
Lactic Bacteria in Artisanal Cheese: Characterization through Metagenomics
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
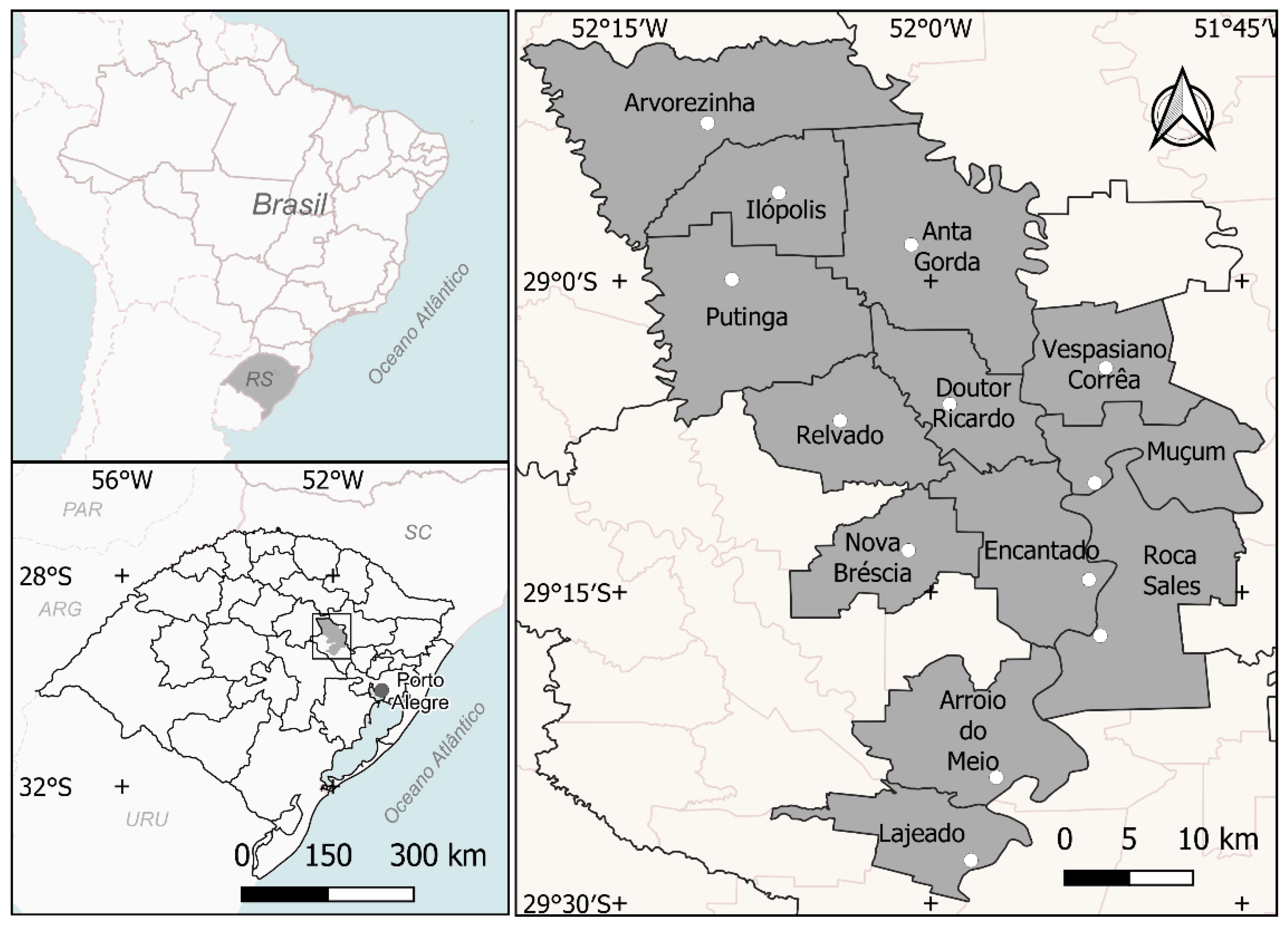
2.1. Characterization of the Region
2.2. Sample Collection
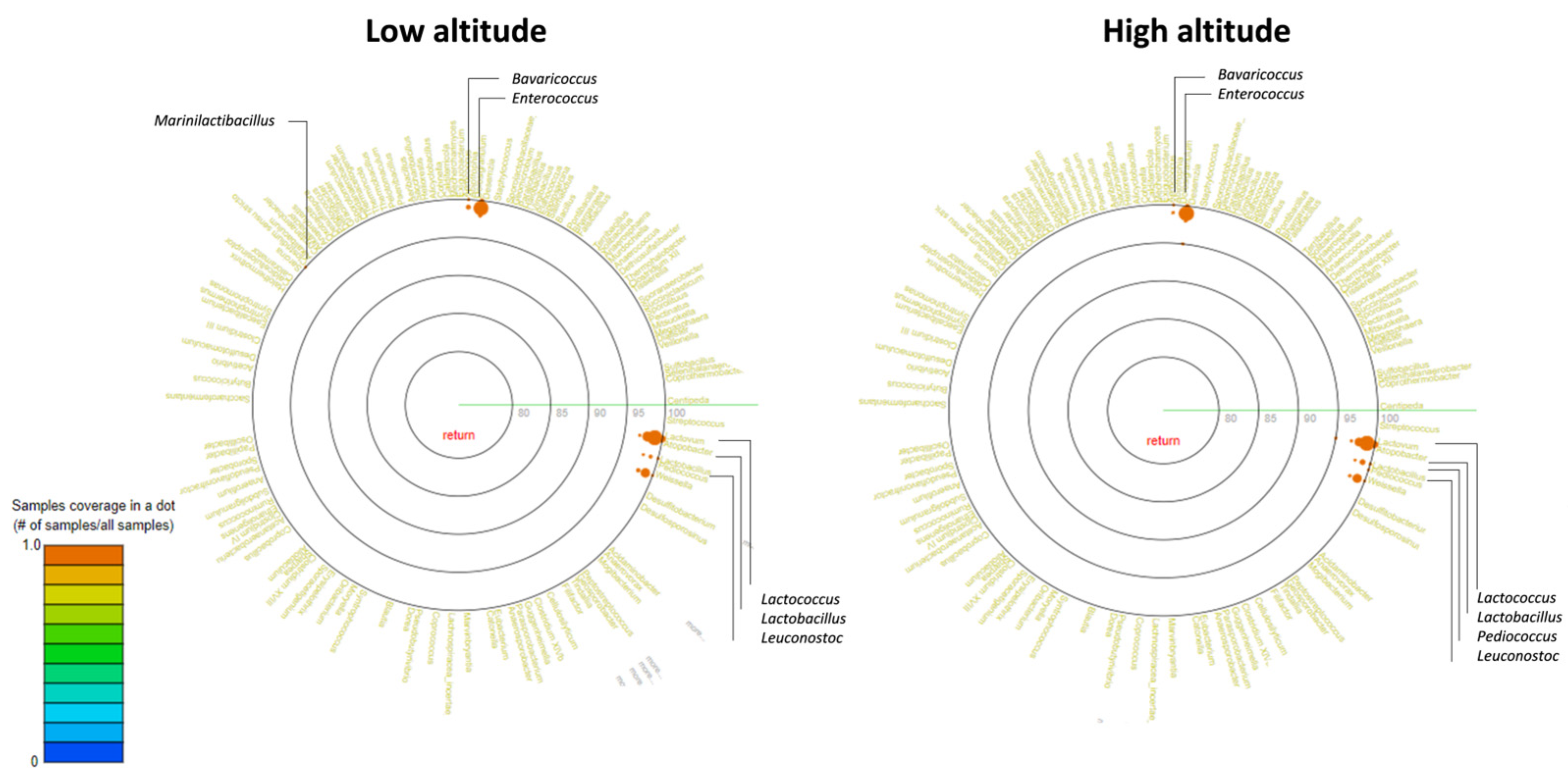
2.3. Metagenomic Analysis
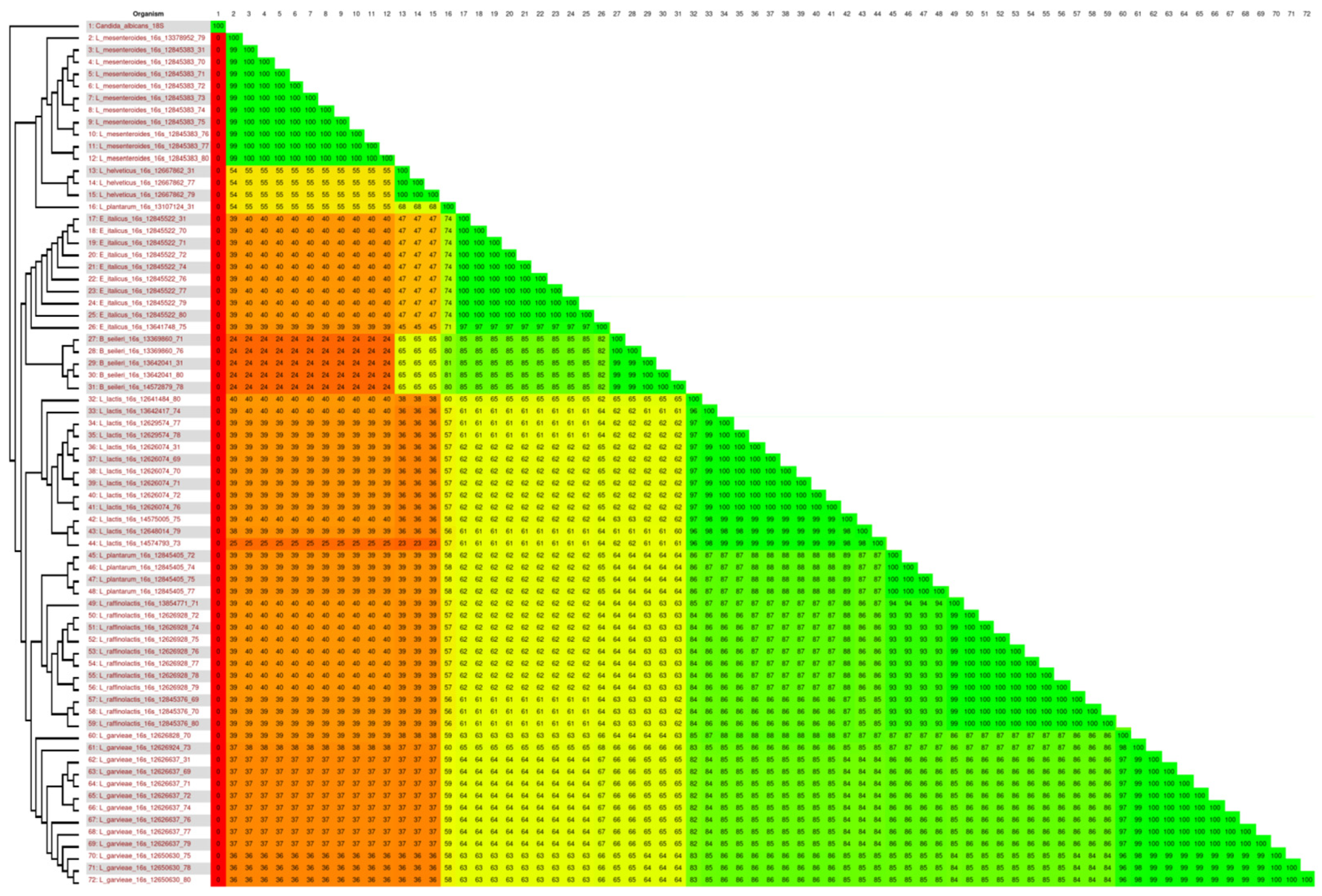
2.4. Phylogenetic Analysis
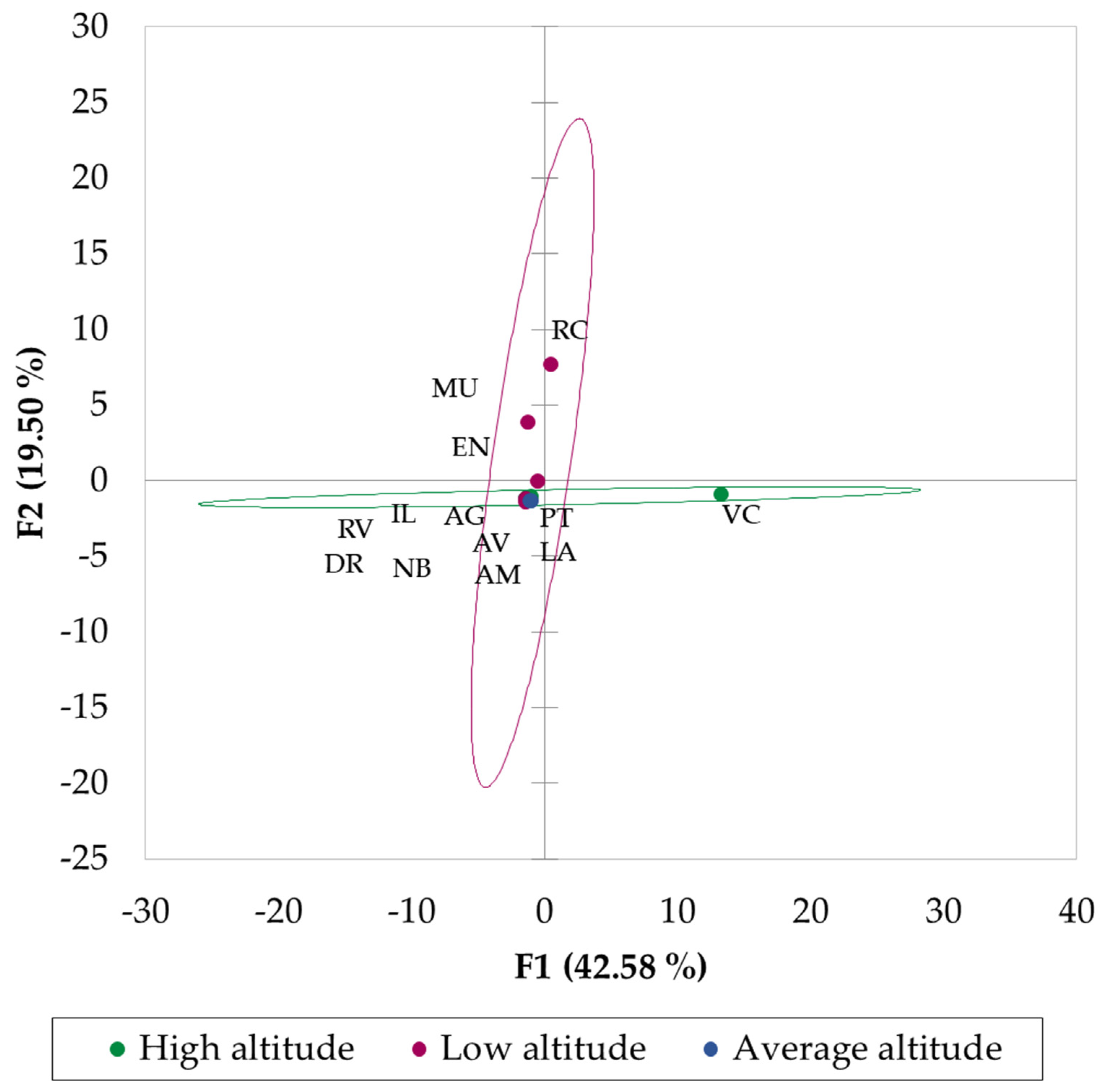
2.5. Statistical Analysis
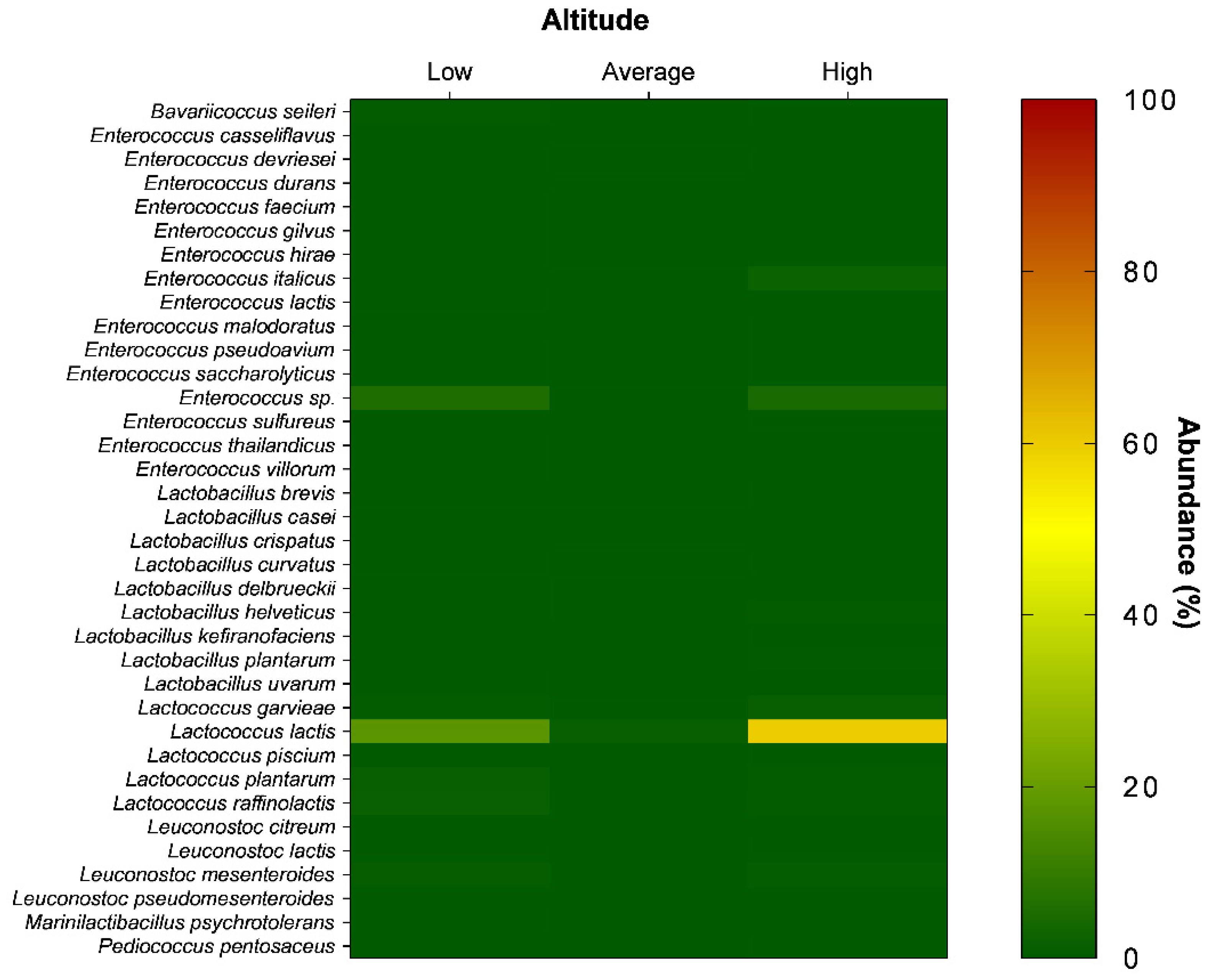
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montel, M.-C.; Buchin, S.; Mallet, A.; Delbes-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Kothe, C.I.; Mohellibi, N.; Renault, P. Revealing the microbial heritage of traditional Brazilian cheeses through metagenomics. Food Res. Int. 2022, 157, 111265. [Google Scholar] [CrossRef] [PubMed]
- Penna, A.; Gigante, M.; Todorov, S. Artisanal Brazilian Cheeses—History, Marketing, Technological and Microbiological Aspects. Foods 2021, 10, 1562. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, B.A.; Magnani, M.; Luciano, W.A.; Campagnollo, F.B.; Pimentel, T.C.; Alvarenga, V.O.; Pelegrino, B.O.; Cruz, A.G.; Sant’Ana, A.S. Brazilian Artisanal Cheeses: An Overview of their Characteristics, Main Types and Regulatory Aspects. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1636–1657. [Google Scholar] [CrossRef] [Green Version]
- Kamimura, B.A.; De Filippis, F.; Sant’Ana, A.S.; Ercolini, D. Large-scale mapping of microbial diversity in artisanal Brazilian cheeses. Food Microbiol. 2018, 80, 40–49. [Google Scholar] [CrossRef]
- Brasil. Decreto n° 11.099, de 21 de Junho de Regulamenta o Art. 10-A da Lei n° 1.283, de 18 de Dezembro de 1950, e a Lei n° 13.860, de 18 de Julho de 2019, Para Dispor Sobre a Elaboração e a Comercialização de Produtos Alimentícios de Origem Animal Produzidos de Forma Artesanal. Available online: http://www.planalto.gov.br/ccivil_03/_ato2019-2022/2022/decreto/D11099.htm#:~:text=DECRETO%20N%C2%BA%2011.099%2C%20DE%2021,animal%20produzidos%20de%20forma%20artesanal> (accessed on 3 November 2022).
- Brasil. Lei n° 13.860, de 18 de Julho de Dispõe Sobre a Elaboração e a Comercialização de Queijos Artesanais e dá Outras Providências. Available online: https://www.gov.br/agricultura/pt-br/assuntos/producao-animal/selo-arte-selo-queijo-artesanal/legislacao/lei-no-13-860-de-18-de-julho-de-2019.pdf/view> (accessed on 28 December 2022).
- Margalho, L.P.; Feliciano, M.D.; Silva, C.E.; Abreu, J.S.; Piran, M.V.F.; Sant’Ana, A.S. Brazilian artisanal cheeses are rich and diverse sources of nonstarter lactic acid bacteria regarding technological, biopreservative, and safety properties—Insights through multivariate analysis. J. Dairy Sci. 2020, 103, 7908–7926. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Button, J.E.; Santarelli, M.; Dutton, R.J. Cheese Rind Communities Provide Tractable Systems for In Situ and In Vitro Studies of Microbial Diversity. Cell 2014, 158, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Dugat-Bony, E.; Garnier, L.; Denonfoux, J.; Ferreira, S.; Sarthou, A.-S.; Bonnarme, P.; Irlinger, F. Highlighting the microbial diversity of 12 French cheese varieties. Int. J. Food Microbiol. 2016, 238, 265–273. [Google Scholar] [CrossRef]
- De Antônio, M.B.; Borelli, B. A importância das bactérias láticas na segurança e qualidade dos queijos minas artesanais. Rev. Inst. Laticínios Cândido Tostes 2020, 75, 204–221. [Google Scholar] [CrossRef]
- Delcenserie, V.; Taminiau, B.; Delhalle, L.; Nezer, C.; Doyen, P.; Crevecoeur, S.; Roussey, D.; Korsak, N.; Daube, G. Microbiota characterization of a Belgian protected designation of origin cheese, Herve cheese, using metagenomic analysis. J. Dairy Sci. 2014, 97, 6046–6056. [Google Scholar] [CrossRef]
- Dimov, S.G. The unusual microbiota of the traditional Bulgarian dairy product Krokmach—A pilot metagenomics study. Int. J. Dairy Technol. 2021, 75, 139–149. [Google Scholar] [CrossRef]
- Papadakis, P.; Konteles, S.; Batrinou, A.; Ouzounis, S.; Tsironi, T.; Halvatsiotis, P.; Tsakali, E.; Van Impe, J.F.M.; Vougiouklaki, D.; Strati, I.F.; et al. Characterization of Bacterial Microbiota of P.D.O. Feta Cheese by 16S Metagenomic Analysis. Microorganisms 2021, 9, 2377. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Anastasiou, R.; Georgalaki, M.; Bounenni, R.; Paximadaki, A.; Charmpi, C.; Alexandraki, V.; Kazou, M.; Tsakalidou, E. Comparison of the Microbiome of Artisanal Homemade and Industrial Feta Cheese through Amplicon Sequencing and Shotgun Metagenomics. Microorganisms 2022, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- Planý, M.; Kuchta, T.; Šoltýs, K.; Szemes, T.; Pangallo, D.; Siekel, P. Metagenomic Analysis of Slovak Bryndza Cheese Using Next-Generation 16S rDNA Amplicon Sequencing. Nova Biotechnol. Chim. 2016, 15, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Lahtinen, S.; Ouwehand, A.C.; Salminen, S.; Von Wright, A. (Eds.) Lactic Acid Bacteria: Microbiological and Functional Aspects; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Sant’Anna, F.M.; Wetzels, S.U.; Cicco, S.H.S.; Figueiredo, R.C.; Sales, G.A.; Figueiredo, N.C.; Nunes, C.A.; Schmitz-Esser, S.; Mann, E.; Wagner, M.; et al. Microbial shifts in Minas artisanal cheeses from the Serra do Salitre region of Minas Gerais, Brazil throughout ripening time. Food Microbiol. 2019, 82, 349–362. [Google Scholar] [CrossRef]
- Resende, M.F.S.; Costa, H.H.S.; Andrade, E.H.P.; Acúrcio, L.B.; Drummond, A.F.; Cunha, A.F.; Souza, M.R. Queijo de minas artesanal da Serra da Canastra: Influência da altitude das queijarias nas populações de bactérias acidolácticas. Arq. Bras. Med. Vet. Zoo. 2011, 63, 1567–1573. [Google Scholar] [CrossRef]
- Topographic. 2022. Available online: https://pt-br.topographic-map.com/map-pb651/Rio-Grande-do-Sul/ (accessed on 14 October 2022).
- Wang, Y.; Qian, P.Y. Conservative Fragments in Bacterial 16S rRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Agren, J.; Sundström, A.; Håfström, T.; Segerman, B. Gegenees: Fragmented alignment of multiple genomes for determining phylogenomic distances and genetic signatures unique for specified target groups. PLoS ONE 2012, 7, e39107. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Kloepper, T.; Bryant, D. SplitsTree 4.0-Computation of phylogenetic trees and networks. Bioinformatics 2008, 14, 68–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Maruyama, T.; Yano, M.; Yamada, T.; Kurokawa, K. VITCOMIC2: Visualization tool for the phylogenetic composition of microbial communities based on 16S rRNA gene amplicons and metagenomic shotgun sequencing. BMC Syst. Biol. 2018, 12, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Margalho, L.P.; Kamimura, B.A.; Brexó, R.P.; Alvarenga, V.O.; Cebeci, A.S.; Janssen, P.W.; Dijkstra, A.; Starrenburg, M.J.; Sheombarsing, R.S.; Cruz, A.G.; et al. High throughput screening of technological and biopreservation traits of a large set of wild lactic acid bacteria from Brazilian artisanal cheeses. Food Microbiol. 2021, 100, 103872. [Google Scholar] [CrossRef]
- Gezginc, Y.; Karabekmez-Erdem, T.; Tatar, H.D.; Dağgeçen, E.C.; Ayman, S.; Akyol, İ. Metagenomics and volatile profile of Turkish artisanal Tulum cheese microbiota. Food Biosci. 2022, 45, 101497. [Google Scholar] [CrossRef]
- Coeuret, V.; Dubernet, S.; Bernardeau, M.; Gueguen, M.; Vernoux, J.P. Isolation, characterisation and identification of lactobacilli focusing mainly on cheeses and other dairy products. Le Lait 2003, 83, 269–306. [Google Scholar] [CrossRef]
- Amraii, H.N.; Abtahi, P.; Jafari, H.R.; Mohajerani, M.R.; Fakhroleslam, N. In vitro study of potentially probiotic lactic acid bacteria strains isolated from traditional dairy products. Jundishapur J. Microbiol. 2014, 7, e10168. [Google Scholar]
- Suárez, N.; Weckx, S.; Minahk, C.; Hebert, E.; Saavedra, L. Metagenomics-based approach for studying and selecting bioprotective strains from the bacterial community of artisanal cheeses. Int. J. Food Microbiol. 2020, 335, 108894. [Google Scholar] [CrossRef]
- Dimov, S.G.; Gyurova, A.; Zagorchev, L.; Dimitrov, T.; Georgieva-Miteva, D.; Peykov, S. NGS-Based Metagenomic Study of Four Traditional Bulgarian Green Cheeses from Tcherni Vit. Lwt 2021, 152, 112278. [Google Scholar] [CrossRef]
- Myazaki, N.L. Identificação e Quantificação de Bactérias em Queijo Colonial Utilizando as Regiões V3–V4 do Gene 16S Rrna; TCC, Departamento de Ciência e Tecnologia de Alimentos, Universidade Federal de Santa Catarina: Florianópolis, Brazil, 2021; 80p. [Google Scholar]
- Ishikawa, M.; Kodama, K.; Yasuda, H.; Okamoto-Kainuma, A.; Koizumi, K.; Yamasato, K. Presence of halophilic and alkaliphilic lactic acid bacteria in various cheeses. Lett. Appl. Microbiol. 2006, 44, 308–313. [Google Scholar] [CrossRef]
- Roth, E.; Miescher Schwenninger, S.; Hasler, M.; Eugster-Meier, E.; Lacroix, C. Population dynamics of two antilisterial cheese surface consortia revealed by temporal temperature gradient gel electrophoresis. BMC Microbiol. 2010, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Nakajima, K.; Yanagi, M.; Yamamoto, Y.; Yamasato, K. Marinilactibacillus psychrotolerans gen. nov., sp. nov., a halophilic and alkaliphilic marine lactic acid bacterium isolated from marine organisms in temperate and subtropical areas of Japan. Int. J. Syst. Evol. Microbiol. 2003, 53, 711–720. [Google Scholar] [CrossRef]
- Jiang, S.; Cai, L.; Lv, L.; Li, L. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell Factories 2021, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Song, A.A.-L.; In, L.L.A.; Lim, S.H.E.; Rahim, R.A. A review on Lactococcus lactis: From food to factory. Microb. Cell Factories 2017, 16, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palles, T.; Beresford, T.; Condon, S.; Cogan, M.T. Citrate Metabolism in Lactobacillus plantarum. J. Appl. Microbiol. 1998, 85, 147–154. [Google Scholar] [CrossRef]
- Parsaeimehr, M.; Khazaei, M.; Jebellijavan, A.; Staji, H. The Isolation and Identification of Dominant Lactic Acid Bacteria by the Sequencing of the 16S rRNA in Traditional Cheese (Khiki) in Semnan, Iran. J. Human, Environ. Health Promot. 2019, 5, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Vermote, L.; Verce, M.; De Vuyst, L.; Weckx, S. Amplicon and shotgun metagenomic sequencing indicates that microbial ecosystems present in cheese brines reflect environmental inoculation during the cheese production process. Int. Dairy J. 2018, 87, 44–53. [Google Scholar] [CrossRef]
- Escobar-Zepeda, A.; Sanchez-Flores, A.; Quirasco Baruch, M. Metagenomic analysis of a Mexican ripened cheese reveals a unique complex microbiota. Food Microbiol. 2016, 57, 116–127. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
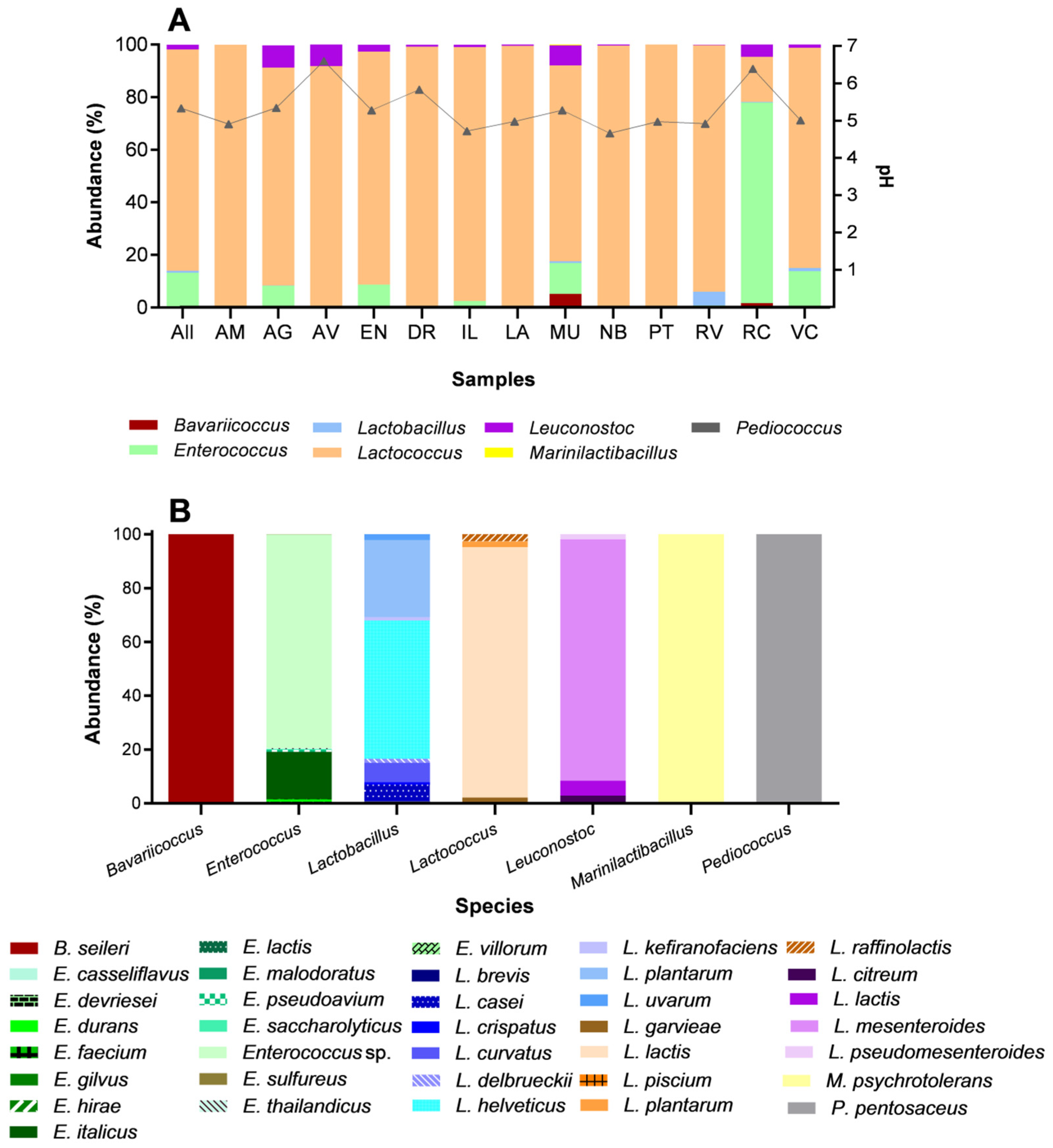
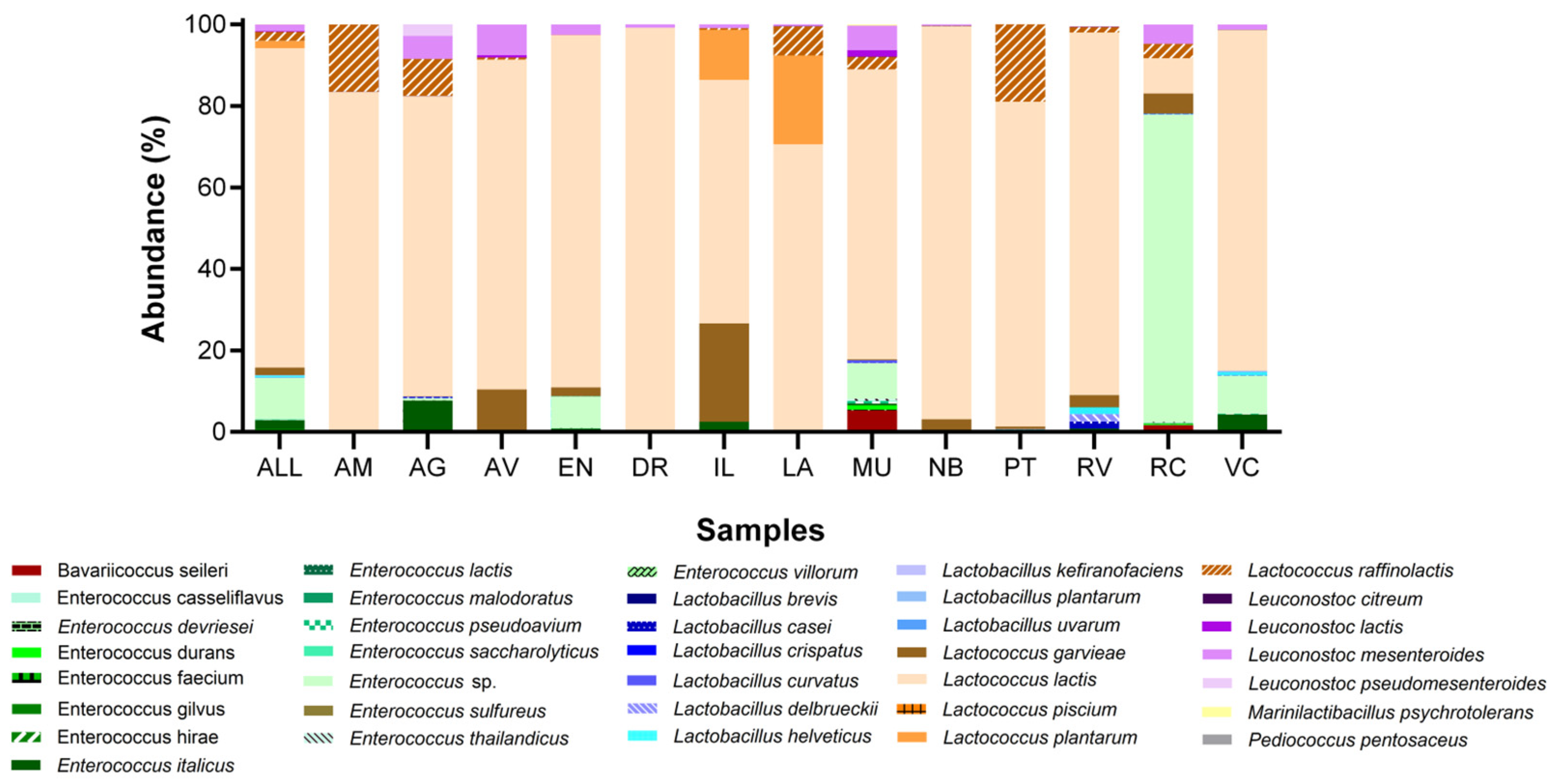
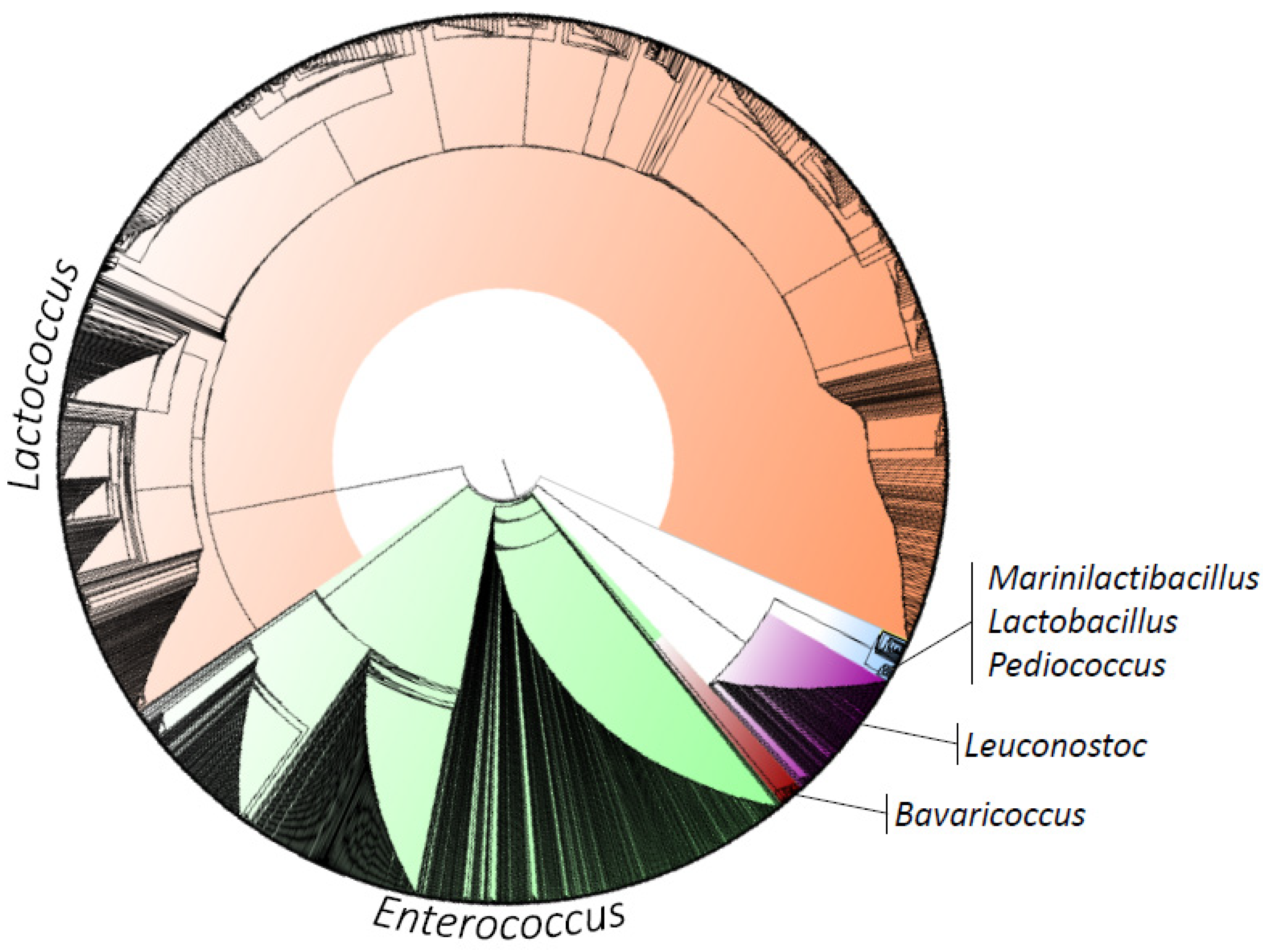
| Phylum | Class | Order | Family | Genus | Specie | Municipality | Bac Total (%) * | LAB Total (%) ** |
|---|---|---|---|---|---|---|---|---|
| Firmicutes | Bacilli | Lactobacillales | Enterococcaceae | Bavariicoccus | Bavariicoccus seileri | AV, MU, PT, RC, VC | 0.270 | 0.449 |
| Enterococcus | Enterococcus casseliflavus | AG, EN, IL, MU, PT, RC, VC | 0.014 | 0.023 | ||||
| E. devriesei | AM, EN, MU, NB, VC | 0.035 | 0.058 | |||||
| E. durans | AM, AG, EN, MU, RC, VC | 0.060 | 0.099 | |||||
| E. faecium | EN, MU, RC | 0.003 | 0.005 | |||||
| E. gilvus | MU | 0.003 | 0.005 | |||||
| E. hirae | EN, MU, RC | 0.002 | 0.002 | |||||
| E. italicus | AG, AV, EN, IL, LA, MU, NB, RV, RC, VC | 1.351 | 2.251 | |||||
| E. lactis | MU, RC | 0.002 | 0.002 | |||||
| E. malodoratus | VC | 0.003 | 0.005 | |||||
| E. pseudoavium | AM, EN, MU, RC, VC | 0.055 | 0.092 | |||||
| E. saccharolyticus | EN, MU, RC, VC | 0.022 | 0.036 | |||||
| E. sulfureus | VC | 0.001 | 0.002 | |||||
| E. thailandicus | MU, RC | 0.025 | 0.041 | |||||
| E. villorum | MU, RC | 0.002 | 0.002 | |||||
| E. sp. | AM, AG, AV, EN, DR, LA, MU, PT, RC VC | 6.102 | 10.168 | |||||
| Lactobacillaceae | Lactobacillus | Lactobacillus brevis | VC | 0.003 | 0.005 | |||
| L. casei | AG, CR, MU, RV, RC, VC | 0.026 | 0.043 | |||||
| L. crispatus | AG, AV, IL, RC, VC | 0.004 | 0.006 | |||||
| L. curvatus | MU, NB, VC | 0.030 | 0.050 | |||||
| L. delbrueckii | NB, RV | 0.006 | 0.010 | |||||
| L. helveticus | NB, RV, VC | 0.217 | 0.361 | |||||
| L. kefiranofaciens | DR, PT | 0.005 | 0.008 | |||||
| L. plantarum | VC | 0.121 | 0.201 | |||||
| L. uvarum | MU, RC | 0.009 | 0.014 | |||||
| Pediococcus | Pediococcus pentosaceus | VC | 0.002 | 0.003 | ||||
| Streptococcaceae | Lactococcus | Lactococcus garvieae | AM, AG, AV, EN, DR, IL, LA, MU, NB, PT, RV, RC, VC | 1.078 | 1.797 | |||
| L. lactis | AM, AG, AV, EN, DR, IL, LA MU, NB, PT, RV, RC, VC | 47.041 | 78.383 | |||||
| L. piscium | IL | 0.001 | 0.002 | |||||
| L. plantarum | EN, IL, LA, NB, | 1.068 | 1.779 | |||||
| L. raffinolactis | AM, AG, AV, EM, DR, IL, LA, MU, NB, PT, RV, RC | 1.350 | 2.250 | |||||
| Leuconostocaceae | Leuconostoc | Leuconostoc citreum | IL, LA, MU, RV, RC, VC | 0.032 | 0.053 | |||
| L. lactis | AV, LA, MU | 0.059 | 0.098 | |||||
| L. mesenteroides | AG, AV, EN, DR, IL, LA, MU, NB, RV, RC, VC | 0.980 | 1.632 | |||||
| L. pseudomesenteroides | AG, EN, LA, MU, RC, VC | 0.021 | 0.034 | |||||
| Carnobacteriaceae | Marinilactibacillus | Marinilactibacillus psychrotolerans | MU | 0.010 | 0.016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erhardt, M.M.; Oliveira, W.d.C.; Fröder, H.; Marques, P.H.; Oliveira, M.B.P.P.; Richards, N.S.P.d.S. Lactic Bacteria in Artisanal Cheese: Characterization through Metagenomics. Fermentation 2023, 9, 41. https://doi.org/10.3390/fermentation9010041
Erhardt MM, Oliveira WdC, Fröder H, Marques PH, Oliveira MBPP, Richards NSPdS. Lactic Bacteria in Artisanal Cheese: Characterization through Metagenomics. Fermentation. 2023; 9(1):41. https://doi.org/10.3390/fermentation9010041
Chicago/Turabian StyleErhardt, Magnolia Martins, Wemerson de Castro Oliveira, Hans Fröder, Pedro Henrique Marques, Maria Beatriz Prior Pinto Oliveira, and Neila Silvia Pereira dos Santos Richards. 2023. "Lactic Bacteria in Artisanal Cheese: Characterization through Metagenomics" Fermentation 9, no. 1: 41. https://doi.org/10.3390/fermentation9010041