation (Seoul, Korea) and Samyang Genex Corporation (Incheon, Korea), respectively. Milk was purchased from Maeil Dairies Co. Ltd. (Seoul, Korea).
2.2. Strains and Starter Cultures
The starter of B. subtilis HA (KCCM 10775P) was cultured in nutrient broth (NB, Becton, Dickinson and Company, Sparks, MD, USA) at 42 °C for 24 h. The cells were harvested and suspended in 10 times the volume of sterilized distilled water.
Lactiplantibacillus plantarum EJ2014 (KCCM 11545P) was grown on DifcoTM Lactobacilli MRS agar (Becton, Dickinson, and Company) at 30 °C for 48 h. Then, a single colony was inoculated in sterilized MRS broth and cultured at 30 °C for 24 h to prepare the starter culture.
2.3. Co-Fermentation Process
The milk was mixed with sterilized MSG and glucose solutions. The final concentrations were adjusted to 3% MSG and 2.5% glucose. The starter of B. subtilis HA was grown in nutrient broth (NB), and cells harvested were suspended in 1/10 the volume of distilled water. The starter culture was inoculated at a 5% concentration and incubated by shaking (160 rpm; SI-900R, Jeio Tech. Co., Ltd., Daejeon, Korea) at 42 °C for 1 day.
Subsequently, for the co-fermentation with lactic acid bacteria, 1.5% MSG was added. The starter of L. plantarum EJ2014 grown in MRS broth was inoculated at a 1% concentration, followed by incubation (HB-201MS-4, HANBAEK Scientific Co., Ltd., Incheon, Korea) at 30 °C for 5 days.
2.4. Viable Bacterial Counts
To determine the viable bacterial counts of B. subtilis, the fermented milk was serially diluted with sterilized physiological saline. Subsequently, a 20-μL sample was plated onto an MRS agar plate and cultured at 42 °C for 24 h to confirm the presence of colony-forming units (CFU). To selectively determine the viable bacterial counts of L. plantarum EJ2014 after the co-fermentation, the MRS agar plate with mixed strains was cultured at 30 °C for 48 h to delay the growth of B. subtilis, and the colonies of L. plantarum were counted.
2.5. pH and Acidity
The pH of the fermented broth was determined by a pH meter (Model 420+, Thermo Orion, Beverly, MA, USA). Titratable acidity (%, w/v) was determined as lactic acid by measuring the amount of 0.1 M NaOH required to reach a pH of 8.3.
2.6. Consistency Index
The fermented broth (5 mL) was centrifuged at 24,948× g for 20 min. The supernatant was mixed with twice the volume of isopropanol, and the precipitate was washed with 95% ethanol and dried at 60 °C for 24 h to yield crude mucilage. The consistency of the fermented broth was determined using a rheometer (HAKKE RheoStress 1, Karlsruhe, Germany) fitted with a cone plate device (Plate PP35Ti, diameter 3.5 cm, 1.0 mm gap) at 20 °C. The sample (1 mL) was loaded between the plate and the cone plate device.
2.7. Mucilage Analysis
To determine total carbohydrates in the mucilage of the fermented milk, the phenol-sulfuric acid method [
16] was used. This is a simple and rapid colorimetric method that can detect all kinds of carbohydrates by visualizing a yellow-gold color. The protein content in the fermented milk was determined using the Pierce BCA Protein Assay Kit (Thermo Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. The crude fat content in the fermented milk was determined by the Rose-Gottlieb method.
To determine the total amino acid content, the mucilage was hydrolyzed in 6 M HCl at 110 °C for 22 h and filtered through a 0.45-μm syringe filter. Amino acid derivatization with AccQ•Tag reagents (Waters, Milton, MA, USA) was conducted according to the manufacturer’s protocol. Hydrolyzed sample (10 μL) was mixed with 70 μL of AccQ•Tag Ultra borate buffer and 20 μL of AccQ-Fluor reagent, followed by heating for 10 min at 55 °C. The sample was analyzed by HPLC (Alliance 2695, Waters) using an AccQ•Tag column (Waters; 3.9 × 150 mm) at 25 °C. A gradient elution was performed at a flow rate of 1 mL/min for 55 min, using a mixture of Waters AccQ•Tag Eluent A: distilled water (1:10) as eluent A, acetonitrile as eluent B, and distilled water as eluent C. Absorbance was measured at λ excitation (250 nm) and λ emission (395 nm) using a fluorescence detector.
2.8. Analysis of Free Sugars
Free sugars were analyzed using a Knauer K-501 HPLC (Berlin, Germany) fitted with a Shodex Asahipak NH2P-50 column (Tokyo, Japan) and a refractive index (RI) detector. Mucilage in the fermented milk was eliminated by washing with isopropyl alcohol. Subsequently, the mucilage-free supernatant was dried using a speed vacuum concentrator. The sample was re-suspended in distilled water and filtered through a 0.45-μm syringe filter. The column was kept at 30 °C, and isocratic elution was performed at a flow rate of 1.0 mL/min for 30 min, using 75% acetonitrile.
2.9. Analysis of Protein Hydrolysis
The protein in the co-fermented milk was analyzed by SDS-PAGE. As a pretreatment, 12.5 μL of supernatant was centrifuged at 15,710× g for 10 min, and then mixed with an equal volume of 2× Laemmli sample buffer containing β-mercaptoethanol (19:1) and heated at 100 °C for 5 min to induce denaturation. Electrophoresis was performed at 120 V for 50 min. The resulting gel was stained with Instant Blue solution (Expedeon Ltd., Cambridgeshire, UK), and the molecular weights of the proteins were estimated visually by their mobilization in the gel. A Page Ruler Prestained Protein Ladder (10–180 kDa; Thermo Scientific) was used as the molecular weight marker.
2.10. Protease Activity Assay
To determine the protease activity, a modified Anson’s method was applied [
17]. Briefly, the fermented broth (0.35 mL) was diluted 10-fold using distilled water, mixed with an equal volume (0.35 mL) of 0.6% casein, and then heated at 37 °C for 10 min. The sample (0.7 mL) was mixed with 0.7 mL of 0.44 M trichloroacetic acid (TCA) and incubated at 37 °C for 30 min. The precipitate was removed by centrifugation at 15,710×
g for 10 min, and the supernatant (1 mL) was mixed with 2.5 mL of 0.55 M Na
2CO
3 and 0.5 mL of Folin-phenol reagent. After incubating at 37 °C for 30 min, absorbance was measured using a spectrophotometer (Ultrospec
®2100 Pro, Amersham Biosciences, Piscataway, NJ, USA) at 660 nm. The peptide content in the supernatant was determined using tyrosine as a standard.
2.11. Analysis of GABA and Glutamic Acid
The contents of GABA and glutamic acid were qualitatively analyzed by thin-layer chromatography (TLC) using a silica gel plate (10 cm × 20 cm). TLC development was carried out in a chamber (30 cm × 25 cm × 10 cm), and 0.5% of each standard was used to compare the residual amounts of MSG and GABA. The developing solvent was n-butyl alcohol: acetic acid glacial: distilled water in a 3:1:1 (v/v) ratio. The solvent was poured into the chamber and allowed to saturate it at room temperature for 4 h. The fermentation product was diluted threefold, and then each sample and standard solution (2 µL) was spotted. The plate was dried and developed using 0.2% ninhydrin followed by heating at 100 °C for 5 min or until spots appeared.
The GABA and glutamic acid contents in the co-fermented milk were also determined by ultra-performance liquid chromatography-mass spectrometry (UPLC/MS, Waters), fitted with an Infinitylab Poroshell 120 column and a QDA detector (Waters). A modified method for the analysis of underivatized amino acids was used [
18]. A gradient elution was performed at a flow rate of 0.7 mL/min for 15 min, using water with 0.1% formic acid as eluent A and acetonitrile with 0.1% formic acid as eluent B. GABA and glutamic acid were used to create a standard curve.
2.12. Volatile Compound Analysis
The volatile composition of the co-fermented milk was analyzed by GC/MS (Agilent 7890B-5977B, Santa Clara, CA, USA) fitted with a DB-WAX column (Agilent). Helium (He) was used as a carrier gas. Samples were collected using solid-phase microextraction (SPME) with a CAR/DVB/PDMS fiber (Sigma-Aldrich). After sample injection, the column temperature was maintained at 40 °C for 2 min, raised to 220 °C at a rate of 2 °C/min, subsequently increased to 20 °C/min until reaching 240 °C, and kept at this temperature for 10 min.
2.13. Statistical Analysis
Statistical analysis was performed using the Statistical Package for the Social Sciences (SPSS Version 25.0, SPSS Inc., Chicago, IL, USA). The mean and standard error were calculated. Significant differences among means were determined by one way-ANOVA and Duncan’s multiple range test (p < 0.05).
3. Results and Discussion
3.1. pH, Acidity, and Viable Bacterial Counts after Fermentation
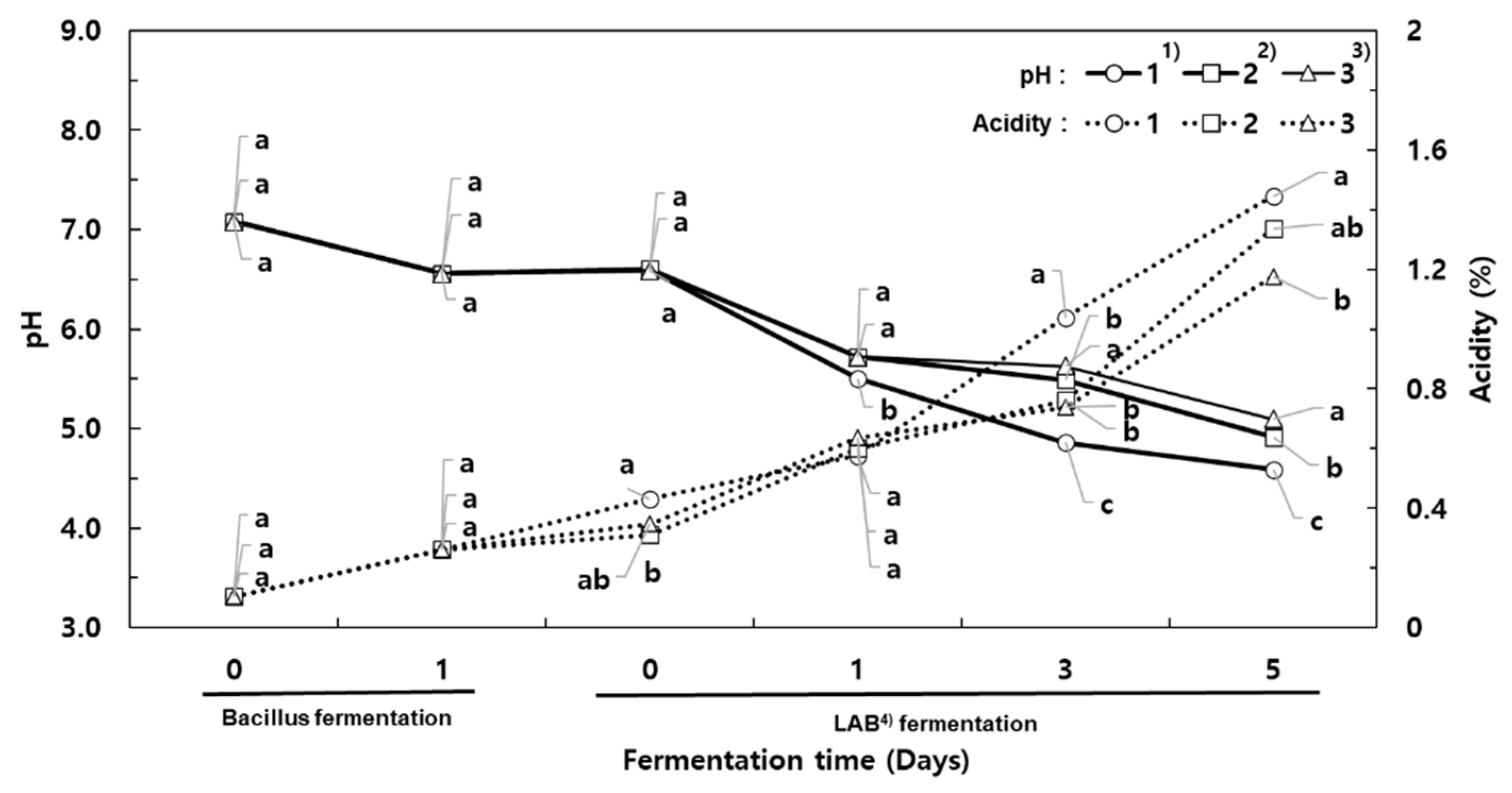
During the fermentation with
B. subtilis HA, the pH of the milk decreased from 7.08 to 6.56, but acidity increased from 0.11% to 0.26% (
Figure 1). Generally, fermentation of soybean with
Bacillus spp. results in increased pH. However, in this study, fermentation of milk with
B. subtilis HA resulted in acid production and thus a lowered pH. It was reported that fermentation with
B. coagulans,
B. macerans, and
B. licheniformis resulted in high acid production, whereas fermentation with
B. subtilis HA resulted in low acid production [
19]. However, we observed the opposite outcome, with
B. subtilis HA producing acid and decreasing the pH, which may be explained by the type of substrate (milk) used for the fermentation.
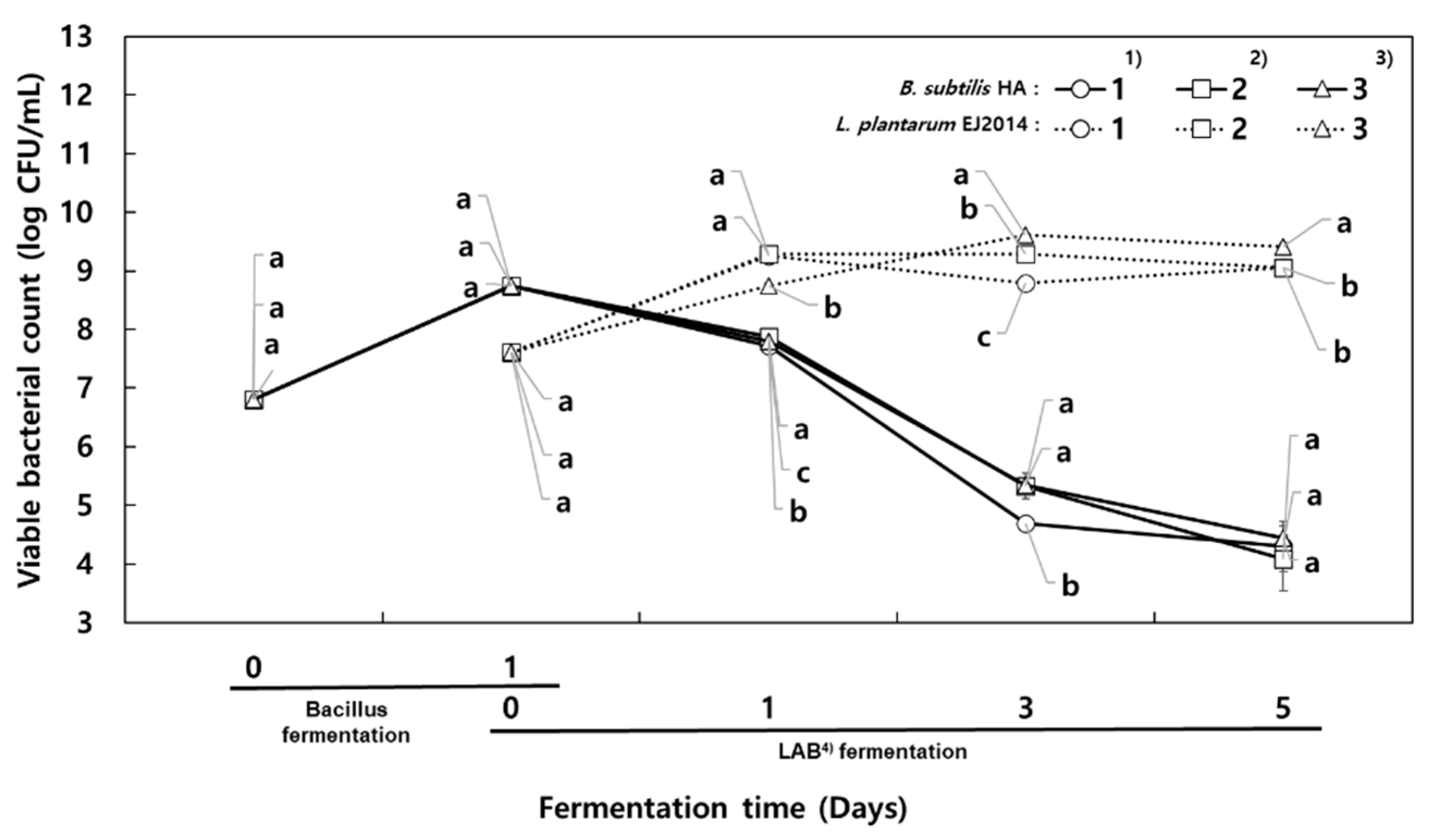
The initial viable bacterial count of
B. subtilis HA was 6.80 log CFU/mL, which increased to 8.74 log CFU/mL after 1 day (
Figure 2). It should be noted that
Bacillus strains can be potential probiotics because of their thermostable and acid-resistant properties [
20], and so their increase is desirable.
For the co-fermentation, the incubation temperature was lowered from 42 °C to 30 °C, and MSG was added at 1.5% and 3% concentrations. The viable cell counts of B. subtilis HA in the milk co-fermented for 5 days were greatly decreased, indicating 4.30 log CFU/mL (control), 4.09 log CFU/mL (1.5% MSG), and 4.44 log CFU/mL (3% MSG). On the other hand, the viable cell counts of L. plantarum increased from 7.61 log CFU/mL to 9.06 log CFU/mL (control), 9.06 log CFU/mL (1.5% MSG), and 9.42 log CFU/mL (3% MSG).
Therefore, B. subtilis can support the growth of lactic acid bacteria, resulting in a suitable environment for producing higher amounts of GABA by lactic fermentation. The initial pH of the co-fermented milk was 6.59 (control), 6.60 (1.5% MSG), and 6.59 (3% MSG), which then decreased to 4.59, 4.91, and 5.09, respectively. The acidity of the milk co-fermented without MSG (control) increased to 1.45% by the end of fermentation. By contrast, the acidity of the co-fermented milk supplemented with MSG was about 0.75% after 3 days, and increased above 1% after 5 days. These results indicate that the decrease in acidity during the second fermentation was related to the MSG concentration added for the production of GABA.
3.2. Mucilage and Viscosity Evaluation
Bacillus subtilis produces mucilage containing γ-PGA during the alkaline fermentation of soybean, herbs used in Oriental medicine, and plant extracts [
6,
21]. Mucilage production depends upon the medium used for the fermentation. It was reported that mucilage production by
B. subtilis in a defined medium varied according to the medium composition (i.e., carbon and nitrogen sources and magnesium and manganese concentration) [
22].
Here, the milk fermented by B. subtilis HA was viscous and had a high consistency index (1.82 Pa·Sn) due to the high production of crude mucilage (4.65%). The enhanced viscosity of the fermented milk encouraged the bioconversion of glutamic acid into γ-PGA.
3.3. Mucilage Composition
The γ-PGA, total sugar, crude protein, and crude fat content in crude mucilage were determined. The crude mucilage contained 15.18% carbohydrate, 18.85% crude protein, and 30.04% crude fat. The analysis of total amino acids revealed that the hydrolyzed crude mucilage contained 35.93% glutamic acid, implying that it was composed of about 36% γ-PGA. In addition, milk fat was a major fraction of the crude mucilage. By comparison, crude mucilage obtained from the fermentation of soybean with
Bacillus spp. is composed of over 60% γ-PGA, and that obtained in chemically defined media contains about 96.9% γ-PGA [
22,
23]. Thus, the fermented milk had a high mucilage content in addition to fat, although the mucilage content was lower than that obtained using soybeans or a defined medium.
3.4. Thin Layer Chromatography (TLC) Analysis
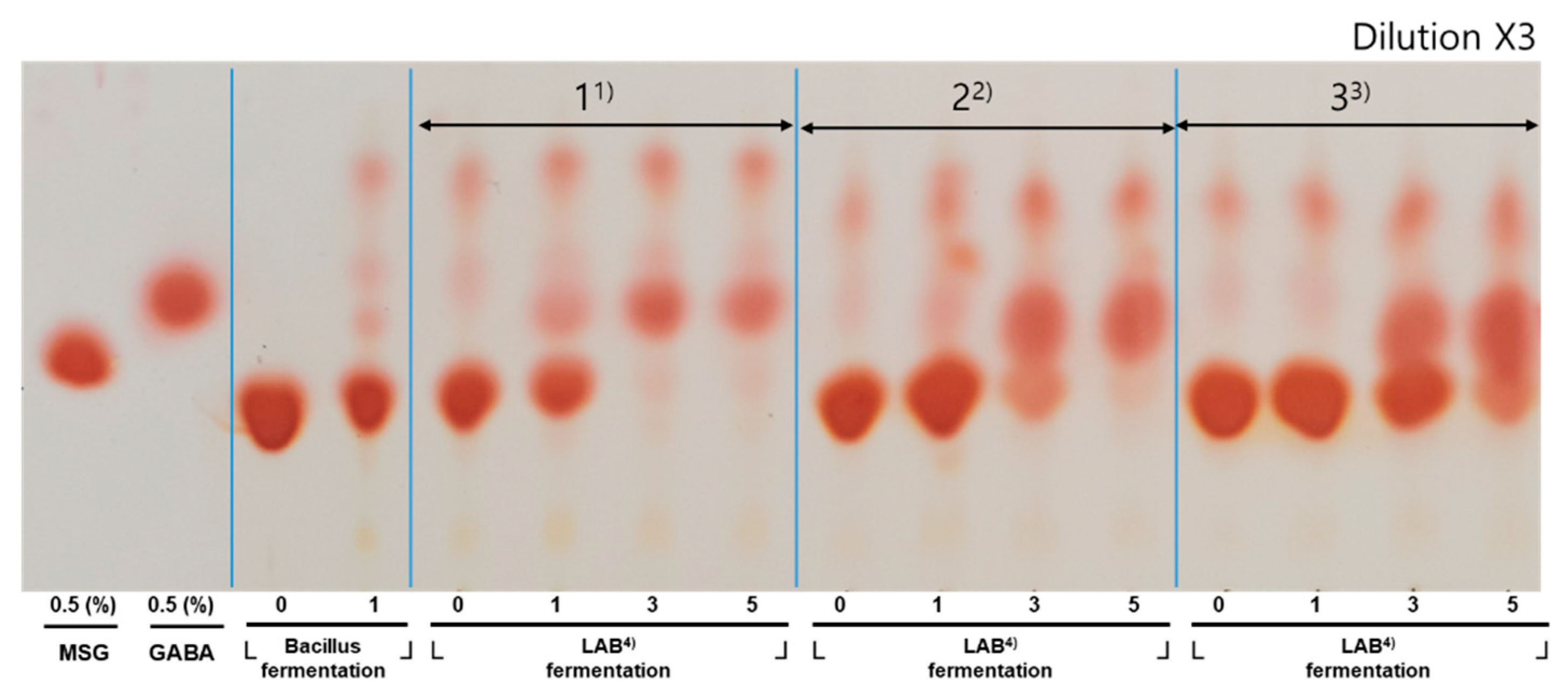
GABA production in the co-fermented milk with various MSG concentrations was monitored by TLC (
Figure 3). The conversion of MSG into GABA was greatly dependent upon the MSG concentration, which is expected because MSG is a GABA precursor. During the first fermentation, MSG was consumed by
B. subtilis HA, resulting in the production of mucilage. During the second fermentation by
L. plantarum EJ2014, the co-fermented milk without MSG showed low production of GABA. However, a higher content of GABA was obtained when MSG was added to the milk. A concentration of 1.5% MSG was optimal for the fortification of GABA in the co-fermented milk as higher concentrations resulted in MSG residues in the co-fermented product, which would be undesirable.
3.5. Saccharide Quantification
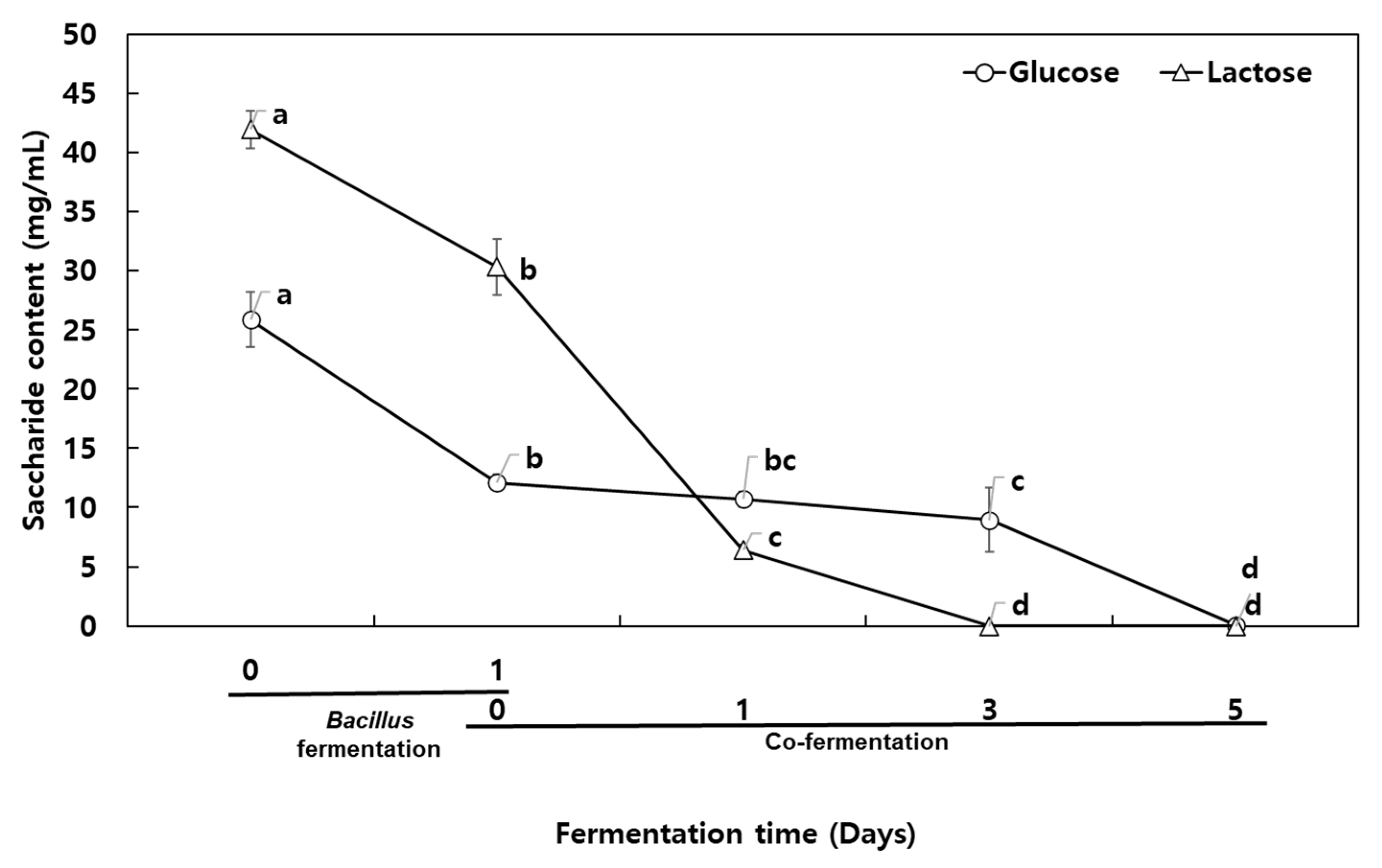
The milk was fortified with 2.5% glucose to enhance bacterial growth during the first alkaline fermentation. The initial glucose and lactose contents of the milk broth were 25.88 mg/g and 41.96 mg/g, respectively. Both glucose and lactose were consumed during the first day of the fermentation, showing a gradual decrease. In particular,
B. subtilis HA showed lactose-fermenting ability, decreasing lactose from 42 mg/g to 30.3 mg/g. During the co-fermentation with
L. plantarum EJ2014, the glucose content was gradually decreased while the lactose was consumed completely. The lactic acid fermentation resulted in an acidic environment, which is unsuitable for the growth of
Bacillus spp. Moreover, total fermenting sugar remained at a 1% concentration after 2 days of co-fermentation. After 5 days of co-fermentation, glucose and lactose were completely consumed by the bacteria (
Figure 4). This suggests that the fermenting sugar content may play an important role in microbial GABA production.
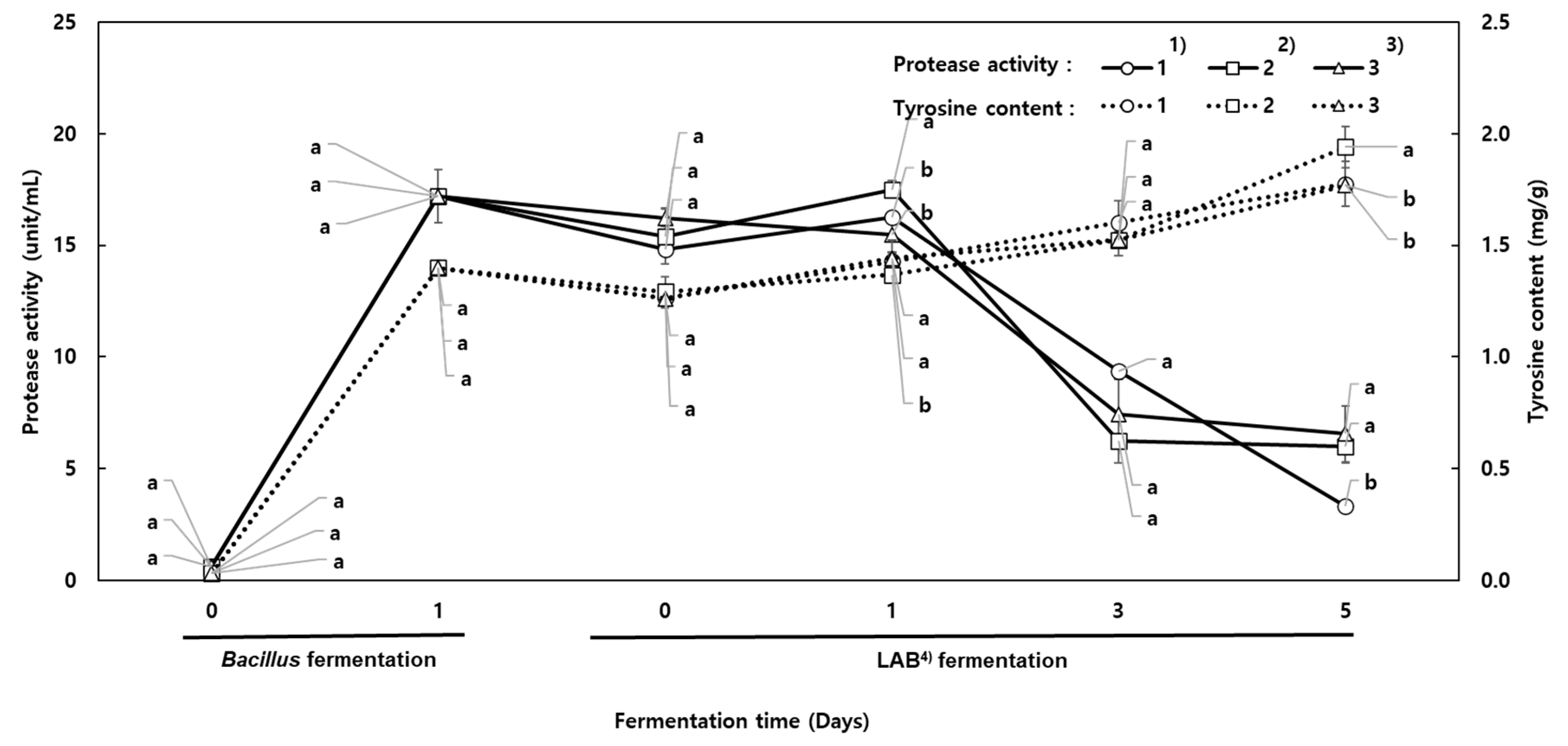
3.6. Milk Protein Hydrolysis
B. subtilis produces protease, which hydrolyzes proteins, resulting in the production of peptides [
24]. The protease activity and tyrosine equivalent content were analyzed in the co-fermented milk. The tyrosine equivalent content increased as more peptides were generated during protein hydrolysis. As shown in
Figure 5, the protease activity was greatly increased during the first day of the alkaline fermentation with
B. subtilis HA, resulting in a higher tyrosine equivalent content. Protease activity indicated 17.21 U/mL and the tyrosine equivalent content also increased from 0.03 mg/g to 1.40 mg/g. When MSG was added, the protease activity in the milk co-fermented for 1 day ranged from 15.49 to 17.50 U/mL. Afterward, the protease activity gradually decreased to values ranging from 3.30 to 7.12 U/mL. During the 5 days of co-fermentation, the tyrosine equivalent content was gradually increased to 1.94 mg/g. However, the protease activity decreased, indicating that in acidic conditions, the protease produced by
B. subtilis hydrolyzed casein into peptides. Peptides have been reported to show bioactive properties such as enhanced absorption in the gut and angiotensin-converting enzyme inhibition; thus, peptide generation by fermentation is desirable [
25].
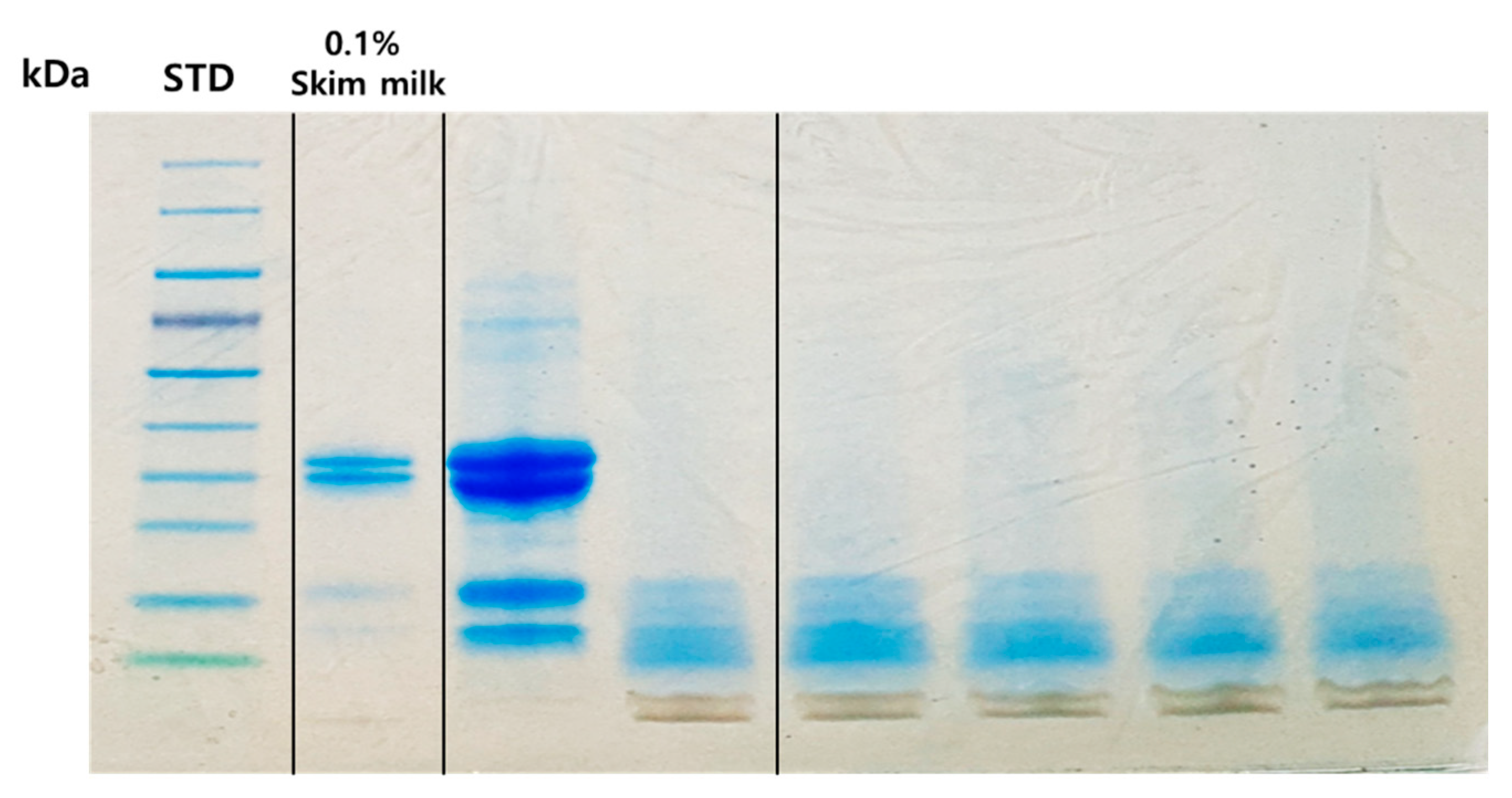
The hydrolyzed pattern of casein by SDS-PAGE analysis is shown in
Figure 6. Casein as a major milk protein and β-lactoglobulin were hydrolyzed during the first day of the alkaline fermentation, resulting in the production of peptides with low molecular weight (<10 kDa). Both α-casein and β-casein were completely hydrolyzed because of effective enzymatic digestion due to their fibrous structure. However, β-lactoglobulin (a globular protein), remained as a faint fraction after digestion. The most thoroughly digested protein fraction could not be detected in SDS-gel because of its low molecular weight. Therefore, alkaline fermentation of milk with
B. subtilis could be a novel means to fortify fermented products with peptides and γ-PGA.
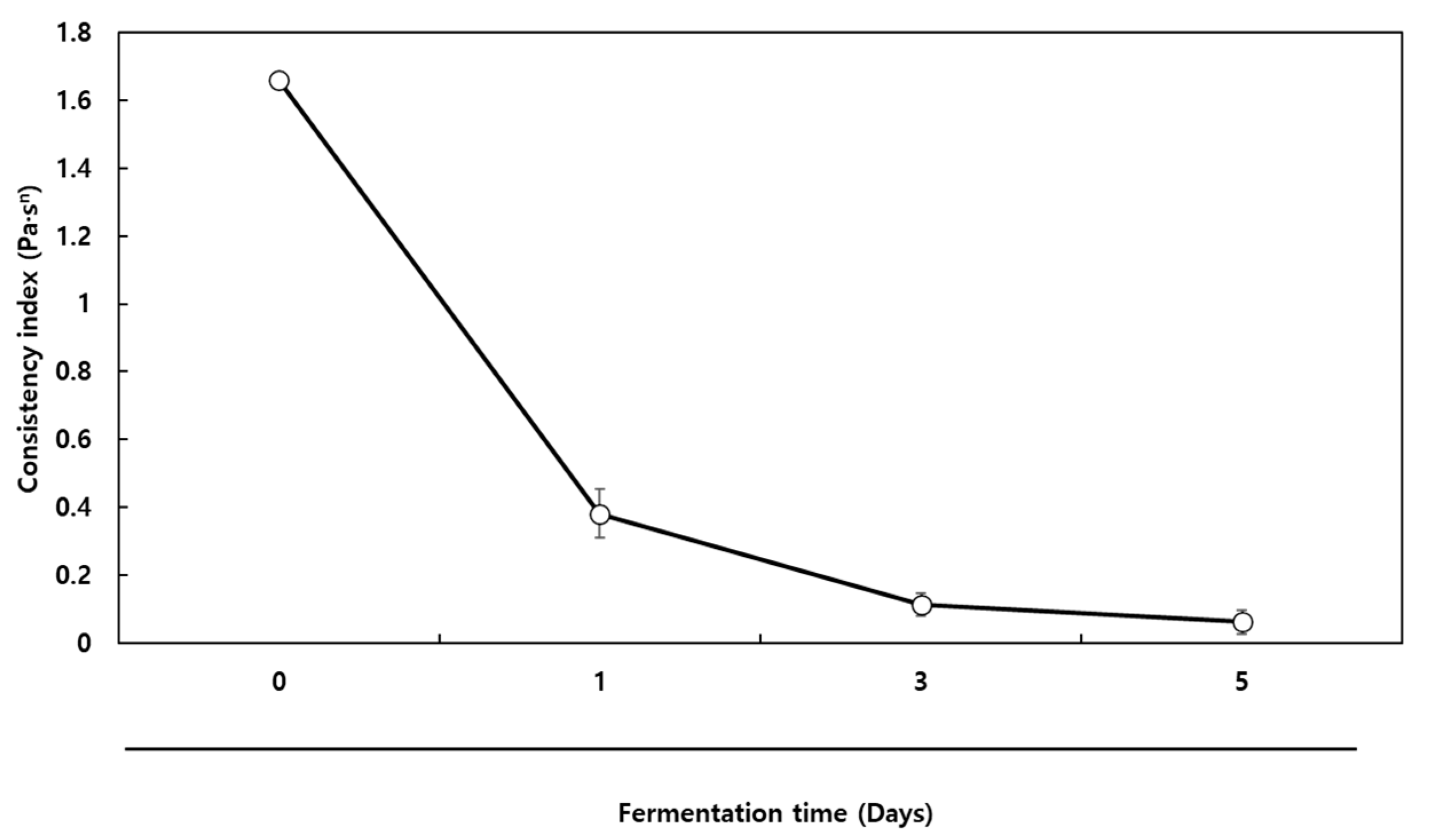
3.7. Consistency Index of the Co-Fermented Milk
The milk co-fermented by
B. subtilis and
L. plantarum was fortified with probiotics, peptides, and γ-PGA. γ-PGA may act as a prebiotic according to a study on its effects on the gut microbiota of mice [
26]. In addition, fermentation of milk with
B. subtilis improved the consistency of the fermented milk. Consistency is an important rheological property because it affects taste, mouthfeel, and whey separation. Generally, hydrocolloids such as gums are applied to fermented milk to enhance its consistency. The addition of prebiotics such as galactooligosaccharide and inulin to Greek yogurt has resulted in enhanced rheological properties, providing a more consistent, elastic, viscous, and firm product [
27].
Thus, alkaline fermentation of milk with
B. subtilis in the presence of MSG could be a potential means to naturally enhance the consistency of fermented products. As shown in
Figure 7, the milk fermented with
B. subtilis for 1 day showed a consistency index of 1.66 Pa·S
n, but serial co-fermentation with
L. plantarum for 1 day decreased to 0.38 Pa·S
n. After 5 days of co-fermentation, the consistency index decreased to 0.06 Pa·S
n (
Figure 7), indicating that the consistency of mucilage was greatly dependent upon the acidic environment, and increased acidity resulted in lowered consistency. γ-PGA is a water-soluble anionic polypeptide, which undergoes structural changes related to the degree of ionization of its γ-carboxyl group. For these reasons, the consistency and molecular weight of γ-PGA can greatly decrease at a low pH [
28]. Although the co-fermented milk had the lowest consistency, it was fortified with bioactive compounds, which may be considered a trade-off.
3.8. GABA Production in Co-Fermented Milk
The glutamic acid and GABA in the co-fermented milk were quantified by UPLC/MS (
Table 1). In the optimal milk broth, which contained 3.0% MSG, the initial glutamic acid content was 26.25 mg/mL, but it decreased to 18.85 mg/mL after the first alkaline fermentation. During the co-fermentation with additional supplementation of 1.5% MSG, the glutamic acid content decreased from 3.05% to 0.01%. The final co-fermented milk contained 2.61% GABA. Thus, serial co-fermentation of milk in the presence of MSG enriched the fermented product with GABA and probiotics.
3.9. Volatile Composition of Co-Fermented Milk
Generally, alkaline fermentation of protein-rich ingredients by Bacillus spp. gives off an unpleasant odor, which decreases consumer acceptance, thus hindering the production of fermented foods using these bacteria. The alkaline fermentation of milk with B. subtilis performed in this study is a special approach in dairy fermentation technology as B. subtilis is generally used in the production of fermented soybean products like chungkookjang but not dairy making. In addition, serial co-fermentation using L. plantarum isolated from fermented vegetables is a novel process since this bacterium is not ordinarily used to ferment milk.
The alkaline fermentation of milk by
B. subtilis HA generated a unique aroma similar to that of aged cheese. GC/MS analysis of the co-fermented milk revealed the presence of three major compounds, namely hexanoic acid, 2,3-butanediol, and acetoin (
Table 2). Hexanoic acid, also known as caproic acid, is a colorless oily liquid with a fatty, cheesy, and waxy aroma. Acetoin is the last synthetic intermediate for 2,3-butanediol production, and metabolic conversion of acetoin to 2,3-butanediol is reversible in most bacteria [
29,
30]. Furthermore, acetoin can be converted to buttery flavor compounds by oxidation. Milk that has been fermented generally contains diacetyl, produced by dehydrogenation of 2,3-butanediol, and is an intermediary in producing odorants like hexanoic acid, acetoin, 2-heptanone, and 2-nonanone [
30].
4. Conclusions
We developed a fermented dairy product enriched with bioactive compounds by a stepwise co-fermentation using B. subtilis HA and L. plantarum EJ2014. MSG was added as a precursor to producing a higher amount of both γ-PGA and GABA. After the first alkaline fermentation, milk supplemented with 3% MSG and 2.5% glucose contained γ-PGA and was highly viscous due to the incomplete bioconversion of MSG. During the co-fermentation with L. plantarum EJ2014, MSG was completely bioconverted, resulting in higher production of GABA (up to 2.61%). The first alkaline fermentation of the milk provided protease activity, generating peptides with low molecular weight through the digestion of milk proteins. The co-fermented milk was effectively fortified with bioactive compounds such as γ-PGA, GABA, and peptides, as well as probiotics. In addition, the co-fermented milk exhibited a unique aroma similar to that of aged cheese. This co-fermented milk could be used as an ingredient in producing novel healthy products. In conclusion, the serial co-fermentation of milk in the presence of MSG could be a novel strategy to develop multi-functional ingredients and enhance flavor.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}