
Evaluation of Rye Bran Enzymatic Hydrolysate Effect on Gene Expression and Bacteriocinogenic Activity of Lactic Acid Bacteria
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Green Raw Materials and Enzyme Preparations
2.2. Preparation of Enzymatic Bran Hydrolysates
2.3. Bacterial Strains and Growth Conditions
2.4. Enumeration of Lactobacilli
2.5. Molecular Genetic Research Methods
2.5.1. LAB Biomass Separation for PCR Analyses
2.5.2. Identification of Bacteriocin-Related Structural Genes
2.5.3. Extraction of Total RNA
2.5.4. Quantitative Reverse Transcription Quantitative PCR and Gene Expression Analysis
2.6. Antimicrobial Activity Assay
3. Results
3.1. Screening of Lactic Acid Bacteria Strains for Bacteriocin Genes
3.2. Screening Lactic Acid Bacteria Strains for Bacteriocinogenic Activity
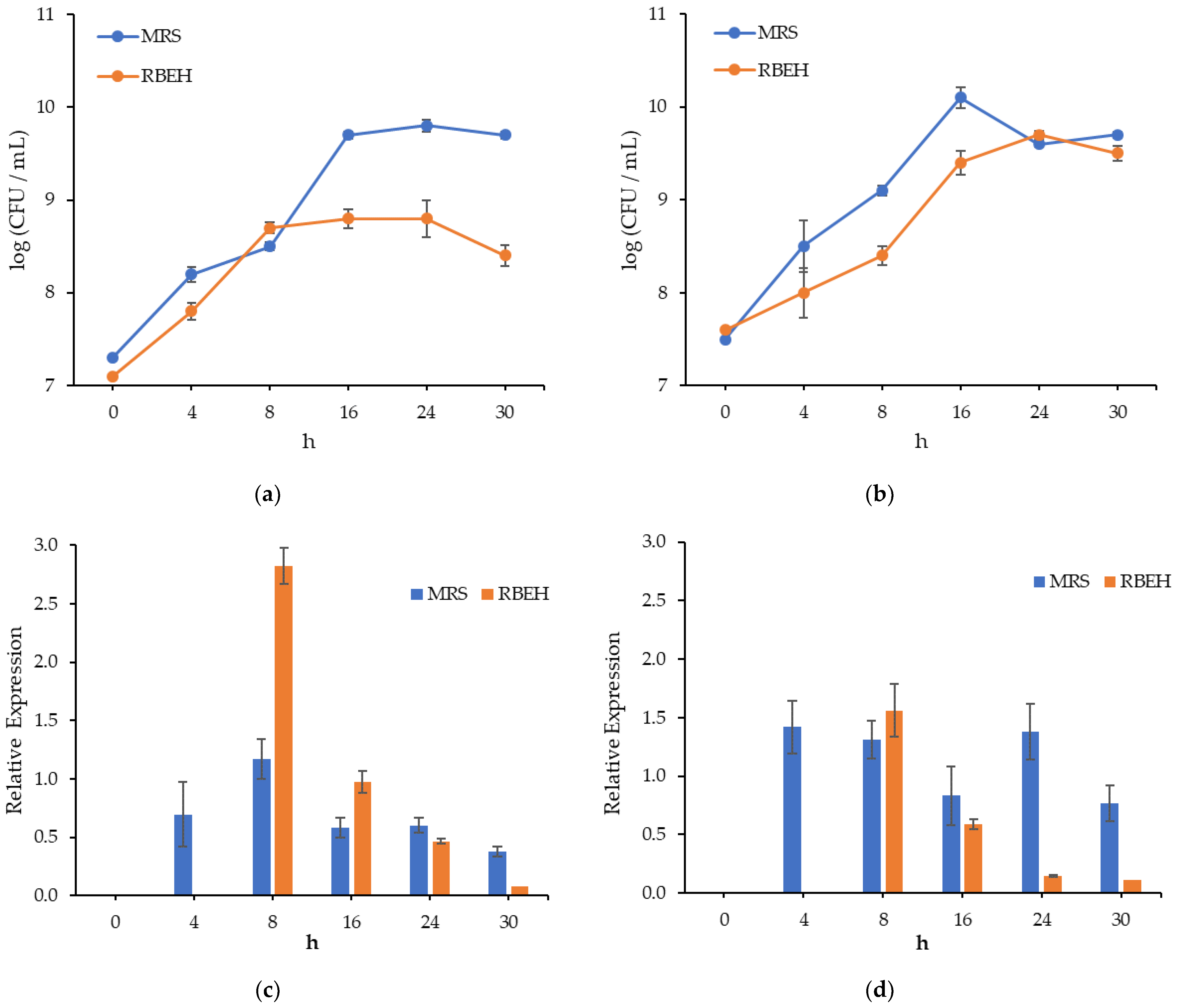
3.3. Dynamics of L. salivarius VKPM B-2214 and L. paracasei VKPM B-11657 Growth and Bacteriocin Genes Expression during Cultivation in MRS Broth and Rye Bran Hydrolysate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, Antimicrobial Peptides from Bacterial Origin: Overview of Their Biology and Their Impact against Multidrug-Resistant Bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Nishie, M.; Nagao, J.; Sonomoto, K. Antibacterial peptides bacteriocins: An overview of their diverse characteristics and applications. Biocontrol Sci. 2012, 17, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, M.; Turroni, F.; Lugli, G.A.; van Sinderen, D. Bifidobacteria of the Human Gut: Our Special Friends. In Diet-Microbe Interactions in the Gut, 1st ed.; Kieran, T., Daniele, D.R., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 41–51. [Google Scholar]
- Gradisteanu Pircalabioru, G.; Popa, L.I.; Marutescu, L.; Gheorghe, I.; Popa, M.; Czobor Barbu, I.; Cristescu, R.; Chifiriuc, M.C. Bacteriocins in the Era of Antibiotic Resistance: Rising to the Challenge. Pharmaceutics 2020, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, P.M.; Kuniyoshi, T.M.; Oliveira, R.P.; Hill, C.; Ross, R.P.; Cotter, P.D. Antimicrobials for food and feed; a bacteriocin perspective. Curr. Opin. Biotechnol. 2020, 61, 160–167. [Google Scholar] [CrossRef]
- Kuniyoshi, T.M.; Mendonça, C.M.; Viera, V.B.; Robl, D. Pediocin PA-1 production by Pediococcus pentosaceus ET34 using non-detoxified hemicellulose hydrolysate obtained from hydrothermal pretreatment of sugarcane bagasse. Bioresour. Technol. 2021, 338, 125565. [Google Scholar] [CrossRef]
- Sharma, S.; Garg, A.P.; Singh, G. Optimization of fermentation conditions for bacteriocin production by Lactococcus lactis CCSULAC1 on modified MRS medium. Int. J. Dairy Sci. 2010, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Furuta, Y.; Maruoka, N.; Nakamura, A.; Omori, T.; Sonomoto, K. Utilization of fermented barley extract obtained from a by-product of barley shochu for nisin production. J. Biosci. Bioeng. 2008, 106, 393–397. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Chen, S. Effects of nutrient supplements on simultaneous fermentation of nisin and lactic acid from cull potatoes. Appl. Biochem. Biotechnol. 2005, 121–124, 475–483. [Google Scholar] [CrossRef]
- Syrokou, M.K.; Stasinopoulou, P.; Paramithiotis, S.; Bosnea, L.; Mataragas, M.; Papadopoulos, G.K.; Skandamis, P.N.; Drosinos, E.H. The Effect of Incubation Temperature, Substrate and Initial pH Value on Plantaricin Activity and the Relative Transcription of pln Genes of Six Sourdough Derived Lactiplantibacillus plantarum Strains. Fermentation 2021, 7, 320. [Google Scholar] [CrossRef]
- Karetkin, B.A.; Panfilov, V.I.; Panfilova, E.V.; Khromova, N.Y.; Shakir, I.V. Heterogeneous submerged fermentation of probiotic in media based on wheat meal and by-products of wheat starch production. SGEM 2017, 17, 711–718. [Google Scholar]
- De Man, J.; Rogosa, M.; Sharpe, M. A medium for the cultivation of Lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T. Composition and metabolic activities of bacterial biofilms colonizing food residues in the human gut. Appl. Environ. Microbiol. 2006, 72, 6204–6211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macwana, S.J.; Muriana, P.M. A bacteriocin PCR array for identification of bacteriocin-related structural genes in lactic acid bacteria. J. Microbiol. Methods 2012, 88, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Wirawan, R.E.; Klesse, N.A.; Jack, R.W.; Tagg, J.R. Molecular and genetic characterization of a novel nisin variant produced by Streptococcus uberis. Appl. Environ. Microbiol. 2006, 72, 1148–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Turpin, W.; Humblot, C.; Noordine, M.L.; Thomas, M.; Guyot, J.P. Lactobacillaceae and cell adhesion: Genomic and functional screening. PLoS ONE 2012, 7, e38034. [Google Scholar] [CrossRef] [Green Version]
- Schillinger, U.; Kaya, M.; Lücke, F.K. Behaviour of Listeria monocytogenes in meat and its control by a bacteriocin-producing strain of Lactobacillus sakei. J. Appl. Bacteriol. 1991, 70, 473–478. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2006, 100, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Nissen-Meyer, J.; Oppegård, C.; Rogne, P.; Haugen, H.S.; Kristiansen, P.E. Structure and Mode-of-Action of the Two-Peptide (Class-IIb) Bacteriocins. Probiotics Antimicrob. Proteins 2010, 2, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Lozo, J.; Jovcic, B.; Kojic, M.; Dalgalarrondo, M.; Chobert, J.M.; Haertlé, T.; Topisirovic, L. Molecular characterization of a novel bacteriocin and an unusually large aggregation factor of Lactobacillus paracasei subsp. paracasei BGSJ2-8, a natural isolate from homemade cheese. Curr. Microbiol. 2007, 55, 266–271. [Google Scholar] [CrossRef]
- Tymoszewska, A.; Walczak, P.; Aleksandrzak-Piekarczyk, T. BacSJ-Another Bacteriocin with Distinct Spectrum of Activity that Targets Man-PTS. Int. J. Mol. Sci. 2020, 21, 7860. [Google Scholar] [CrossRef] [PubMed]
- Tolinački, M.; Lozo, J.; Veljović, K.; Kojić, M.; Fira, Đ.; Topisirović, L. Examination of antimicrobial potential in naturalisolates of Lactobacillus casei/paracasei group. Genetika 2012, 44, 661–677. [Google Scholar] [CrossRef]
- Castro, V.S.; Mutz, Y.d.S.; Rosario, D.K.A.; Cunha-Neto, A.; Figueiredo, E.E.d.S.; Conte-Junior, C.A. Inactivation of Multi-Drug Resistant Non-Typhoidal Salmonella and Wild-Type Escherichia coli STEC Using Organic Acids: A Potential Alternative to the Food Industry. Pathogens 2020, 9, 849. [Google Scholar] [CrossRef] [PubMed]
- Ramarao, N.; Tran, S.-L.; Marin, M.; Vidic, J. Advanced Methods for Detection of Bacillus cereus and Its Pathogenic Factors. Sensors 2020, 20, 2667. [Google Scholar] [CrossRef]
- Heyndrickx, M.; Scheldeman, P. Bacilli Associated with Spoilage in Dairy Products and Other Food. In Applications and Systematics of Bacillus and Relatives; Berkeley, R., Heyndrickx, M., Logan, N., De Vos, P., Eds.; Blackwell Science Ltd.: Malden, MA, USA, 2002; pp. 64–82. [Google Scholar]
- Zarb, P.; Coignard, B.; Griskeviciene, J.; Muller, A.; Vankerckhoven, V.; Weist, K. The European Centre for Disease Prevention and Control (ECDC) Pilot Point Prevalence Survey of Healthcare-Associated Infections and Antimicrobial Use. Eurosurveillance 2012, 17, 20316. [Google Scholar] [CrossRef]
- Enemchukwu, C.M.; Oli, A.N.; Okoye, E.I.; Ujam, N.T.; Osazuwa, E.O.; Emechebe, G.O.; Okeke, K.N.; Ifezulike, C.C.; Ejiofor, O.S.; Okoyeh, J.N. Winning the War against Multi-Drug Resistant Diarrhoeagenic Bacteria. Microorganisms 2019, 7, 197. [Google Scholar] [CrossRef] [Green Version]
- Lone, A.; Mottawea, W.; Ait Chait, Y.; Hammami, R. Dual Inhibition of Salmonella enterica and Clostridium perfringens by New Probiotic Candidates Isolated from Chicken Intestinal Mucosa. Microorganisms 2021, 9, 166. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Nanda, P.K.; Pateiro, M.; Lorenzo, J.M. Lactic Acid Bacteria and Bacteriocins: Novel Biotechnological Approach for Biopreservation of Meat and Meat Products. Microorganisms 2022, 10, 2058. [Google Scholar] [CrossRef]
- Zapaśnik, A.; Sokołowska, B.; Bryła, M. Role of Lactic Acid Bacteria in Food Preservation and Safety. Foods 2022, 11, 1283. [Google Scholar] [CrossRef]
- Campbell, L.L.; Sniff, E.E. Nisin sensitivity of Bacillus coagulans. Appl. Microbiol. 1959, 7, 289–291. [Google Scholar] [CrossRef]
- Lucas, R.; Grande, M.J.; Abriouel, H.; Maqueda, M.; Omar, N.B.; Valdivia, E.; Gálvez, A. Application of the broad-spectrum bacteriocin enterocin AS-48 to inhibit Bacillus coagulans in canned fruit and vegetable foods. Food Chem. Toxicol. 2006, 44, 1774–1781. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Fu, Y.; Kong, L.; Shen, Q.; Liu, M. Production of a Class IIb Bacteriocin with Broad-spectrum Antimicrobial Activity in Lactiplantibacillus plantarum RUB1. Probiotics Antimicrob. Proteins 2021, 13, 1820–1832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhang, Y.; Li, L.; Jiang, X. Biosynthesis and Production of Class II Bacteriocins of Food-Associated Lactic Acid Bacteria. Fermentation 2022, 8, 217. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Azmi, T.; Ibrahim, T. Fermentation factors influencing the production of bacteriocins by lactic acid bacteria. RSC Adv. 2017, 7, 29395–29420. [Google Scholar] [CrossRef]
- Lajis, A.F.B. Biomanufacturing process for the production of bacteriocins from Bacillaceae family. Bioresour. Bioprocess. 2020, 7, 8. [Google Scholar] [CrossRef]
- Abo-Amer, A.E. Optimization of bacteriocin production by Lactobacillus acidophilus AA11, a strain isolated from Egyptian cheese. Ann. Microbiol. 2011, 61, 445–452. [Google Scholar] [CrossRef]
- Picard, F.; Dressaire, C.; Girbal, L.; Cocaign-Bousquet, M. Examination of post-transcriptional regulations in prokaryotes by integrative biology. Comptes Rendus Biol. 2009, 332, 958–973. [Google Scholar] [CrossRef]
- Mavrič, A.; Tompa, G.; Trmcic, A.; Rogelj, I.; Matijašić, B.B. Bacteriocins of Lactobacillus gasseri K7—Monitoring of gassericin K7 A and B genes’ expression and isolation of an active component. Proc. Biochem. 2014, 49, 1251–1259. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Expression of Lactobacillus pentosus B96 bacteriocin genes under saline stress. Food Microbiol 2011, 28, 1339–1344. [Google Scholar] [CrossRef]
- Hurtado, A.; Ben Othman, N.; Chammem, N.; Hamdi, M.; Ferrer, S.; Reguant, C.; Bordons, A.; Rozès, N. Characterization of Lactobacillus isolates from fermented olives and their bacteriocin gene profiles. Food Microbiol. 2011, 28, 1514–1518. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Papadelli, M.; Pardali, E.; Mataragas, M.; Drosinos, E.H. Evaluation of plantaricin genes expression during fermentation of Raphanus sativus roots with a plantaricin-producing Lactobacillus plantarum starter. Curr. Microbiol. 2019, 76, 909–916. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Strains | Origin |
|---|---|---|
| Lactobacillus acidophilus | VKPM B-2105 | Fermented dairy product |
| Lactobacillus acidophilus | VKPM B-2900 | Human |
| Lactobacillus acidophilus | VKPM B-6551 | Human |
| Lactobacillus acidophilus | VKPM B-6552 | Human |
| Lactobacillus buchneri | VKPM B-7641 | Tomato pulp |
| Lactobacillus delbrueckii subsp. bulgaricus | VKPM B-11948 | Bulgarian yoghurt |
| Lactobacillus casei | VKPM B-2872 | Human |
| Lactobacillus casei | VKPM B-2873 | Human |
| Lactobacillus fermentum | VKPM B-8724 | Fermented beets |
| Lactobacillus gallinarum | VKPM B-10907 | Chicken crop |
| Lactobacillus graminis | VKPM B-11233 | Grass silage |
| Lactobacillus paracasei | VKPM B-4079 | Fermented sugar beet tops |
| Lactobacillus paracasei | VKPM B-11657 | Human |
| Lactobacillus plantarum | VKPM B-11007 | Lactobacterin |
| Lactobacillus rhamnosus | VKPM B-8238 | Infant |
| Lactobacillus reuteri | VKPM B-9448 | Human |
| Lactobacillus sakei | VKPM B-8936 | Italian dry-cured sausage |
| Lactobacillus salivarius | VKPM B-2214 | Human |
| No | Product | Gene | Primer Sequence (5′-3′) | Size, bp | Reference |
|---|---|---|---|---|---|
| 1. | Acidocin 8912 | acdT | F: AAGAATTAGCATTAATTTCTGGGGG | 96 | This study |
| R: CAGTATAACGAAGGCTTTCCCA | |||||
| 2. | Acidocin B | acdB | F: GCCACAGCGAACATTTATTGGA | 131 | This study |
| R: ACGCCCAAAATAGCAGCAAAG | |||||
| 3. | Bacteriocin BacSJ2-8 | bacSJ2-8 | F: ACTACCCCAGCCATCTCCAA | 90 | This study |
| R: TGGAGAGACAAGAGGGGTCA | |||||
| 4. | Gassericin T | gasT | F: GGAGTAGGTGGAGCGACAGT | 126 | This study |
| R: TCCACCAGTAGCTGCCGTTA | |||||
| 5. | Helveticin J | hlvJ | F: TCAAACAAACCAAAGTGACC | 440 | This study |
| R: CAAGTTGGTGCAGTAAATGGTG | |||||
| 6. | Helveticin homolog | Hlv | F: CCATTTGTTCGCCATACCAGCA | 198 | This study |
| R: CCACACCATCATTCAGCCGTTC | |||||
| 7. | Lactococcin 972 family | LacC0470_08940 | F: GCTTATACAGTTGATGTGCAAGGT | 128 | This study |
| R: CTTGATGCGCTGTATGCTCC | |||||
| 8. | Lactococcin 972 family | LG542_08235 | F: TTTCTTACACCAGCACCCCATC | 100 | This study |
| R: ATTCGGGATTCAGTGGAACAAAAG | |||||
| 9. | PapA | papA | F: CTTGTGGCAAACATTCCTGCT | 95 | This study |
| R: CTTGATGTCCACCAGTAGCCC | |||||
| 10. | Plantaricin A | plnA | F: AGCAACTTAGTAATAAGGAAATGCAAA | 102 | [14] |
| R: ACAGTTTCTTTACCTGTTTAATTGCAG | |||||
| 11. | Plantaricin E | plnE | F: ATACCACGAATGCCTGCAAC | 93 | This study |
| R: ATCTGGTGGTTTTAATCGGGG | |||||
| 12. | Plantaricin F | plnF | F: CTAATGACCCAATCGGCAGG | 94 | This study |
| R: ATGCTATTTCAGGTGGCGTTTT | |||||
| 13. | Plantaricin J | plnJ | F: ATAATAAGTTGAACGGGGTTGTTGG | 90 | This study |
| R: TGCCAGCTTCGCCATCATAAA | |||||
| 14. | Plantaricin K | plnK | F: TAATCCCTTGAACCACCAAGCA | 124 | This study |
| R: TAACTGCTGACGCTGAAAAGAA | |||||
| 15. | Sakacin A | sakA | F: TTCCAGCTAAACCACTAGCCC | 82 | This study |
| R: AAATGTTGGGTAAATCGGGGTG | |||||
| 16. | Sakacin P | sppA | F: AACAGCAATTACAGGTGGAAAA | 150 | This study |
| R: TATTTATTCCAGCCAGCGTTTC | |||||
| 17. | Sakacin T alpha | sakTα | F: AGAAGAATTGGTACTTGTAGTCGGT | 90 | This study |
| R: CCTGCTCCTGTACCAGCAATAC | |||||
| 18. | Sakacin T beta | sakTβ | F: TAATTGGGGATCAGTCGTGGG | 104 | This study |
| R: GTCCTGCACCGACTAAGCATC | |||||
| 19. | Sakacin Q | sakQ | F: TTGGTAAATGTGTAGTTGGTGCTTG | 81 | This study |
| R: CCATTCCCCAGAGACCACCA | |||||
| 20. | Sakacin X | sakX | F: TTGTCGGGGGAAAATACTACGG | 126 | This study |
| R: CAGCTCCACCGGTAGTCAAA | |||||
| 21. | Salivaricin ABP-118 α | abp118α | F: GCAAAGGTTGATGGTGGGAAAC | 145 | This study |
| R: TAAGTGCTCCGCCTACCATTC | |||||
| 22. | class II b bacteriocin | LRHK_2402 | F: GATCGTTCCCATCCTTGCTTG | 95 | This study |
| R: ATCCCTGTCGCTGCTATTCTTG | |||||
| 23. | class II b bacteriocin | LCA12A_2788 | F: AATTAGTGCTGAGACACAAGGA | 154 | This study |
| R: GTCTTAATCCAAGGAGGAGCCA | |||||
| 24. | Class I-Type I Lantibiotic | lanB | F: TATGATCGAGAARYAKAWAGATATGG | ~250 | [15] |
| R: TTATTAIRCAIATGIAYDAWACT | |||||
| 25. | Class I-Type I Lantibiotic | lanC | F: TAATTTAGGATWISYIMAYGG | ~400 | [15] |
| R: ACCWGKIIIICCRTRRCACCA | |||||
| 26. | Class I-Type II Lantibiotic | lanM | F: ATGCWAGWYWTGCWCATGG | 330 | [15] |
| R: CCTAATGAACCRTRRYAYCA |
| Function | Product | Gene | Primer Sequence (5′-3′) | Size, bp | Reference |
|---|---|---|---|---|---|
| Housekeeping genes | 16S rRNA | 16S rRNA | F: ACTCCTACGGGAGGCAGCAG | 200 | [16] |
| R: GTATTACCGCGGCTGCTGG | |||||
| elongation factor Tu | Ef-Tu | F: TCGAYGCTGCWCCDGAAGA | 194 | [17] | |
| R: TGGCATWGGRCCATCAGTWG | |||||
| Bacteriocin-related gene | class II b bacteriocin | LCA12A_2788 | F: ATTAGTGCTGAGACACAAGGA | 154 | This study |
| R: TCTTAATCCAAGGAGGAGCCA |
| Test Strains | Diameter of Growth Inhibition Zone ± SD (mm) | ||||
|---|---|---|---|---|---|
| L. acidophilus VKPM B-2105 | L. paracasei VKPM B-11657 | L. salivarius VKPM B-2214 | L. plantarum VKPM B-11007 | Control * | |
| B. megaterium VKPM B-3750 | 7.3 ± 0.6 | 13.8 ± 0.5 | 12.2 ± 0.5 | 7.2 ± 0.5 | 21.0 ± 0.9 |
| B. coagulans VKPM B-4521 | - *** | 14.0 ± 0.7 | 15.7 ± 0.6 | - | 12.0 ± 0.7 |
| B. coagulans VKPM B-9868 | 14.0 ± 1.0 | 11.7 ± 0.9 | 9.8 ± 0.9 | 7.5 ± 0.6 | 7.3 ± 0.5 |
| B. coagulans VKPM B-10468 | - | 7.2 ± 0.5 | 8.0 ± 0.7 | - | 11.8 ± 0.9 |
| B. cereus ATCC 9634 | - | - | - | - | 11.8 ± 0.9 |
| S. aureus ATCC 4330 | - | - | - | - | 10.7 ± 0.6 |
| S. epidermidis ATCC 12228 | 16.2 ± 0.5 | - | 7.2 ± 0.9 | - | 19.5 ± 1.2 |
| S. enterica VKPM B-5300 ** | 7.8 ± 0.9 | 7.5 ± 0.6 | 12.2 ± 0.5 | 7.3 ± 0.6 | 20.5 ± 1.1 |
| Test Strains | Diameter of Growth Inhibition Zone ± SD (mm) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L. paracasei VKPM B-11657 | L. salivarius VKPM B-2214 | RBEH | |||||||||
| MRS | RBEH | MRS | RBEH | before | |||||||
| 16 h | 24 h | 30 h | 16 h | 24 h | 16 h | 24 h | 30 h | 16 h | 24 h | Inoculation | |
| B. megaterium VKPMB-3750 | 11.7 ± 1.3 | 18.8 ± 1.0 | 18.0 ± 0.9 | 11.7 ± 0.9 | 8.7 ± 0.9 | 11.0 ± 0.9 | 20.8 ± 1.0 | 19.2 ± 0.8 | 7.5 ± 0.7 | 9.3 ± 0.5 | - |
| B. coagulans VKPM B-4521 | 12.8 ± 0.8 | 18.0 ± 0.7 | 17.2 ± 0.8 | 12.7 ± 0.5 | 11.2 ± 0.8 | 16.3 ± 1.1 | 25.0 ± 0.7 | 23.0 ± 0.7 | 11.7 ± 0.9 | 13.2 ± 0.8 | - |
| B. coagulans VKPM B-9868 | 10.0 ± 1.3 | 24.7 ± 0.9 | 24.0 ± 0.9 | 12.3 ± 0.9 | 9.7 ± 1.1 | 9.2 ± 0.8 | 27.0 ± 0.9 | 24.3 ± 0.9 | 10.7 ± 0.9 | 12.0 ± 0.7 | - |
| B. coagulans VKPM B-10468 | 7.5 ± 0.6 | 11.0 ± 0.9 | 9.7 ± 0.5 | 12.0 ± 0.9 | 10.0 ± 0.7 | 7.2 ± 0.4 | 11.0 ± 0.4 | 10.2 ± 0.4 | 11.5 ± 1.1 | 14.2 ± 0.8 | - |
| B. cereus ATCC 9634 | - * | - | - | - | - | - | - | - | - | - | - |
| S. aureus ATCC 4330 | - | - | - | - | - | - | - | - | - | - | - |
| S. epidermidis ATCC 14990 | - | - | - | 13.8 ± 1.0 | 13.3 ± 0.9 | 8.0 ± 0.7 | 11.8 ± 0.4 | 9.8 ± 0.8 | 8.7 ± 0.5 | 15.2 ± 0.8 | - |
| S. enterica VKPM B-5300 | 7.8 ± 0.8 | 10.2 ± 0.8 | - | 10.7 ± 0.9 | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Epishkina, J.M.; Romanova, M.V.; Chalenko, M.A.; Khromova, N.Y.; Karetkin, B.A.; Beloded, A.V.; Kornienko, M.A.; Averina, Y.M.; Shakir, I.V.; Panfilov, V.I. Evaluation of Rye Bran Enzymatic Hydrolysate Effect on Gene Expression and Bacteriocinogenic Activity of Lactic Acid Bacteria. Fermentation 2022, 8, 752. https://doi.org/10.3390/fermentation8120752
Epishkina JM, Romanova MV, Chalenko MA, Khromova NY, Karetkin BA, Beloded AV, Kornienko MA, Averina YM, Shakir IV, Panfilov VI. Evaluation of Rye Bran Enzymatic Hydrolysate Effect on Gene Expression and Bacteriocinogenic Activity of Lactic Acid Bacteria. Fermentation. 2022; 8(12):752. https://doi.org/10.3390/fermentation8120752
Chicago/Turabian StyleEpishkina, Julia M., Maria V. Romanova, Marina A. Chalenko, Natalya Yu. Khromova, Boris A. Karetkin, Andrey V. Beloded, Maria A. Kornienko, Yulia M. Averina, Irina V. Shakir, and Victor I. Panfilov. 2022. "Evaluation of Rye Bran Enzymatic Hydrolysate Effect on Gene Expression and Bacteriocinogenic Activity of Lactic Acid Bacteria" Fermentation 8, no. 12: 752. https://doi.org/10.3390/fermentation8120752