
Bacterial Diversity and Dynamics during Spontaneous Cheese Whey Fermentation at Different Temperatures

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Whey Samples and Whey Fermentation
2.2. pH Measurement and Lactic Acid Content
2.3. Mesophilic and Lactic Acid Bacteria Enumeration
2.4. DNA Extraction and Amplification
2.5. Library Construction and Illumina Sequencing
2.6. Illumina Data Analysis and Sequence Identification
3. Results and Discussion
3.1. Viable Bacterial Count in Spontaneously Fermented Whey
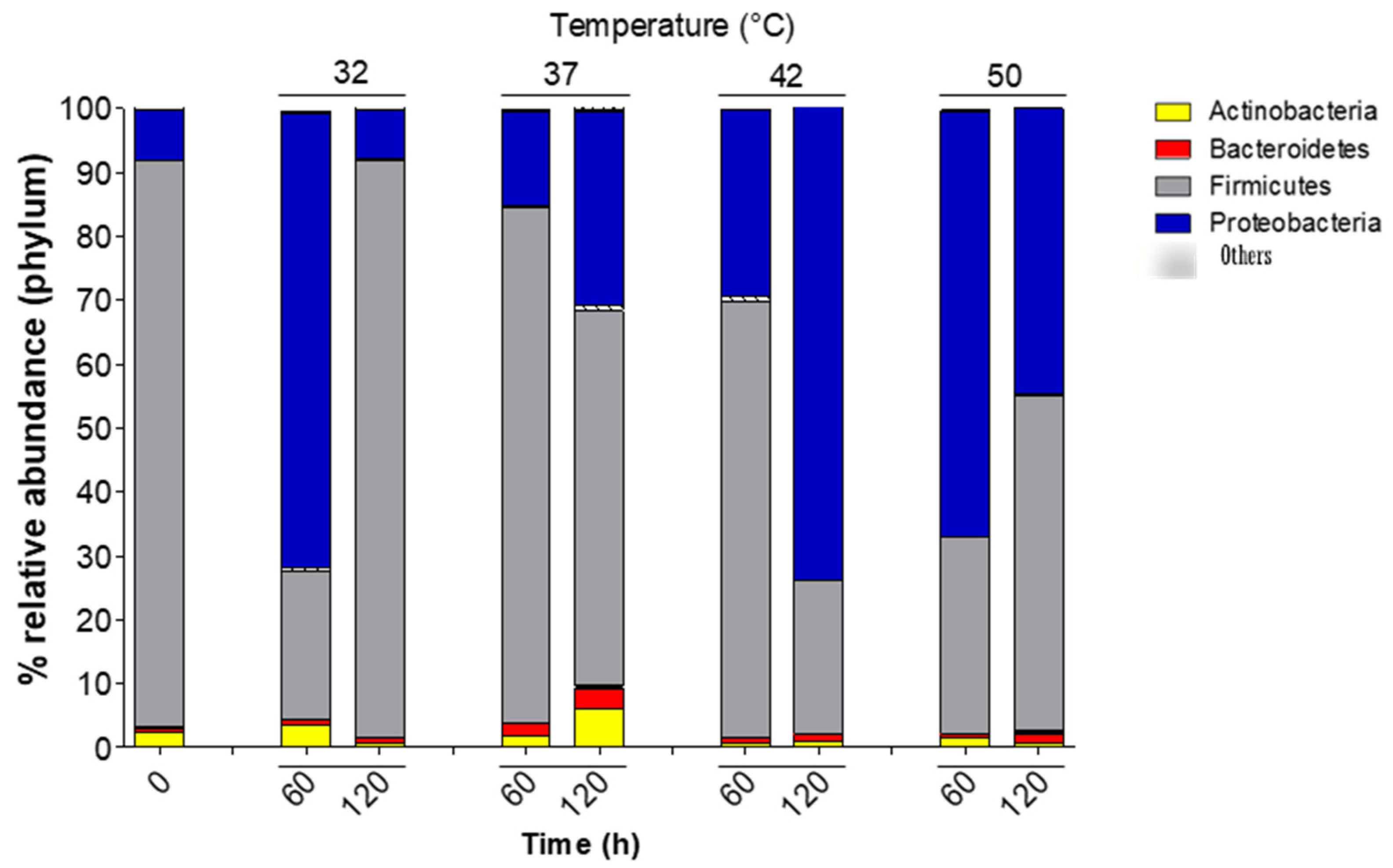
3.2. Bacterial Phylum Diversity during Spontaneous Whey Fermentation
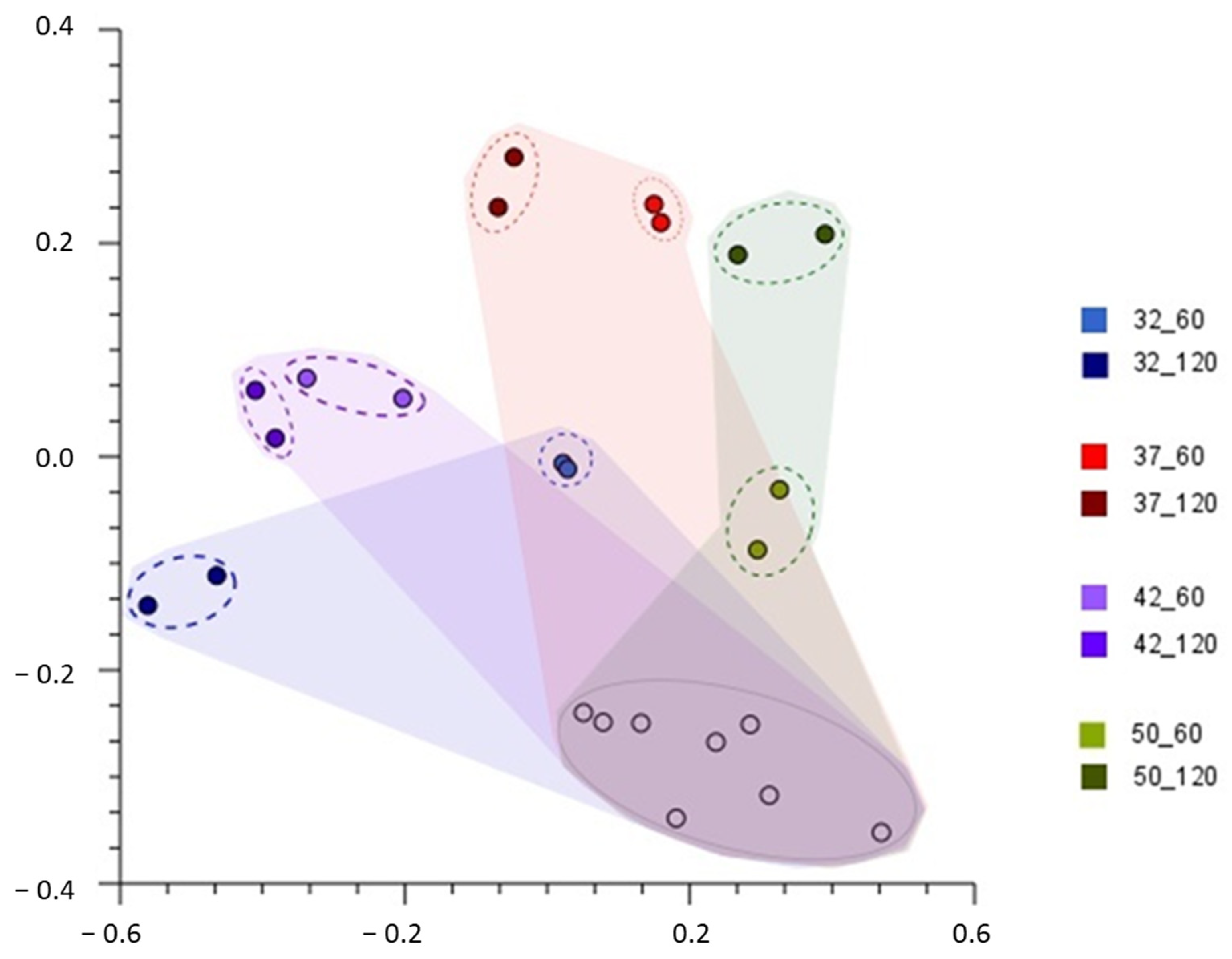
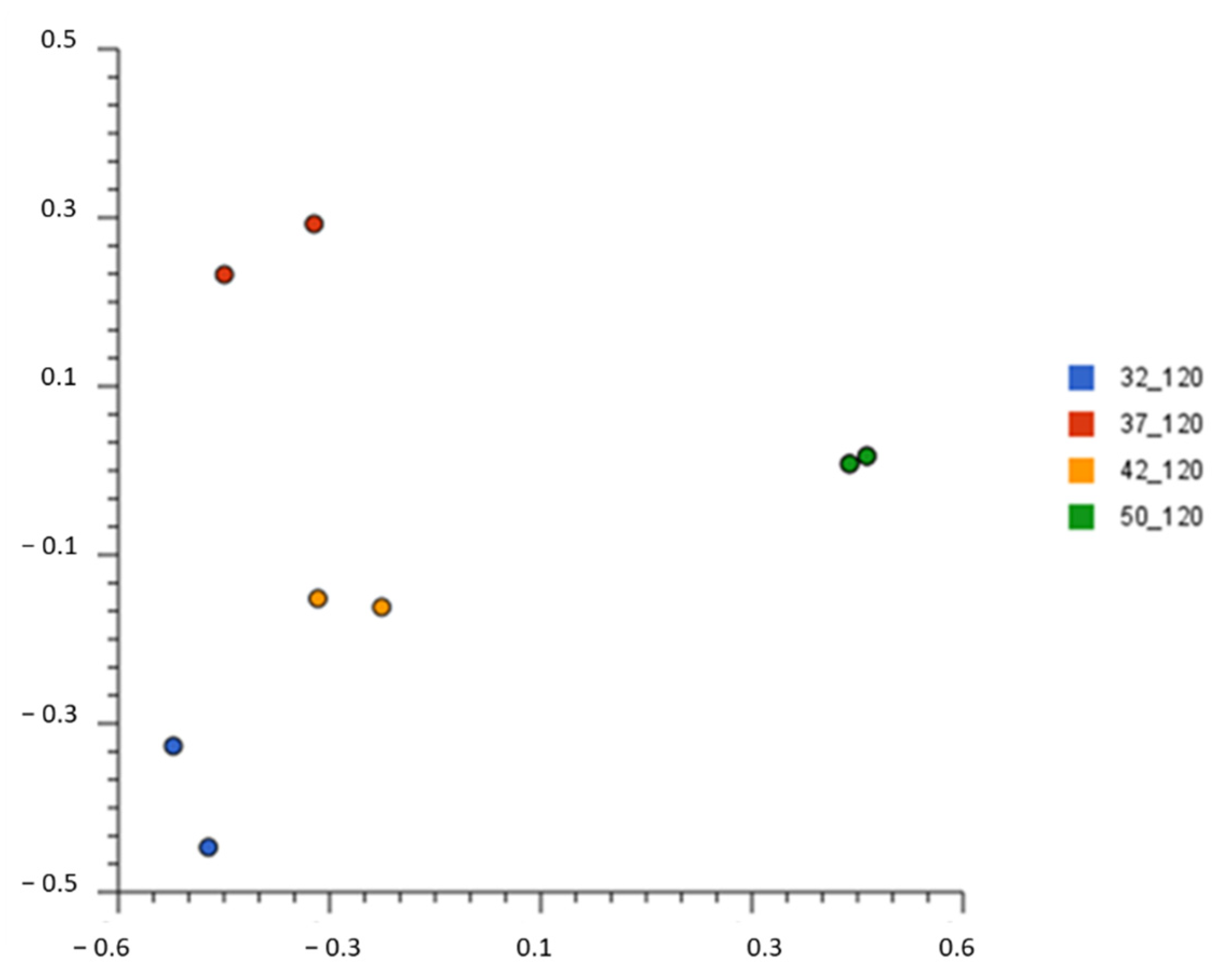
3.3. Bacterial Diversity during Whey Fermentation at Genus Level
3.4. Species Bacterial Diversity in Fermented Whey
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazorra-Manzano, M.A.; Moreno-Hernández, J.M. Properties and options for the valorization of whey from the artisanal cheese industry. CienciaUAT 2019, 14, 133–144. [Google Scholar]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous food fermentations and potential risks for human health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevas-González, P.F.; Heredia-Castro, P.Y.; Méndez-Romero, J.I.; Hernández-Mendoza, A.; Reyes-Díaz, R.; Vallejo-Cordoba, B.; González-Córdova, A.F. Artisanal Sonoran cheese (Cocido cheese): An exploration of its production process, chemical composition and microbiological quality. J. Sci. Food Agric. 2017, 97, 4459–4466. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; González-Velázquez, D.A.; Torres-Llanez, M.J.; Martínez-Porchas, M.; García-Sifuentes, C.O.; González-Córdova, A.F.; Vallejo-Córdoba, B. Cheese whey fermentation by its native microbiota: Proteolysis and bioactive peptides release with ACE-inhibitory activity. Fermentation 2020, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent advances in lactic acid production by microbial fermentation processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef]
- Martín-del-Campo, S.T.; Martínez-Basilio, P.C.; Sepúlveda-Álvarez, J.C.; Gutiérrez-Melchor, S.E.; Galindo-Peña, K.D.; Lara-Domínguez, A.L.; Cardador-Martínez, A. Production of antioxidant and ACEI peptides from cheese whey discarded from Mexican white cheese production. Antioxidants 2019, 8, 158. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Guo, J.; Huang, A. Study on bacterial diversity in traditional sour whey of Yunnan province. J. Food Process. Preserv. 2018, 42, e13553. [Google Scholar] [CrossRef]
- Torres-Llanez, M.J.; Vallejo-Cordoba, B.; Díaz-Cinco, M.E.; Mazorra-Manzano, M.A.; González-Córdova, A.F. Characterization of the natural microflora of artisanal Mexican Fresco cheese. Food Control 2006, 17, 683–690. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- De Candia, S.; De Angelis, M.; Dunlea, E.; Minervini, F.; McSweeney, P.; Faccia, M.; Gobbetti, M. Molecular identification and typing of natural whey starter cultures and microbiological and compositional properties of related traditional Mozzarella cheeses. Int. J. Food Microbiol. 2007, 119, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. Molecular approaches to analyzing the microbial composition of raw milk and raw milk cheese. Int. J. Food Microbiol. 2011, 150, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Delbés, C.; Ali-Mandjee, L.; Montel, M.-C. Monitoring bacterial communities in raw milk and cheese by culture-dependent and -independent 16S rRNA gene-based analyses. Appl. Environ. Microbiol. 2007, 73, 1882–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taale, E.; Savadogo, A.; Zongo, C.; Ilboudo, A.J.; Traore, A.S. Bioactive molecules from bacteria strains: Case of bacteriocins producing bacteria isolated from foods. Curr. Res. Microbiol. Biotechnol. 2013, 1, 80–88. [Google Scholar]
- Aldrete-Tapia, A.; Escobar-Ramírez, M.C.; Tamplin, M.L.; Hernández-Iturriaga, M. High-throughput sequencing of microbial communities in Poro cheese, an artisanal Mexican cheese. Food Microbiol. 2014, 44, 136–141. [Google Scholar] [CrossRef]
- Lusk, T.S.; Ottesen, A.R.; White, J.R.; Allard, M.W.; Brown, E.W.; Kase, J.A. Characterization of microflora in Latin-style cheeses by next-generation sequencing technology. BMC Microbiol. 2012, 12, 254. [Google Scholar] [CrossRef] [Green Version]
- Fuhrman, J.A. Microbial community structure and its functional implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef]
- Ryssel, M.; Johansen, P.; Al-Soud, W.A.; Sørensen, S.; Arneborg, N.; Jespersen, L. Microbial diversity and dynamics throughout manufacturing and ripening of surface ripened semi-hard Danish Danbo cheeses investigated by culture-independent techniques. Int. J. Food Microbiol. 2015, 215, 124–130. [Google Scholar] [CrossRef]
- Motato, K.E.; Milani, C.; Ventura, M.; Valencia, F.E.; Ruas-Madiedo, P.; Delgado, S. Bacterial diversity of the Colombian fermented milk “Suero Costeño” assessed by culturing and and DGGE analysis of 16S rRNA gene amplicons. Food Microbiol. 2017, 68, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. High-throughput sequencing for detection of subpopulations of bacteria not previously associated with artisanal cheeses. Appl. Environ. Microbiol. 2012, 78, 5717–5723. [Google Scholar] [CrossRef] [Green Version]
- Pescuma, M.; Hébert, E.M.; Mozzi, F.; Font de Valdez, G. Whey fermentation by thermophilic lactic acid bacteria: Evolution of carbohydrates and protein content. Food Microbiol. 2008, 25, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Vera-Peña, M.Y.; Rodriguez-Rodriguez, W.L. Effect of pH on the growth of three lactic acid bacteria strains isolated from sour cream. Univ. Sci. 2020, 25, 341–358. [Google Scholar]
- Abol-Fotouh, D.M.; Bayoumi, R.A.; Hassan, M.A. Production of thermoalkaliphilic lipase from Geobacillus thermoleovorans DA2 and application in leather industry. Enzym. Res. 2016, 2016, 9034364. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Fermented Whey (Treatment) | Bacterial Counts (log CFU/mL) | |||||
|---|---|---|---|---|---|---|
| Temperature | Time (h) | Mesophilic (PCA) | Streptococci (M17) | Lactobacilli (MRS) | pH | Lactic Acid (g/L) |
| 37 °C | 0 | 5.29 | 5.08 | 4.99 | 6.49 | 0.90 |
| 60 | 7.76 | 6.87 | 7.99 | 3.65 | 7.42 | |
| 120 | 6.65 | 7.37 | 6.21 | 3.35 | 12.33 | |
| 42 °C | 0 | 5.23 | 5.08 | 4.74 | 6.56 | 0.89 |
| 60 | 7.01 | 5.36 | 7.27 | 3.35 | 9.38 | |
| 120 | 5.12 | 6.63 | 5.43 | 3.30 | 12.83 | |
| Fermented Whey (Treatment) | Classified Reads | Phylum | Class | Order | Family | Genus | Species |
|---|---|---|---|---|---|---|---|
| % | |||||||
| 32 °C_0 h (Control) | 89.95 | 84.19 | 81.05 | 77.93 | 71.59 | 54.68 | 35.36 |
| 32 °C_60 h | 93.45 | 86.27 | 82.88 | 74.50 | 70.56 | 52.09 | 18.48 |
| 37 °C_60 h | 92.12 | 85.76 | 82.63 | 77.99 | 71.23 | 50.63 | 35.55 |
| 42 °C_60 h | 91.82 | 85.96 | 84.02 | 77.36 | 74.90 | 38.38 | 7.73 |
| 50 °C_60 h | 89.42 | 85.17 | 77.84 | 75.08 | 66.35 | 49.68 | 29.33 |
| 32 °C_120 h | 92.28 | 87.66 | 84.39 | 81.44 | 78.41 | 55.18 | 40.39 |
| 37 °C_120 h | 81.87 | 74.67 | 69.80 | 64.39 | 55.97 | 39.56 | 21.87 |
| 42 °C_120 h | 82.59 | 76.82 | 74.26 | 62.45 | 60.73 | 39.22 | 30.24 |
| 50 °C_120 h | 84.40 | 74.93 | 71.31 | 69.36 | 57.38 | 51.04 | 40.04 |
| Genus of Lactic Acid Bacterium | Genus of Non-Fermentative Bacterium | ||||
|---|---|---|---|---|---|
| Staphylococcus | 9.5 | Salmonella | 3.7 | ||
| Streptococcus | 17.2 | Alteromonas | 8.0 | Macrococcus | 3.5 |
| Lactococcus | 11.2 | Geobacillus | 7.7 | Pantoea | 3.5 |
| Lactobacillus | 8.0 | Acinetobacter | 4.5 | Erwinia | 3.2 |
| Tetragenococcus | 5.5 | Arcanobacterium | 4.2 | Bacillus | 3.0 |
| Ketogulonicigenium | 3.7 | Nocardioides | 3.0 | ||
| Spontaneous Fermented Whey | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 60 h | 120 h | ||||||||
| Temperature (°C) | |||||||||
| Bacteria | Fresh Whey | 32 | 37 | 42 | 50 | 32 | 37 | 42 | 50 |
| Alteromonas macleodii | |||||||||
| Streptococcus agalactiae | |||||||||
| Tetragenococcus halophilus | |||||||||
| Arcanobacterium haemolyticum | |||||||||
| Geobacillus thermoleovorans | |||||||||
| Lactococcus lactis | |||||||||
| Staphylococcus aureus | |||||||||
| Rhodopirellula baltica | |||||||||
| Sulfurimonas autotrophica | |||||||||
| Enterococcus casseliflavus | |||||||||
| Pantoea vagans | |||||||||
| Brachybacterium faecium | |||||||||
| Streptococcus salivarius | |||||||||
| Streptococcus thermophilus | |||||||||
| Cytophaga hutchinsonii | |||||||||
| Macrococcus caseolyticus | |||||||||
| Ketogulonicigenium vulgare | |||||||||
| Bacillus subtilis | |||||||||
| Lactobacillus sakei | |||||||||
| Deinococcus gobiensis | |||||||||
| Erwinia pyrifoliae | |||||||||
| Bacillus cereus | |||||||||
| Jannaschia sp. CCS1 | |||||||||
| Lactobacillus salivarius | |||||||||
| Lactobacillus acidophilus | |||||||||
| Lactobacillus reuteri | |||||||||
| Marivirga tractuosa | |||||||||
| Plautia stali symbiont | |||||||||
| Microbacterium testaceum | |||||||||
| Sphingobacterium sp.21 | |||||||||
| Salmonella enterica | |||||||||
| Acinetobacter baumannii | |||||||||
| Streptococcus mutans | |||||||||
| Lactobacillus helveticus | |||||||||
| Riemerella anatipestifer | |||||||||
| Lactococcus garvieae | |||||||||
| Croceibacter atlanticus | |||||||||
| Ilumatobacter coccineus | |||||||||
| Dinoroseobacter shibae | |||||||||
| Escherichia coli | |||||||||
| Salmonella bongori | |||||||||
| Erysipelothrix rhusiopathiae | |||||||||
| Ruegeria pomeroyi | |||||||||
| Secondary endosymbiont of Heteropsylla cubana | |||||||||
| Moraxella catarrhalis | |||||||||
| Kucuria rhizophila | |||||||||
| Leuconostoc mesenteroides | |||||||||
| Dickeya dadantii | |||||||||
| Candidatus Moranella endobia | |||||||||
Relative abundance (%)  . .
| |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; Martínez-Porchas, M.; Ramírez-Suárez, J.C.; García-Sifuentes, C.O.; Torres-Llanez, M.J.; González-Córdova, A.F.; Hernández-Mendoza, A.; Vallejo-Cordoba, B. Bacterial Diversity and Dynamics during Spontaneous Cheese Whey Fermentation at Different Temperatures. Fermentation 2022, 8, 342. https://doi.org/10.3390/fermentation8070342
Mazorra-Manzano MA, Robles-Porchas GR, Martínez-Porchas M, Ramírez-Suárez JC, García-Sifuentes CO, Torres-Llanez MJ, González-Córdova AF, Hernández-Mendoza A, Vallejo-Cordoba B. Bacterial Diversity and Dynamics during Spontaneous Cheese Whey Fermentation at Different Temperatures. Fermentation. 2022; 8(7):342. https://doi.org/10.3390/fermentation8070342
Chicago/Turabian StyleMazorra-Manzano, Miguel A., Glen R. Robles-Porchas, Marcel Martínez-Porchas, Juan C. Ramírez-Suárez, Celia O. García-Sifuentes, María J. Torres-Llanez, Aarón F. González-Córdova, Adrián Hernández-Mendoza, and Belinda Vallejo-Cordoba. 2022. "Bacterial Diversity and Dynamics during Spontaneous Cheese Whey Fermentation at Different Temperatures" Fermentation 8, no. 7: 342. https://doi.org/10.3390/fermentation8070342