
Recombinant Protein Production in E. coli Using the phoA Expression System


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Product
2.2. Bioreactor Cultivations
2.2.1. Strain Characterization
T7lac-Based Expression (pET Cultivations)
phoA-Based Expression (pAT Cultivations)
Sampling
2.2.2. Characterization of the pAT System
- Ft feed rate [g/h];
- F0 initial feed rate [g/h];
- µ specific growth rate [1/h];
- t cultivation time [h].
- F0 initial feed rate at time point 0 [g/h];
- µ specific growth rate [1/h];
- x0 biomass conc. at time point 0 [g/L];
- V0 culture volume at time point 0 [L];
- cs,Feed glucose conc. in feed medium [g/L];
- YX/S biomass yield on glucose [g/g].
2.3. Analytics
2.3.1. Sample Preparation for Product Analysis
2.3.2. Soluble Product Quantification by Affinity HPLC
2.3.3. Product IB Quantification by Size Exclusion HPLC
2.3.4. Investigation of PO4 Quantification Methods
ICP-OES
IC
Colorimetric Assay Kit
Cedex Bio HT Analyzer
3. Results
3.1. Characterization of T7lac-Based Fab Production (pET)
3.1.1. Strain Physiology
3.1.2. Fab Productivity
3.2. Characterization of phoA-Based Fab Production (pAT)
3.2.1. Impact of Cultivation Conditions on the Overall Cultivation Time
3.2.2. Strain Physiology
3.2.3. Fab Productivity
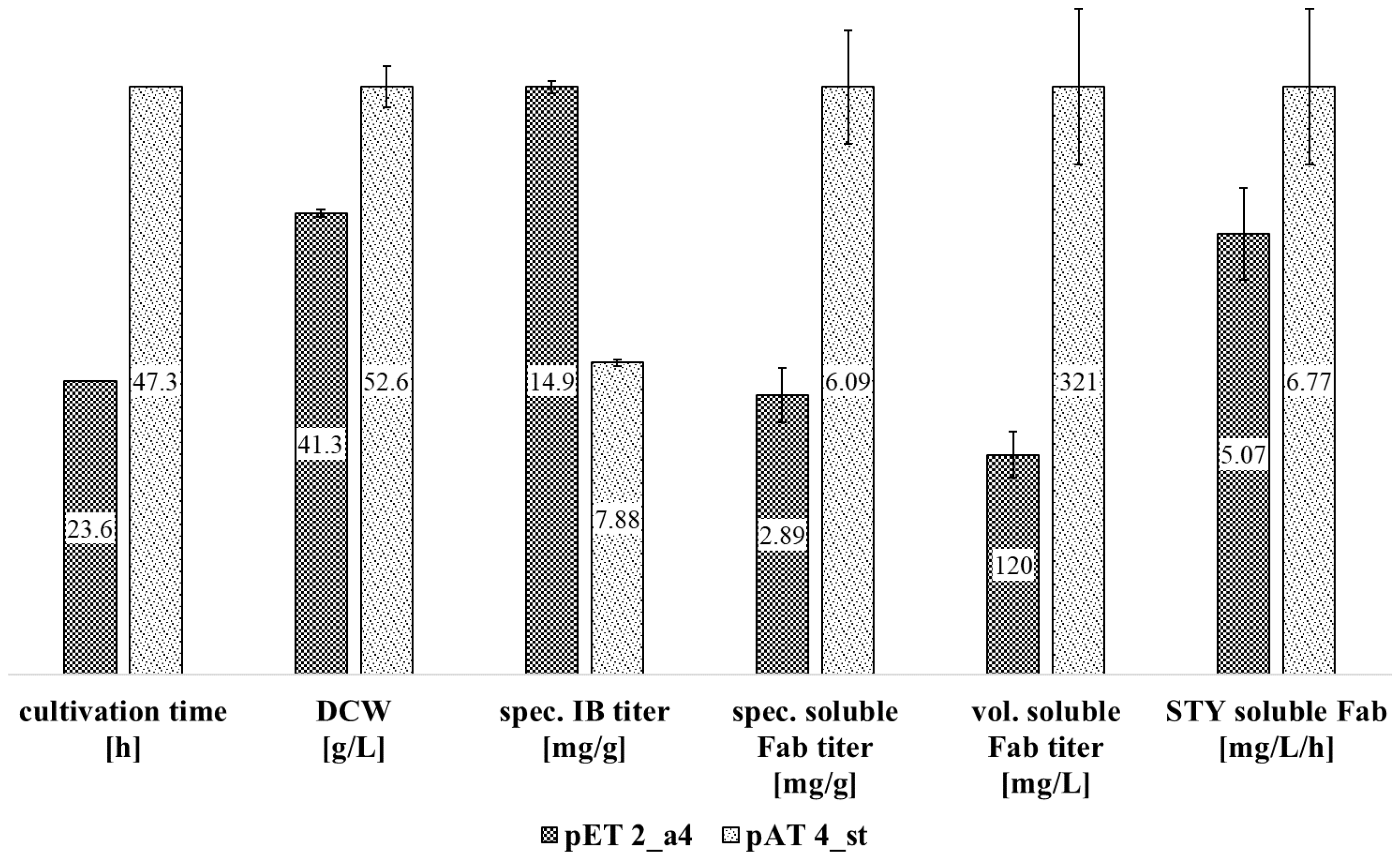
3.3. Direct Comparison of T7lac- and phoA-Based Fab Production
3.4. Detailed Characterization of the pAT System
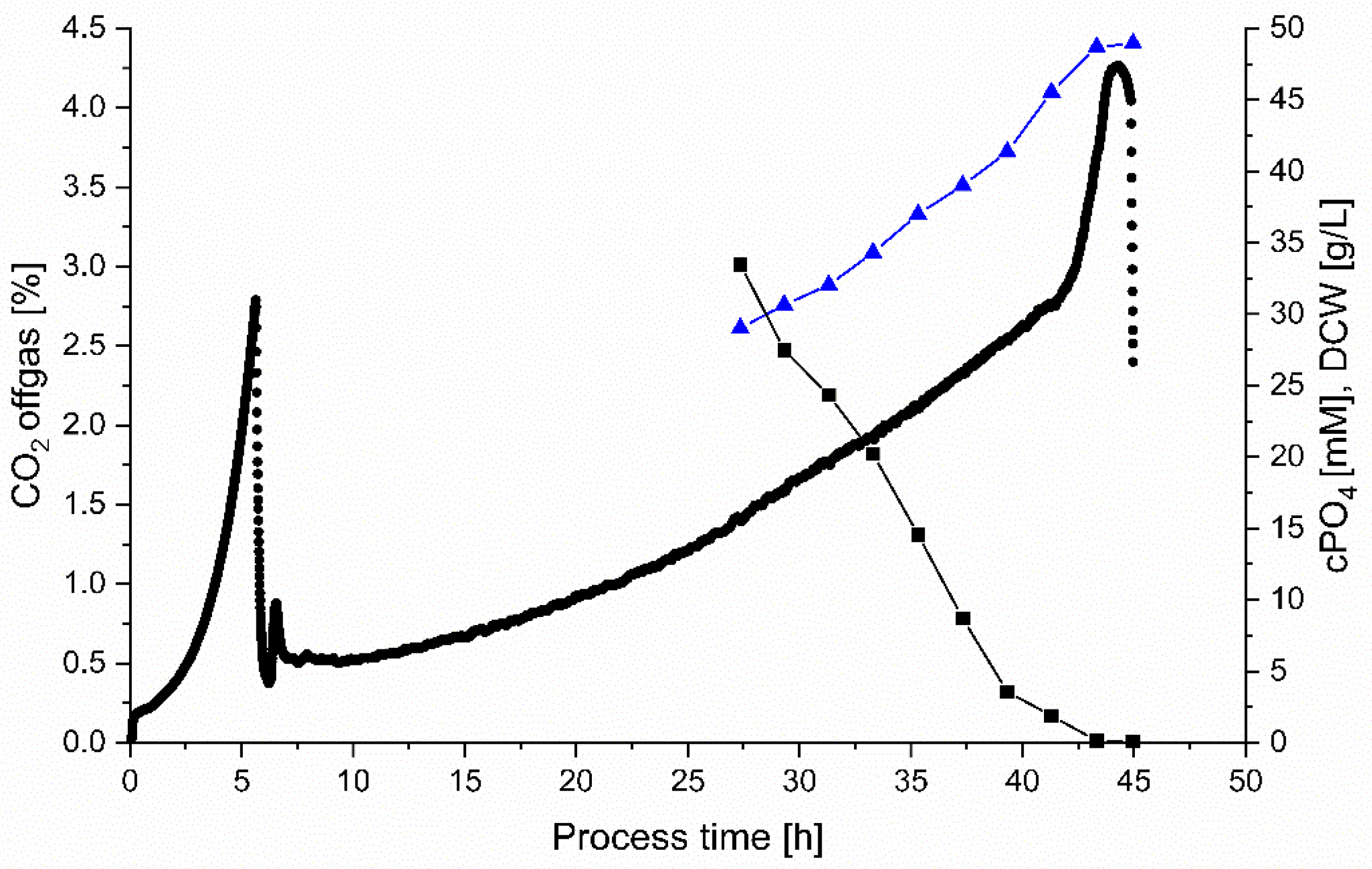
3.4.1. PO4 Monitoring
ICP-OES
IC
Phosphate Colorimetric Assay Kit
Cedex Bio HT
3.4.2. Impact of PO4 Conditions on Strain Physiology
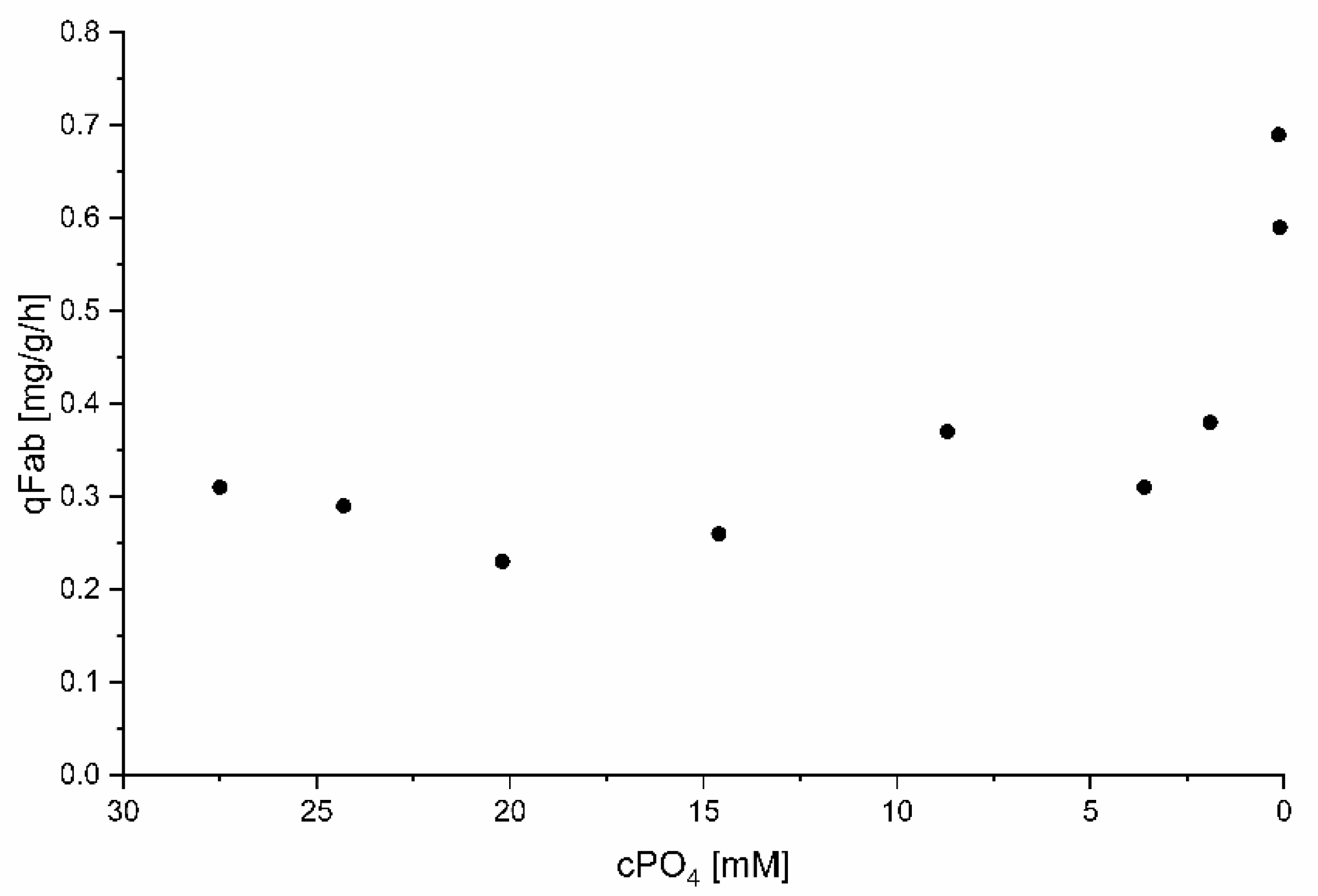
3.4.3. Impact of PO4 Conditions on Fab Productivity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holt, L.J.; Herring, C.; Jespers, L.S.; Woolven, B.P.; Tomlinson, I.M. Domain antibodies: Proteins for therapy. Trends Biotechnol. 2003, 21, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, G.; Gruvegard, M.; Van Alstine, J.M. Antibody Fragments and Their Purification by Protein L Affinity Chromatography. Antibodies 2015, 4, 259–277. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Zhao, M.; Cagliero, C.; Jiang, H.; Xie, Y.; Zhu, J.; Yang, H.; Zhang, M.; Zheng, Y.; Yuan, Y.; et al. A general platform for efficient extracellular expression and purification of Fab from Escherichia coli. Appl. Microbiol. Biotechnol. 2019, 103, 3341–3353. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadokura, H.; Katzen, F.; Beckwith, J. Protein disulfide bond formation in prokaryotes. Annu. Rev. Biochem. 2003, 72, 111–135. [Google Scholar] [CrossRef] [PubMed]
- Gundinger, T.; Spadiut, O. A comparative approach to recombinantly produce the plant enzyme horseradish peroxidase in Escherichia coli. J. Biotechnol. 2017, 248, 15–24. [Google Scholar] [CrossRef]
- Skerra, A.; Pluckthun, A. Assembly of a functional immunoglobulin Fv fragment in Escherichia coli. Science 1988, 240, 1038–1041. [Google Scholar] [CrossRef]
- Ukkonen, K.; Veijola, J.; Vasala, A.; Neubauer, P. Effect of culture medium, host strain and oxygen transfer on recombinant Fab antibody fragment yield and leakage to medium in shaken E. coli cultures. Microb. Cell Fact. 2013, 12, 73. [Google Scholar] [CrossRef] [Green Version]
- Carter, P.; Kelley, R.F.; Rodrigues, M.L.; Snedecor, B.; Covarrubias, M.; Velligan, M.D.; Wong, W.L.; Rowland, A.M.; Kotts, C.E.; Carver, M.E.; et al. High level Escherichia coli expression and production of a bivalent humanized antibody fragment. Biotechnology 1992, 10, 163–167. [Google Scholar] [CrossRef]
- Better, M.; Chang, C.P.; Robinson, R.R.; Horwitz, A.H. Escherichia coli secretion of an active chimeric antibody fragment. Science 1988, 240, 1041–1043. [Google Scholar] [CrossRef]
- Ellis, M.; Patel, P.; Edon, M.; Ramage, W.; Dickinson, R.; Humphreys, D.P. Development of a high yielding E. coli periplasmic expression system for the production of humanized Fab’ fragments. Biotechnol. Prog. 2017, 33, 212–220. [Google Scholar] [CrossRef]
- Shibui, T.; Munakata, K.; Matsumoto, R.; Ohta, K.; Matsushima, R.; Morimoto, Y.; Nagahari, K. High-level production and secretion of a mouse-human chimeric Fab fragment with specificity to human carcino embryonic antigen in Escherichia coli. Appl. Microbiol. Biotechnol. 1993, 38, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Kulmala, A.; Huovinen, T.; Lamminmaki, U. Effect of DNA sequence of Fab fragment on yield characteristics and cell growth of E. coli. Sci. Rep. 2017, 7, 3796. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.; Ahluwalia, K.; Bohmann, D.J.; Giang, H.M.; Schwimmer, L.J.; Issafras, H.; Reddy, N.B.; Chan, C.; Horwitz, A.H.; Takeuchi, T. Enhancement of antibody fragment secretion into the Escherichia coli periplasm by co-expression with the peptidyl prolyl isomerase, FkpA, in the cytoplasm. J. Immunol. Methods 2013, 394, 10–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, B.; Renshaw, M.W.; Autote, K.; Smith, L.M.; Calveley, P.; Bowdish, K.S.; Frederickson, S. A step-wise approach significantly enhances protein yield of a rationally-designed agonist antibody fragment in E. coli. Protein Expr. Purif. 2008, 59, 55–63. [Google Scholar] [CrossRef]
- Frenzel, A.; Hust, M.; Schirrmann, T. Expression of recombinant antibodies. Front. Immunol. 2013, 4, 217. [Google Scholar] [CrossRef] [Green Version]
- Rosano, G.A.-O.; Morales, E.S.; Ceccarelli, E.A.-O. New tools for recombinant protein production in Escherichia coli: A 5-year update. Protein Sci. 2019, 28, 1412–1422. [Google Scholar] [CrossRef]
- Ukkonen, K.; Vasala, A.; Ojamo, H.; Neubauer, P. High-yield production of biologically active recombinant protein in shake flask culture by combination of enzyme-based glucose delivery and increased oxygen transfer. Microb. Cell Fact. 2011, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, A.; Kelley, L.L.; Momany, C. Optimization of a mouse recombinant antibody fragment for efficient production from Escherichia coli. Protein Expr. Purif. 2007, 52, 219–229. [Google Scholar] [CrossRef]
- Friedrich, L.; Stangl, S.; Hahne, H.; Kuster, B.; Kohler, P.; Multhoff, G.; Skerra, A. Bacterial production and functional characterization of the Fab fragment of the murine IgG1/lambda monoclonal antibody cmHsp70.1, a reagent for tumour diagnostics. Protein Eng. Des. Sel. PEDS 2010, 23, 161–168. [Google Scholar] [CrossRef]
- Gupta, S.K.; Shukla, P. Microbial platform technology for recombinant antibody fragment production: A review. Crit. Rev. Microbiol. 2017, 43, 31–42. [Google Scholar] [CrossRef]
- De Palma, A. Advances in protein expression. Genet. Eng. Biotechnol. News 2014, 34, 24–25, 27. [Google Scholar] [CrossRef]
- Pack, P.; Kujau, M.; Schroeckh, V.; Knupfer, U.; Wenderoth, R.; Riesenberg, D.; Pluckthun, A. Improved bivalent miniantibodies, with identical avidity as whole antibodies, produced by high cell density fermentation of Escherichia coli. Biotechnology 1993, 11, 1271–1277. [Google Scholar] [CrossRef]
- Shin, P.K.; Seo, J.H. Analysis of E. coli phoA-lacZ fusion gene expression inserted into a multicopy plasmid and host cell’s chromosome. Biotechnol. Bioeng. 1990, 36, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Morinaga, Y.; Tsuchiya, M.; Ikemura, H.; Inouyea, M. Control of Folding of Proteins Secreted by a High Expression Secretion Vector, pIN-III-ompA: 16-Fold Increase in Production of Active Subtilisin E in Escherichia coli. Biotechnology 1988, 6, 948. [Google Scholar] [CrossRef]
- Harrison, J.S.; Keshavarz-Moore, E. Production of antibody fragments in Escherichia coli. Ann. N. Y. Acad. Sci. 1996, 782, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Baneyx, F. Recombinant protein expression in Escherichia coli. Curr. Opin. Biotechnol. 1999, 10, 411–421. [Google Scholar] [CrossRef]
- Mergulhao, F.J.; Monteiro, G.A. Secretion capacity limitations of the Sec pathway in Escherichia coli. J. Microb. Biotechnol. 2004, 14, 128–133. [Google Scholar]
- Zhang, L.; Cao, Y.; Liu, M.; Chen, X.; Xiang, Q.; Tian, J. Functional recombinant single-chain variable fragment antibody against Agkistrodon acutus venom. Exp. Med 2019, 17, 3768–3774. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Xu, L.; Li, B.; Zhong, F.; Liu, X.; Zhang, X. Canine Parvovirus is diagnosed and neutralized by chicken IgY-scFv generated against the virus capsid protein. Vet. Res. 2020, 51, 110. [Google Scholar] [CrossRef]
- Sharma, S.K.; Suresh, M.R.; Wuest, F.R. Improved soluble expression of a single-chain antibody fragment in E. coli for targeting CA125 in epithelial ovarian cancer. Protein Expr. Purif. 2014, 102, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Wechselberger, P.; Sagmeister, P.; Herwig, C. Real-time estimation of biomass and specific growth rate in physiologically variable recombinant fed-batch processes. Bioprocess Biosyst. Eng. 2013, 36, 1205–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLisa, M.P.; Li, J.; Rao, G.; Weigand, W.A.; Bentley, W.E. Monitoring GFP-operon fusion protein expression during high cell density cultivation of Escherichia coli using an on-line optical sensor. Biotechnol. Bioeng. 1999, 65, 54–64. [Google Scholar] [CrossRef]
- Wurm, D.J.; Veiter, L.; Ulonska, S.; Eggenreich, B.; Herwig, C.; Spadiut, O. The E. coli pET expression system revisited-mechanistic correlation between glucose and lactose uptake. Appl. Microbiol. Biotechnol. 2016, 100, 8721–8729. [Google Scholar] [CrossRef] [Green Version]
- Marschall, L.; Sagmeister, P.; Herwig, C. Tunable recombinant protein expression in E. coli: Enabler for continuous processing? Appl. Microbiol. Biotechnol. 2016, 100, 5719–5728. [Google Scholar] [CrossRef] [Green Version]
- Larentis, A.L.; Nicolau, J.F.M.Q.; dos Santos Esteves, G.; Vareschini, D.T.; de Almeida, F.V.R.; dos Reis, M.G.; Galler, R.; Medeiros, M.A. Evaluation of pre-induction temperature, cell growth at induction and IPTG concentration on the expression of a leptospiral protein in E. coli using shaking flasks and microbioreactor. BMC Res. Notes 2014, 7, 671. [Google Scholar] [CrossRef] [Green Version]
- Gundinger, T.; Pansy, A.; Spadiut, O. A sensitive and robust HPLC method to quantify recombinant antibody fragments in E. coli crude cell lysate. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1083, 242–248. [Google Scholar] [CrossRef]
- Wurm, D.J.; Quehenberger, J.; Mildner, J.; Eggenreich, B.; Slouka, C.; Schwaighofer, A.; Wieland, K.; Lendl, B.; Rajamanickam, V.; Herwig, C.; et al. Teaching an old pET new tricks: Tuning of inclusion body formation and properties by a mixed feed system in E. coli. Appl. Microbiol. Biotechnol. 2018, 102, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Marisch, K.; Bayer, K.; Cserjan-Puschmann, M.; Luchner, M.; Striedner, G. Evaluation of three industrial Escherichia coli strains in fed-batch cultivations during high-level SOD protein production. Microb. Cell Fact. 2013, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Rajamanickam, V.; Wurm, D.; Slouka, C.; Herwig, C.; Spadiut, O. A novel toolbox for E. coli lysis monitoring. Anal. Bioanal. Chem. 2017, 409, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Wurm, D.J.; Marschall, L.; Sagmeister, P.; Herwig, C.; Spadiut, O. Simple monitoring of cell leakiness and viability in Escherichia coli bioprocesses—A case study. Eng. Life Sci. 2017, 17, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, J.; Slouka, C.; Ulonska, S.; Kager, J.; Fricke, J.; Spadiut, O.; Herwig, C. Impact of Glycerol as Carbon Source onto Specific Sugar and Inducer Uptake Rates and Inclusion Body Productivity in E. coli BL21(DE3). Bioengineering 2017, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klumpp, S.; Zhang, Z.; Hwa, T. Growth rate-dependent global effects on gene expression in bacteria. Cell 2009, 139, 1366–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terpe, K. Overview of bacterial expression systems for heterologous protein production: From molecular and biochemical fundamentals to commercial systems. Appl. Microbiol. Biotechnol. 2006, 72, 211. [Google Scholar] [CrossRef] [PubMed]
- Takkinen, K.; Laukkanen, M.L.; Sizmann, D.; Alfthan, K.; Immonen, T.; Vanne, L.; Kaartinen, M.; Knowles, J.K.; Teeri, T.T. An active single-chain antibody containing a cellulase linker domain is secreted by Escherichia coli. Protein Eng. 1991, 4, 837–841. [Google Scholar] [CrossRef]
- Striedner, G.; Cserjan-Puschmann, M.; Pötschacher, F.; Bayer, K. Tuning the Transcription Rate of Recombinant Protein in Strong Escherichiacoli Expression Systems through Repressor Titration. Biotechnol. Prog. 2003, 19, 1427–1432. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Yoda, K.; Yamasaki, M.; Tamura, G. The nucleotide sequence of the promoter and the amino-terminal region of alkaline phosphatase structural gene (phoA) of Escherichia coli. Nucleic Acids Res. 1981, 9, 5671–5678. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, Y.; Luo, M.; Cagliero, C.; Jiang, H.; Xie, Y.; Zhu, J.; Lu, H. A PhoA-STII Based Method for Efficient Extracellular Secretion and Purification of Fab from Escherichia coli. Bio-Protocol 2019, 9, e3370. [Google Scholar] [CrossRef]
- Schuhmacher, T.; Löffler, M.; Hurler, T.; Takors, R. Phosphate limited fed-batch processes: Impact on carbon usage and energy metabolism in Escherichia coli. J. Biotechnol. 2014, 190, 96–104. [Google Scholar] [CrossRef]
- Rao, N.N.; Liu, S.; Kornberg, A. Inorganic polyphosphate in Escherichia coli: The phosphate regulon and the stringent response. J. Bacteriol. 1998, 180, 2186–2193. [Google Scholar] [CrossRef] [Green Version]
- Wanner, B.L. Phosphorus assimilation and control of the phosphate regulon. In Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed.; ASM Press: Washington, DC, USA, 1996; Volume 41, pp. 1357–1381. [Google Scholar]
- Wanner, B. Phosphate regulation of gene expression in E. coli. In Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology; ASM Press: Washington, DC, USA, 1987; Volume 2, pp. 1326–1333. [Google Scholar]
- Marzan, L.W.; Shimizu, K. Metabolic regulation of Escherichia coli and its phoB and phoR genes knockout mutants under phosphate and nitrogen limitations as well as at acidic condition. Microb. Cell Fact. 2011, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baca, M.; Wells, J.A. Anti-VEGF Antibodies. US Patent 6,884,879, 26 April 2005. [Google Scholar]
- Romano, S.; Schulz-Vogt, H.N.; González, J.M.; Bondarev, V. Phosphate limitation induces drastic physiological changes, virulence-related gene expression, and secondary metabolite production in Pseudovibrio sp. strain FO-BEG1. Appl. Environ. Microbiol. 2015, 81, 3518–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doran, P.M. Bioprocess Engineering Principles, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Santos-Beneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef] [Green Version]
- Ghorbel, S.; Smirnov, A.; Chouayekh, H.; Sperandio, B.; Esnault, C.; Kormanec, J.; Virolle, M.J. Regulation of ppk expression and in vivo function of Ppk in Streptomyces lividans TK24. J. Bacteriol. 2006, 188, 6269–6276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanner, B.L. Signal transduction in the control of phosphate-regulated genes of Escherichia coli. Kidney Int. 1996, 49, 964–967. [Google Scholar] [CrossRef] [Green Version]
- Muller, J.M.; Wetzel, D.; Flaschel, E.; Friehs, K.; Risse, J.M. Constitutive production and efficient secretion of soluble full-length streptavidin by an Escherichia coli “leaky mutant”. J. Biotechnol. 2016, 221, 91–100. [Google Scholar] [CrossRef]
- Morowvat, M.H.; Babaeipour, V.; Rajabi-Memari, H.; Vahidi, H.; Maghsoudi, N. Overexpression of Recombinant Human Beta Interferon (rhINF-beta) in Periplasmic Space of Escherichia coli. Iran. J. Pharm. Res. 2014, 13, 151–160. [Google Scholar]
- Song, H.; Jiang, J.; Wang, X.; Zhang, J. High purity recombinant human growth hormone (rhGH) expression in Escherichia coli under phoA promoter. Bioengineered 2017, 8, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Agbogbo, F.K.; Ramsey, P.; George, R.; Joy, J.; Srivastava, S.; Huang, M.; McCool, J. Upstream development of Escherichia coli fermentation process with PhoA promoter using design of experiments (DoE). J. Ind. Microbiol. Biotechnol. 2020, 47, 789–799. [Google Scholar] [CrossRef]
- Sohoni, S.V.; Nelapati, D.; Sathe, S.; Javadekar-Subhedar, V.; Gaikaiwari, R.P.; Wangikar, P.P. Optimization of high cell density fermentation process for recombinant nitrilase production in E. coli. Bioresour. Technol. 2015, 188, 202–208. [Google Scholar] [CrossRef]
- Maldonado, L.M.; Hernandez, V.E.; Rivero, E.M.; Barba de la Rosa, A.P.; Flores, J.L.; Acevedo, L.G.; De Leon Rodriguez, A. Optimization of culture conditions for a synthetic gene expression in Escherichia coli using response surface methodology: The case of human interferon beta. Biomol. Eng. 2007, 24, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Saida, F. Overview on the expression of toxic gene products in Escherichia coli. Curr. Protoc. Protein Sci. 2007. [Google Scholar] [CrossRef]
- Grunberg, S.; Wolf, E.J.; Jin, J.; Ganatra, M.B.; Becker, K.; Ruse, C.; Taron, C.H.; Correa, I.R., Jr.; Yigit, E. Enhanced expression and purification of nucleotide-specific ribonucleases MC1 and Cusativin. Protein Expr. Purif. 2022, 190, 105987. [Google Scholar] [CrossRef] [PubMed]
- Fathi-Roudsari, M.; Maghsoudi, N.; Maghsoudi, A.; Niazi, S.; Soleiman, M. Auto-induction for high level production of biologically active reteplase in Escherichia coli. Protein Expr. Purif. 2018, 151, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, V.M.; Campani, G.; Santos, M.P.; Silva, G.G.; Pires, M.C.; Goncalves, V.M.; de Giordano, C.R.; Sargo, C.R.; Horta, A.C.L.; Zangirolami, T.C. Cost analysis based on bioreactor cultivation conditions: Production of a soluble recombinant protein using Escherichia coli BL21(DE3). Biotechnol. Rep. 2020, 26, e00441. [Google Scholar] [CrossRef]
- Tahara, N.; Tachibana, I.; Takeo, K.; Yamashita, S.; Shimada, A.; Hashimoto, M.; Ohno, S.; Yokogawa, T.; Nakagawa, T.; Suzuki, F.; et al. Boosting Auto-Induction of Recombinant Proteins in Escherichia coli with Glucose and Lactose Additives. Protein Pept. Lett. 2020, 28, 1180–1190. [Google Scholar] [CrossRef]
- Studier, F.W. Use of T7 RNA polymerase to direct expression of cloned genes. Methods Enzymol. 1990, 185, 60–89. [Google Scholar]
- Rao, N.N.; Wang, E.; Yashphe, J.; Torriani, A. Nucleotide pool in pho regulon mutants and alkaline phosphatase synthesis in Escherichia coli. J. Bacteriol. 1986, 166, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ding, H.; Du, P.; Gan, R.; Ye, Q. Production of phoA promoter-controlled human epidermal growth factor in fed-batch cultures of Escherichia coli YK537 (pAET-8). Process Biochem. 2005, 40, 3068–3074. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cultivation | µ | Temp. | Induction | DCW | Glucose | Acetate | YX/S | YCO2/S | C-Balance |
|---|---|---|---|---|---|---|---|---|---|
| (h−1) | (°C) | (h) | (g/L) | (g/L) | (g/L) | (C-mol/C-mol) | (C-mol/C-mol) | ||
| pET 1 | 0.1 | 35 | 4 h | 40.1 | 1.52 | 0 | 0.33 | 0.47 | 0.80 |
| 8 h | 40.3 + | 93.1 | 0.42 | n.d. * | n.d. * | n.d. * | |||
| pET 2 | 0.1 | 30 | 4 h | 41.3 | 0.93 | 0.87 | 0.36 | 0.47 | 0.85 |
| 8 h | 42.6 | 21.7 | 0 | 0.25 | 0.61 | 0.84 | |||
| pET 3 | 0.05 | 35 | 4 h | 33.9 | 0 | 0.33 | 0.33 | 0.52 | 0.85 |
| 8 h | 36.9 | 0 | 0 | 0.27 | 0.56 | 0.83 | |||
| pET 4 | 0.05 | 30 | 4 h | 36.4 | 0 | 0.52 | 0.37 | 0.48 | 0.85 |
| 8 h | 40.4 | 0 | 0.59 | 0.27 | 0.51 | 0.78 |
| Fab Insoluble (IBs) | Fab Soluble | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cult. | µ | Temp. | Ind. | Spec. Titer | Vol. Titer | STY | Spec. Titer | Vol. Titer | STY | Ratio |
| (h−1) | (°C) | (h) | (mg/g) | (mg/L) | (mg/L/h) | (mg/g) | (mg/L) | (mg/L/h) | IB:SP * | |
| pET 1 | 0.1 | 35 | 4 h | 19.5 | 782.6 | 33.4 | 2.22 | 89.0 | 3.79 | 8.8 |
| 8 h | 22.2 | 896.6 | 32.2 | 1.89 | 76.5 | 2.75 | 11.8 | |||
| pET 2 | 0.1 | 30 | 4 h | 14.9 | 613.0 | 26.0 | 2.89 | 119.5 | 5.07 | 5.1 |
| 8 h | 22.1 | 942.4 | 33.9 | 2.42 | 102.9 | 3.71 | 9.1 | |||
| pET 3 | 0.05 | 35 | 4 h | 20.4 | 693.6 | 29.3 | 2.38 | 80.6 | 3.41 | 8.6 |
| 8 h | 24.6 | 907.7 | 33.5 | 2.20 | 82.0 | 2.99 | 11.2 | |||
| pET 4 | 0.05 | 30 | 4 h | 12.0 | 436.5 | 18.3 | 2.81 | 102.9 | 4.32 | 4.2 |
| 8 h | 20.9 | 841.0 | 30.4 | 2.50 | 101.5 | 3.67 | 8.3 | |||
| Cultivation | µ | Temp. | Sample | PO4 Conc. | Cultivation Time |
|---|---|---|---|---|---|
| (h−1) | (°C) | (-) | (mM) | (h) | |
| pAT 1 | 0.1 | 35 | >1 mM PO4 | 2.37 | 22.3 |
| PO4 starvation | 0.16 | 27.3 | |||
| pAT 2 | 0.1 | 30 | >1 mM PO4 | 2.82 | 22.4 |
| PO4 starvation | <0.10 | 25.2 | |||
| pAT 3 | 0.05 | 35 | >1 mM PO4 | 4.57 | 43.2 |
| PO4 starvation | 0.39 * | 54.8 | |||
| pAT 4 | 0.05 | 30 | >1 mM PO4 | 4.46 | 37.1 |
| PO4 starvation | 0.10 | 47.3 |
| Cult. | µ | Temp. | Sample | DCW | qPO4 | Glucose | Acetate | YX/S | YCO2/S | C-Balance |
|---|---|---|---|---|---|---|---|---|---|---|
| (h−1) | (°C) | (-) | (g/L) | (mmol/g/h) | (g/L) | (g/L) | (C-mol/C-mol) | (C-mol/C-mol) | (-) | |
| pAT 1 | 0.1 | 35 | >1 mM PO4 | 42.8 | 0.112 | 0 | 0.28 | 0.45 | 0.42 | 0.87 |
| PO4 starvation | 53.2 | 0.008 | 1.70 | 0.99 | 0.30 | 0.49 | 0.80 | |||
| pAT 2 | 0.1 | 30 | >1 mM PO4 | 47.6 | 0.097 | 0 | 0.46 | 0.44 | 0.52 | 0.96 |
| PO4 starvation | 52.0 | 0.018 | 0 | 0.69 | 0.27 | 0.85 | 1.13 | |||
| pAT 3 | 0.05 | 35 | >1 mM PO4 | 31.8 | 0.052 | 0 | - | 0.20 | 0.65 | 0.85 |
| PO4 starvation | 47.1 | 0.008 | 0 | 0.34 | 0.32 | 0.57 | 0.89 | |||
| pAT 4 | 0.05 | 30 | >1 mM PO4 | 43.0 | 0.055 | 0 | 0.28 | 0.47 | 0.49 | 0.97 |
| PO4 starvation | 52.6 | 0.008 | 0 | 0.58 | 0.29 | 0.70 | 0.99 |
| Fab Insoluble (IBs) | Fab Soluble | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cult. | µ | Temp. | Sample | Spec. Titer | Vol. Titer | STY | Spec. Titer | Vol. Titer | STY | Ratio |
| h−1) | (°C) | (-) | (mg/g) | (mg/L) | (mg/L/h) | (mg/g) | (mg/L) | (mg/L/h) | IB:SP * | |
| pAT 1 | 0.1 | 35 | >1 mM PO4 | 0 | 0 | 0 | 2.28 | 97.30 | 4.36 | n.a. |
| PO4 starvation | 0 | 0 | 0 | 3.21 | 170.9 | 6.24 | n.a. | |||
| pAT 2 | 0.1 | 30 | >1 mM PO4 | 0 | 0 | 0 | 2.95 | 140.4 | 6.27 | n.a. |
| PO4 starvation | 0 | 0 | 0 | 2.91 | 150.1 | 5.98 | n.a. | |||
| pAT 3 | 0.05 | 35 | >1 mM PO4 | 0 | 0 | 0 | 2.53 | 80.62 | 1.86 | n.a. |
| PO4 starvation | 8.16 | 385.0 | 7.02 | 2.54 | 119.5 | 2.18 | 3.2 | |||
| pAT 4 | 0.05 | 30 | >1 mM PO4 | 0 | 0 | 0 | 4.63 | 198.8 | 5.37 | n.a. |
| PO4 starvation | 7.88 | 414.2 | 8.76 | 6.09 | 321.1 | 6.77 | 1.3 | |||
| ICP-OES | IC | Colorimetric Kit | Cedex Bio HT | |
|---|---|---|---|---|
| Analyte | P | PO4 | PO4 | PO4 |
| Limit of Quantification | 65 µmol/L | 4 µmol/L | 5 µmol/L | 100 µmol/L |
| Equipment costs | − | − | + | − |
| Sample preparation | − | − | − | + |
| Operator’s impact | ~ | ~ | − | + |
| Time | >>30 min | >>30 min | >>30 min | 15 min |
| Automation | − | − | − | + |
| At-line measurement | − | − | − | + |
| Process Time | DCW | µ | cPO4 | qPO4 | YCO2/S | YX/S | C-Balance |
|---|---|---|---|---|---|---|---|
| (h) | (g/L) | (h−1) | (mM) | (mmol/g/h) | (C-mol/C-mol) | (C-mol/C-mol) | (-) |
| 27.4 | 29.0 | n.a. | 33.5 | n.a. | 0.54 | 0.55 | 1.09 |
| 29.3 | 30.7 | 0.040 | 27.5 | 0.090 | 0.58 | 0.38 | 0.98 |
| 31.3 | 32.0 | 0.036 | 24.3 | 0.059 | 0.59 | 0.32 | 0.89 |
| 33.3 | 34.3 | 0.049 | 20.2 | 0.053 | 0.58 | 0.42 | 0.99 |
| 35.3 | 37.0 | 0.054 | 14.6 | 0.070 | 0.57 | 0.45 | 1.01 |
| 37.3 | 39.0 | 0.045 | 8.7 | 0.071 | 0.56 | 0.36 | 0.91 |
| 39.3 | 41.4 | 0.049 | 3.6 | 0.061 | 0.56 | 0.38 | 0.93 |
| 41.3 | 45.5 | 0.070 | 1.9 | 0.018 | 0.54 | 0.54 | 1.07 |
| 43.3 | 48.7 | 0.057 | 0.13 | 0.018 | 0.56 | 0.43 | 0.99 |
| 45.0 * | 48.9 | 0.021 | <0.10 | 0.0003 | 0.82 | 0.19 | 0.99 |
| Process Time | cPO4 | qPO4 | Spec. Fab Titer | Vol. Fab Titer | qFab | Fab STY |
|---|---|---|---|---|---|---|
| (h) | (mM) | (mmol/g/h) | (mg/g) | (mg/L) | (mg/g/h) | (mg/L/h) |
| 27.4 | 33.5 | n.a. | 5.26 | 153 | n.a. | n.a. |
| 29.3 | 27.5 | 0.090 | 5.46 | 167 | 0.31 | 9.4 |
| 31.3 | 24.3 | 0.059 | 5.65 | 181 | 0.29 | 9.2 |
| 33.3 | 20.2 | 0.053 | 5.56 | 191 | 0.23 | 7.6 |
| 35.3 | 14.6 | 0.070 | 5.49 | 203 | 0.26 | 9.3 |
| 37.3 | 8.7 | 0.071 | 5.72 | 223 | 0.37 | 14.0 |
| 39.3 | 3.6 | 0.061 | 5.77 | 239 | 0.31 | 12.4 |
| 41.3 | 1.9 | 0.018 | 5.73 | 261 | 0.38 | 16.6 |
| 43.3 | 0.13 | 0.018 | 6.43 | 313 | 0.69 | 32.5 |
| 45.0 * | <0.10 | 0.0003 | 7.15 | 350 | 0.59 | 28.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gundinger, T.; Kittler, S.; Kubicek, S.; Kopp, J.; Spadiut, O. Recombinant Protein Production in E. coli Using the phoA Expression System. Fermentation 2022, 8, 181. https://doi.org/10.3390/fermentation8040181
Gundinger T, Kittler S, Kubicek S, Kopp J, Spadiut O. Recombinant Protein Production in E. coli Using the phoA Expression System. Fermentation. 2022; 8(4):181. https://doi.org/10.3390/fermentation8040181
Chicago/Turabian StyleGundinger, Thomas, Stefan Kittler, Sabine Kubicek, Julian Kopp, and Oliver Spadiut. 2022. "Recombinant Protein Production in E. coli Using the phoA Expression System" Fermentation 8, no. 4: 181. https://doi.org/10.3390/fermentation8040181