
Enhancement of PHA Production by a Mixed Microbial Culture Using VFA Obtained from the Fermentation of Wastewater from Yeast Industry

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
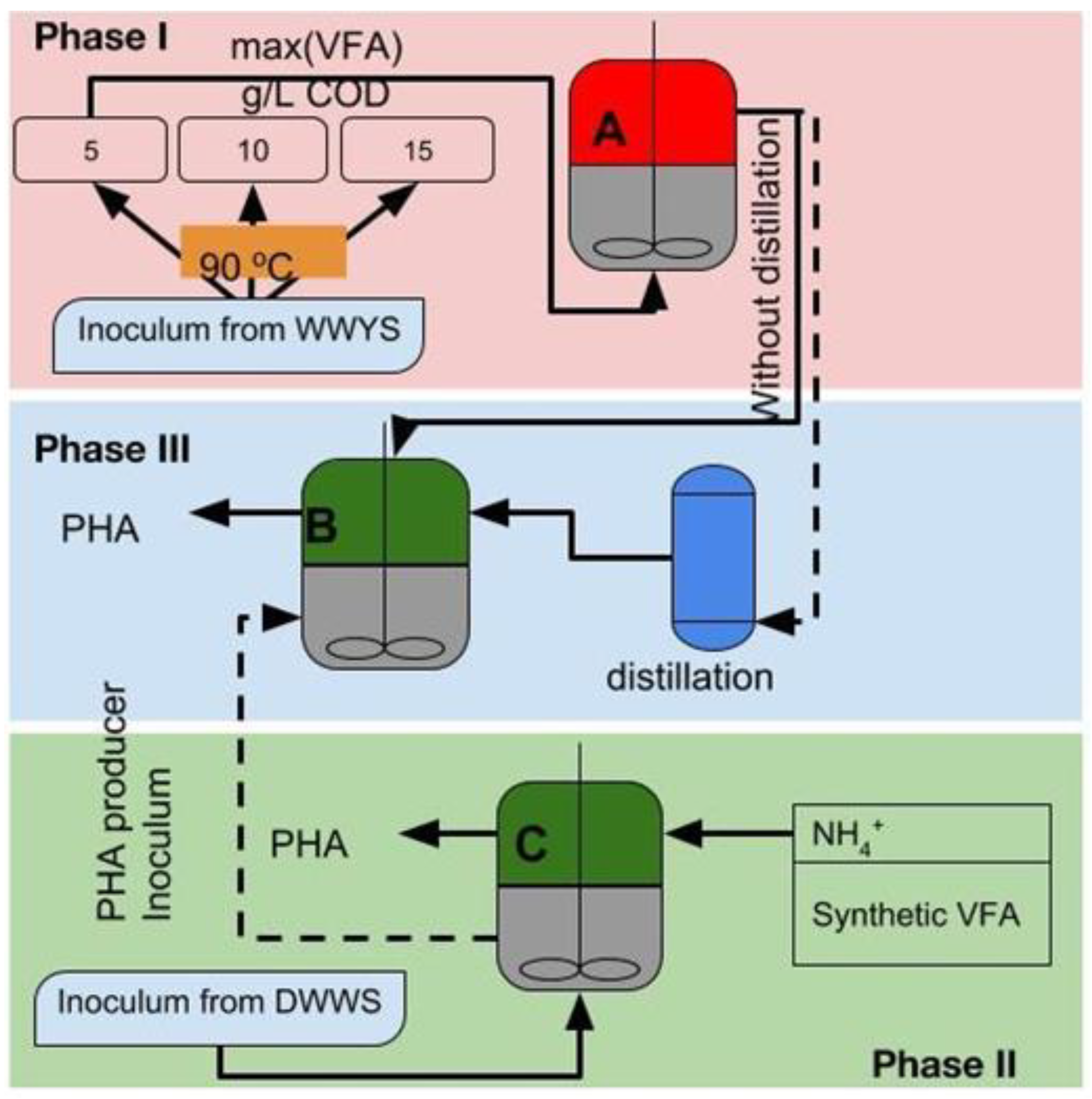
2.1. Phase I: Inoculum Adaptation for VFA Production
2.2. Phase II: Adaptation of Inoculum for PHA Production
2.3. Phase III: PHA Production in a Coupled System
2.4. Physicochemical Analysis
2.5. Microbiological Quantification of Anaerobic Bacteria
2.6. Molecular Techniques for Diversity Analysis
3. Results and Discussion
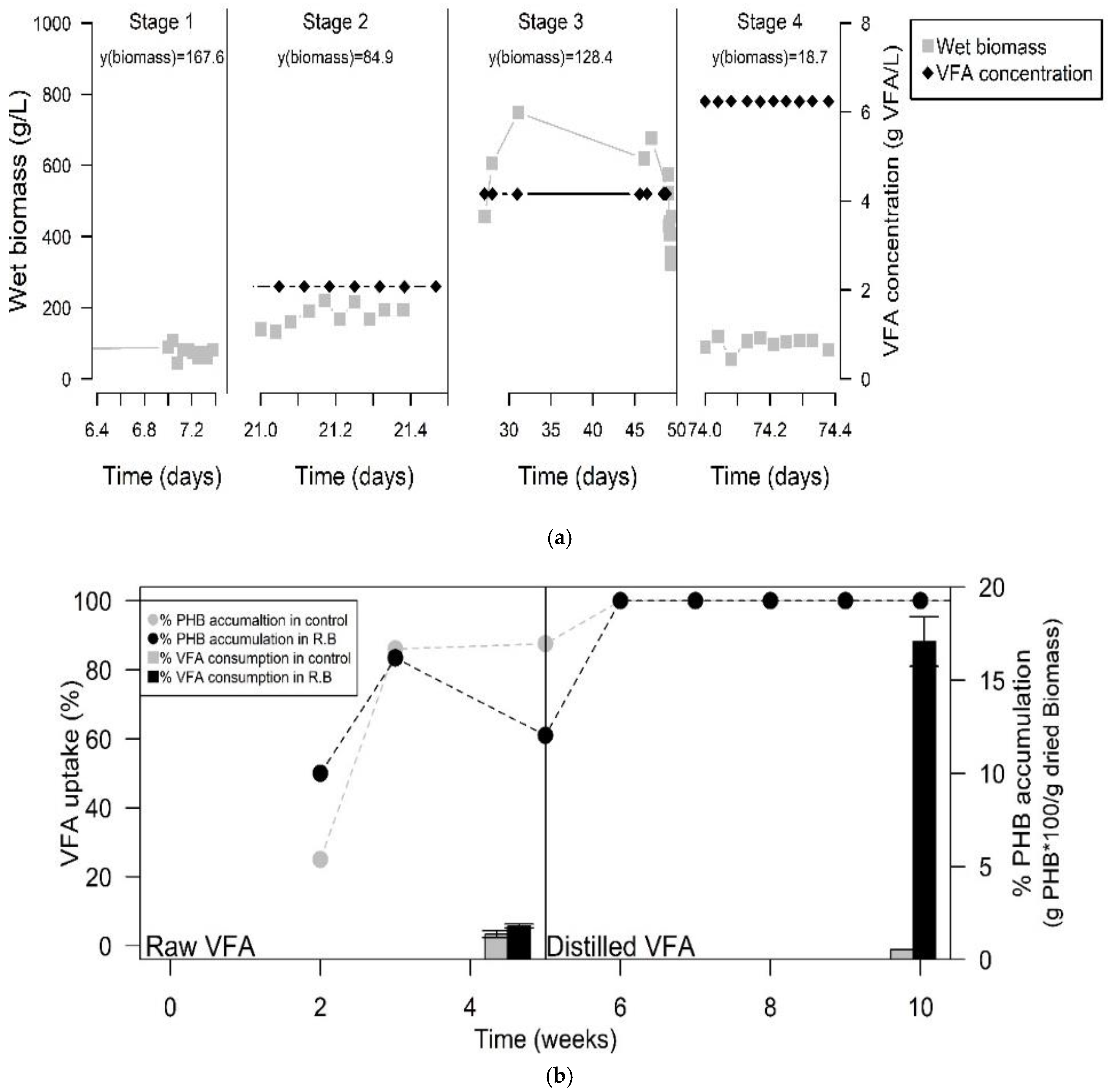
3.1. Dark Fermentation of WWY
3.2. Assessment of Anaerobic Bacteria in the Acidogenic Reactor
3.3. Adaptation of the Inoculum for PHA Production
3.4. PHA Production in a Coupled System
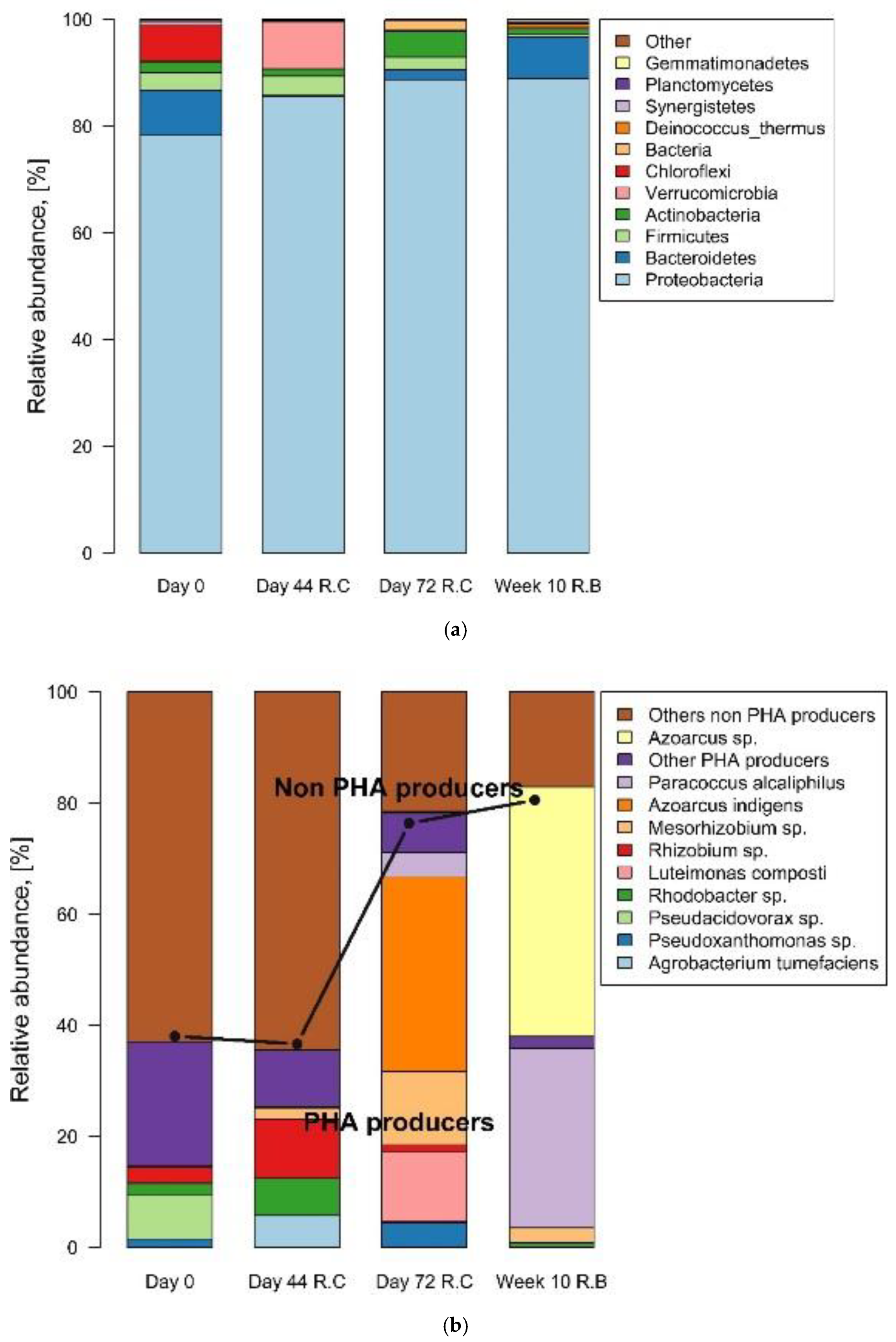
3.5. Characterization of the PHB-Producing Microbial Community
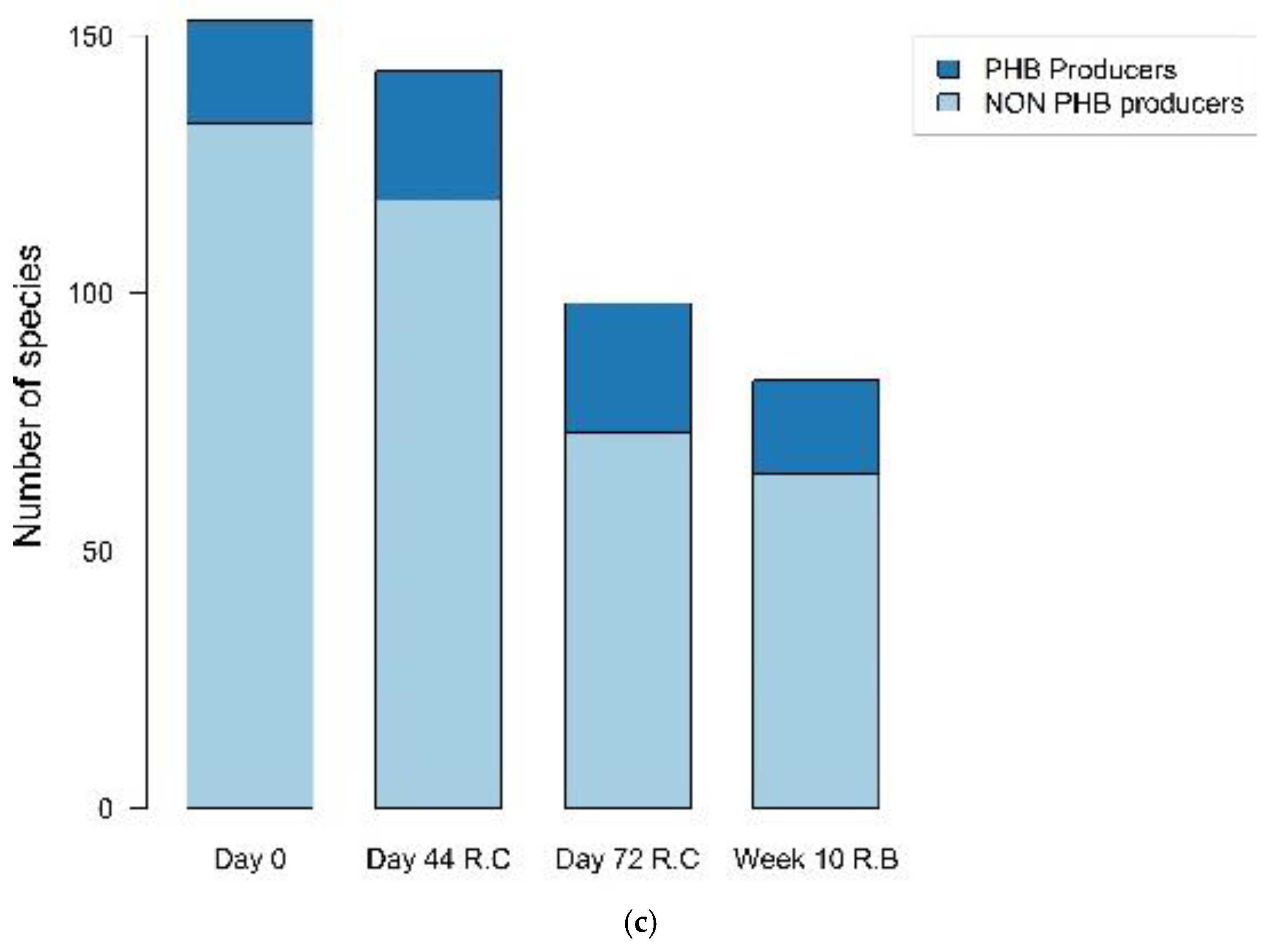
3.6. Analysis of Abundance of PHA-Producing Species
3.7. Biodiversity Indexes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heitmann, M.; Zannini, E.; Arendt, E. Impact of Saccharomyces Cerevisiae Metabolites Produced during Fermentation on Bread Quality Parameters: A Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1152–1164. [Google Scholar] [CrossRef] [PubMed]
- Pirsaheb, M.; Mohamadi, S.; Rahmatabadi, S.; Hossini, H.; Motteran, F. Simultaneous Wastewater Treatment and Biogas Production Using Integrated Anaerobic Baffled Reactor Granular Activated Carbon from Baker’s Yeast Wastewater. Environ. Technol. 2018, 39, 2724–2735. [Google Scholar] [CrossRef] [PubMed]
- Ersahin, M.E.; Ozgun, H.; Dereli, R.K.; Ozturk, I. Anaerobic Treatment of Industrial Effluents: An Overview of Applications. In Waste Water—Treatment and Reutilization; InTech: Rijeka, Croatia, 2011; ISBN 9789789533072494. [Google Scholar]
- Igwegbe, C.A.; Obiora-Okafo, I.A.; Iwuozor, K.O.; Ghosh, S.; Kurniawan, S.B.; Rangabhashiyam, S.; Kanaoujiya, R.; Ighalo, J.O. Treatment Technologies for Bakers’ Yeast Production Wastewater. Environ. Sci. Pollut. Res. 2022, 29, 11004–11026. [Google Scholar] [CrossRef] [PubMed]
- Pirsaheb, M.; Rostamifar, M.; Mansouri, A.M.; Zinatizadeh, A.A.L.; Sharafi, K. Performance of an Anaerobic Baffled Reactor (ABR) Treating High Strength Baker’s Yeast Manufacturing Wastewater. J. Taiwan Inst. Chem. Eng. 2015, 47, 137–148. [Google Scholar] [CrossRef]
- Mohan, S.; Vivekanandhan, V.; Priyadharshini, S. Performance Evaluation of Modified UASB Reactor for Treating Bakery Effluent. Int. J. Appl. Environ. Sci. 2017, 12, 1883–1894. [Google Scholar]
- Balcıoğlu, G.; Gönder, Z.B. Recovery of Baker’s Yeast Wastewater with Membrane Processes for Agricultural Irrigation Purpose: Fouling Characterization. Chem. Eng. J. 2014, 255, 630–640. [Google Scholar] [CrossRef]
- Uysal, A.; Demir, S.; Sayilgan, E.; Eraslan, F.; Kucukyumuk, Z. Optimization of Struvite Fertilizer Formation from Baker’s Yeast Wastewater: Growth and Nutrition of Maize and Tomato Plants. Environ. Sci. Pollut. Res. 2014, 21, 3264–3274. [Google Scholar] [CrossRef]
- Begea, M.; Berkesy, C.; Berkesy, L.; Cîrîc, A.; Bărbulescu, I.D.; Gavriloaie, C. Study of the Recovery in Agriculture of the Waste Resulted from Baker’s Yeast Industry. Adv. Agric. Bot. 2017, 9, 136–145. [Google Scholar]
- Sajad Hashemi, S.; Karimi, K.; Taherzadeh, M.J. Integrated Process for Protein, Pigments, and Biogas Production from Baker’s Yeast Wastewater Using Filamentous Fungi. Bioresour. Technol. 2021, 337, 125356. [Google Scholar] [CrossRef]
- Pakalapati, H.; Chang, C.-K.; Show, P.L.; Arumugasamy, S.K.; Lan, J.C.-W. Development of Polyhydroxyalkanoates Production from Waste Feedstocks and Applications. J. Biosci. Bioeng. 2018, 126, 282–292. [Google Scholar] [CrossRef]
- Bhalerao, A.; Banerjee, R.; Nogueira, R. Continuous Cultivation Strategy for Yeast Industrial Wastewater-Based Polyhydroxyalkanoate Production. J. Biosci. Bioeng. 2020, 129, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, M.G.E.; Concas, S.; Bengtsson, S.; Reis, M.A.M. Mixed Culture Polyhydroxyalkanoates Production from Sugar Molasses: The Use of a 2-Stage CSTR System for Culture Selection. Bioresour. Technol. 2010, 101, 7123–7133. [Google Scholar] [CrossRef] [PubMed]
- Sathya, A.B.; Sivasubramanian, V.; Santhiagu, A.; Sebastian, C.; Sivashankar, R. Production of Polyhydroxyalkanoates from Renewable Sources Using Bacteria. J. Polym. Environ. 2018, 26, 3995–4012. [Google Scholar] [CrossRef]
- Guerra-Blanco, P.; Cortes, O.; Poznyak, T.; Chairez, I.; García-Peña, E.I. Polyhydroxyalkanoates (PHA) Production by Photoheterotrophic Microbial Consortia: Effect of Culture Conditions over Microbial Population and Biopolymer Yield and Composition. Eur. Polym. J. 2018, 98, 94–104. [Google Scholar] [CrossRef]
- Majone, M.; Dircks, K.; Beim, J.J. Aerobic Storage under Dynamic Conditions in Activated Sludge Processes. The State of the Art. Water Sci. Technol. 1999, 39, 61–73. [Google Scholar] [CrossRef]
- Valentino, F.; Morgan-Sagastume, F.; Campanari, S.; Villano, M.; Werker, A.; Majone, M. Carbon Recovery from Wastewater through Bioconversion into Biodegradable Polymers. New Biotechnol. 2017, 37, 9–23. [Google Scholar] [CrossRef] [Green Version]
- Lazaro, C.Z.; Perna, V.; Etchebehere, C.; Varesche, M.B.A. Sugarcane Vinasse as Substrate for Fermentative Hydrogen Production: The Effects of Temperature and Substrate Concentration. Int. J. Hydrogen Energy 2014, 39, 6407–6418. [Google Scholar] [CrossRef]
- Shen, L.; Hu, H.; Ji, H.; Cai, J.; He, N.; Li, Q.; Wang, Y. Production of Poly(Hydroxybutyrate-Hydroxyvalerate) from Waste Organics by the Two-Stage Process: Focus on the Intermediate Volatile Fatty Acids. Bioresour. Technol. 2014, 166, 194–200. [Google Scholar] [CrossRef]
- Janarthanan, O.M.; Laycock, B.; Montano-Herrera, L.; Lu, Y.; Arcos-Hernandez, M.V.; Werker, A.; Pratt, S. Fluxes in PHA-Storing Microbial Communities during Enrichment and Biopolymer Accumulation Processes. New Biotechnol. 2016, 33, 61–72. [Google Scholar] [CrossRef]
- Morgan-Sagastume, F. Characterisation of Open, Mixed Microbial Cultures for Polyhydroxyalkanoate (PHA) Production. Rev. Environ. Sci. Bio/Technol. 2016, 15, 593–625. [Google Scholar] [CrossRef]
- Valentino, F.; Karabegovic, L.; Majone, M.; Morgan-Sagastume, F.; Werker, A. Polyhydroxyalkanoate (PHA) Storage within a Mixed-Culture Biomass with Simultaneous Growth as a Function of Accumulation Substrate Nitrogen and Phosphorus Levels. Water Res. 2015, 77, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Korkakaki, E.; van Loosdrecht, M.C.M.; Kleerebezem, R. Survival of the Fastest: Selective Removal of the Side Population for Enhanced PHA Production in a Mixed Substrate Enrichment. Bioresour. Technol. 2016, 216, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bengtsson, S.; Oehmen, A.; Carvalho, G.; Werker, A.; Reis, M.A.M. Application of Dissolved Oxygen (DO) Level Control for Polyhydroxyalkanoate (PHA) Accumulation with Concurrent Nitrification in Surplus Municipal Activated Sludge. New Biotechnol. 2019, 50, 37–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamis, J.; Lužkov, K.; Jiang, Y.; van Loosdrecht, M.C.M.; Kleerebezem, R. Enrichment of Plasticicumulans Acidivorans at Pilot-Scale for PHA Production on Industrial Wastewater. J. Biotechnol. 2014, 192, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiao, K.; Jiang, X.; Shen, N.; Zeng, R.J.; Zhou, Y. In-Situ Sludge Pretreatment in a Single-Stage Anaerobic Digester. Bioresour. Technol. 2017, 238, 102–108. [Google Scholar] [CrossRef] [PubMed]
- APHA. Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association: Washington, DC, USA, 2017; ISBN 9780875532875. [Google Scholar]
- Martla, M.; Umsakul, K.; Sudesh, K. Production and Recovery of Poly(3-Hydroxybutyrate- Co -3-Hydroxyvalerate) from Biodiesel Liquid Waste (BLW). J. Basic Microbiol. 2018, 58, 977–986. [Google Scholar] [CrossRef]
- ASTM. Standard Test Methods for Chemical Oxygen Demand (Dichromate Oxygen Demand) of Water D1252-06; ASTM International: West Conshohocken, PA, USA, 2012. [Google Scholar]
- Moreno, P.; Yañez, C.; Cardozo, N.S.M.; Escalante, H.; Combariza, M.Y.; Guzman, C. Influence of Nutritional and Physicochemical Variables on PHB Production from Raw Glycerol Obtained from a Colombian Biodiesel Plant by a Wild-Type Bacillus Megaterium Strain. New Biotechnol. 2015, 32, 682–689. [Google Scholar] [CrossRef]
- Sutton, S. The Most Probable Number Method and Its Uses in Enumeration, Qualification, and Validation. J. Valid. Technol. 2010, 16, 35–38. [Google Scholar]
- Keskin, A.; Bursali, A.; Snow, D.E.; Dowd, S.E.; Tekin, S. Assessment of Bacterial Diversity in Hyalomma Aegyptium, H. Marginatum and H. Excavatum Ticks through Tag-Encoded Pyrosequencing. Exp. Appl. Acarol. 2017, 73, 461–475. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and Tools for High Throughput RRNA Analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [Green Version]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamran, M. Bioenergy. In Renewable Energy Conversion Systems; Elsevier: Amsterdam, The Netherlands, 2021; pp. 243–264. [Google Scholar]
- Zheng, H.; Zeng, R.J.; O’Sullivan, C.; Clarke, W.P. Critical Analysis of Hydrogen Production from Mixed Culture Fermentation under Thermophilic Condition (60 °C). Appl. Microbiol. Biotechnol. 2016, 100, 5165–5176. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Yan, B.; Wong, J.W.C.; Zhang, Y. Enhanced Volatile Fatty Acids Production from Anaerobic Fermentation of Food Waste: A Mini-Review Focusing on Acidogenic Metabolic Pathways. Bioresour. Technol. 2018, 248, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Buitrón, G.; Carvajal, C. Biohydrogen Production from Tequila Vinasses in an Anaerobic Sequencing Batch Reactor: Effect of Initial Substrate Concentration, Temperature and Hydraulic Retention Time. Bioresour. Technol. 2010, 101, 9071–9077. [Google Scholar] [CrossRef]
- Murali, N.; Srinivas, K.; Ahring, B.K. Biochemical Production and Separation of Carboxylic Acids for Biorefinery Applications. Fermentation 2017, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Tamang, P.; Arndt, C.; Bruns-Hellberg, J.; Nogueira, R. Polyhydroxyalkanoates Production from Industrial Wastewaters Using a Mixed Culture Enriched with Thauera Sp.: Inhibitory Effect of the Wastewater Matrix. Environ. Technol. Innov. 2021, 21, 101328. [Google Scholar] [CrossRef]
- Morgan-Sagastume, F.; Karlsson, A.; Johansson, P.; Pratt, S.; Boon, N.; Lant, P.; Werker, A. Production of Polyhydroxyalkanoates in Open, Mixed Cultures from a Waste Sludge Stream Containing High Levels of Soluble Organics, Nitrogen and Phosphorus. Water Res. 2010, 44, 5196–5211. [Google Scholar] [CrossRef]
- Queirós, D.; Rossetti, S.; Serafim, L.S. PHA Production by Mixed Cultures: A Way to Valorize Wastes from Pulp Industry. Bioresour. Technol. 2014, 157, 197–205. [Google Scholar] [CrossRef]
- Urakami, T.; Tamaoka, J.; Suzuki, K.-I.; Komagata, K. Paracoccus Alcoliphilus Sp. Nov., an Alkoliphilic and Facultatively Methylatrophic Bacterium. Int. J. Syst. Bacteriol. 1989, 39, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Fernández, H.; Prandoni, N.; Fernández-Pascual, M.; Fajardo, S.; Morcillo, C.; Díaz, E.; Carmona, M. Azoarcus Sp. CIB, an Anaerobic Biodegrader of Aromatic Compounds Shows an Endophytic Lifestyle. PLoS ONE 2014, 9, e110771. [Google Scholar] [CrossRef]
- Reinhold-Hurek, B.; Hurek, T.; Gillis, M.; Hoste, B.; Vancanneyt, M.; Kersters, K.; de Ley, J. Azoarcus Gen. Nov., Nitrogen-Fixing Proteobacteria Associated with Roots of Kallar Grass (Leptochloa Fusca (L.) Kunth), and Description of Two Species, Azoarcus Indigens Sp. Nov. and Azoarcus Communis Sp. Nov. Int. J. Syst. Bacteriol. 1993, 43, 574–584. [Google Scholar] [CrossRef]
- Wijeyekoon, S.; Carere, C.R.; West, M.; Nath, S.; Gapes, D. Mixed Culture Polyhydroxyalkanoate (PHA) Synthesis from Nutrient Rich Wet Oxidation Liquors. Water Res. 2018, 140, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Naeem, S.; Inchausti, P. Biodiversity and Ecosystem Functioning: Synthesis and Perspectives; Oxford University Press: Oxford, UK, 2002; ISBN 0198515715. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Preliminary Tests | Reactor A | Reactor C | Reactor B | Control Reactor |
|---|---|---|---|---|---|
| Type or reactor | Anaerobic Batch | Anaerobic SBR | Aerobic SBR | Aerobic SBR | Aerobic SBR |
| Type of wastewater | Yeast wastetwater | Yeast wastetwater | Mineral salts medium | Raw and distilled VFA | Raw VFA |
| Working Volume (L) | 0.7 | 1.7 | 1 | 0.2 | 0.2 |
| Volume of inoculum (L) | 0.03 | 0.1 | 0.5 | 0.1 | 0.1 |
| COD (g/L) | 5, 10 or 15 | 5 | - | - | - |
| VFA in feed (g/L) | - | - | 0.416–4.16 | 2–2.5 | 2–2.5 |
| VSS (mg/L) | 56–170 | 56.4 | 2.4 | 2.6 | 2.8 |
| pH | 7.65 | 7.65 | 8.9 | 7.8 | 8.7 |
| DO (mg/L) | - | - | 4 | 4.6 | 4.2 |
| HRT (days) | 18 | 7 | 2 | 2 | 2 |
| Total length of operation (days) | 18 | 70 | 78 | 70 | 70 |
| Air flow (L/min) | - | - | 6 | 6 | 6 |
| Stirring speed (rpm) | 110 | 110 | - | - | - |
| Stage | Time (Days) | VFA Concentration (g/L) | % PHB Accumulation (g PHB·100/g dry biomass−1) | PHB Concentration (g/L) | y(PHA) |
|---|---|---|---|---|---|
| 1 | 0 | 0.416 | 5.2 | 0.12 | 0.0011 |
| 2 | 21 | 2.08 | 10.3 | 0.80 | 0.0070 |
| 3 | 31 | 4.16 | 0.6 | 0.09 | 0.0005 |
| 3 | 49 | 4.16 | 1.0 | 0.09 | 0.0008 |
| 4 | 74 | 6.24 | 2.9 | 0.32 | 0.0180 |
| Time | Observed Richness | Simpson Diversity Index | Total Sequences | OTUs | |
|---|---|---|---|---|---|
| Reactor C | Day 0 | 40.55 | 0.93 | 5637 | 166 |
| Day 44 | 22.95 | 0.93 | 11,001 | 151 | |
| Day 74 | 14.43 | 0.84 | 11,755 | 107 | |
| Reactor B | Week 10 | 6.38 | 0.69 | 6282 | 87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ospina-Betancourth, C.; Echeverri, S.; Rodriguez-Gonzalez, C.; Wist, J.; Combariza, M.Y.; Sanabria, J. Enhancement of PHA Production by a Mixed Microbial Culture Using VFA Obtained from the Fermentation of Wastewater from Yeast Industry. Fermentation 2022, 8, 180. https://doi.org/10.3390/fermentation8040180
Ospina-Betancourth C, Echeverri S, Rodriguez-Gonzalez C, Wist J, Combariza MY, Sanabria J. Enhancement of PHA Production by a Mixed Microbial Culture Using VFA Obtained from the Fermentation of Wastewater from Yeast Industry. Fermentation. 2022; 8(4):180. https://doi.org/10.3390/fermentation8040180
Chicago/Turabian StyleOspina-Betancourth, Carolina, Sergio Echeverri, Claudia Rodriguez-Gonzalez, Julien Wist, Marianny Y. Combariza, and Janeth Sanabria. 2022. "Enhancement of PHA Production by a Mixed Microbial Culture Using VFA Obtained from the Fermentation of Wastewater from Yeast Industry" Fermentation 8, no. 4: 180. https://doi.org/10.3390/fermentation8040180