
Evaluation of the Thermal Processes on Changing the Phenotypic Characteristics of Escherichia coli Strains from Ice Cream Compared to Non-Pasteurized Milk

Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling
2.2. Livestock Milk Sampling
2.3. Heat Shock in Bacteria Isolated from Raw Milk
2.4. Phenotypic Tests
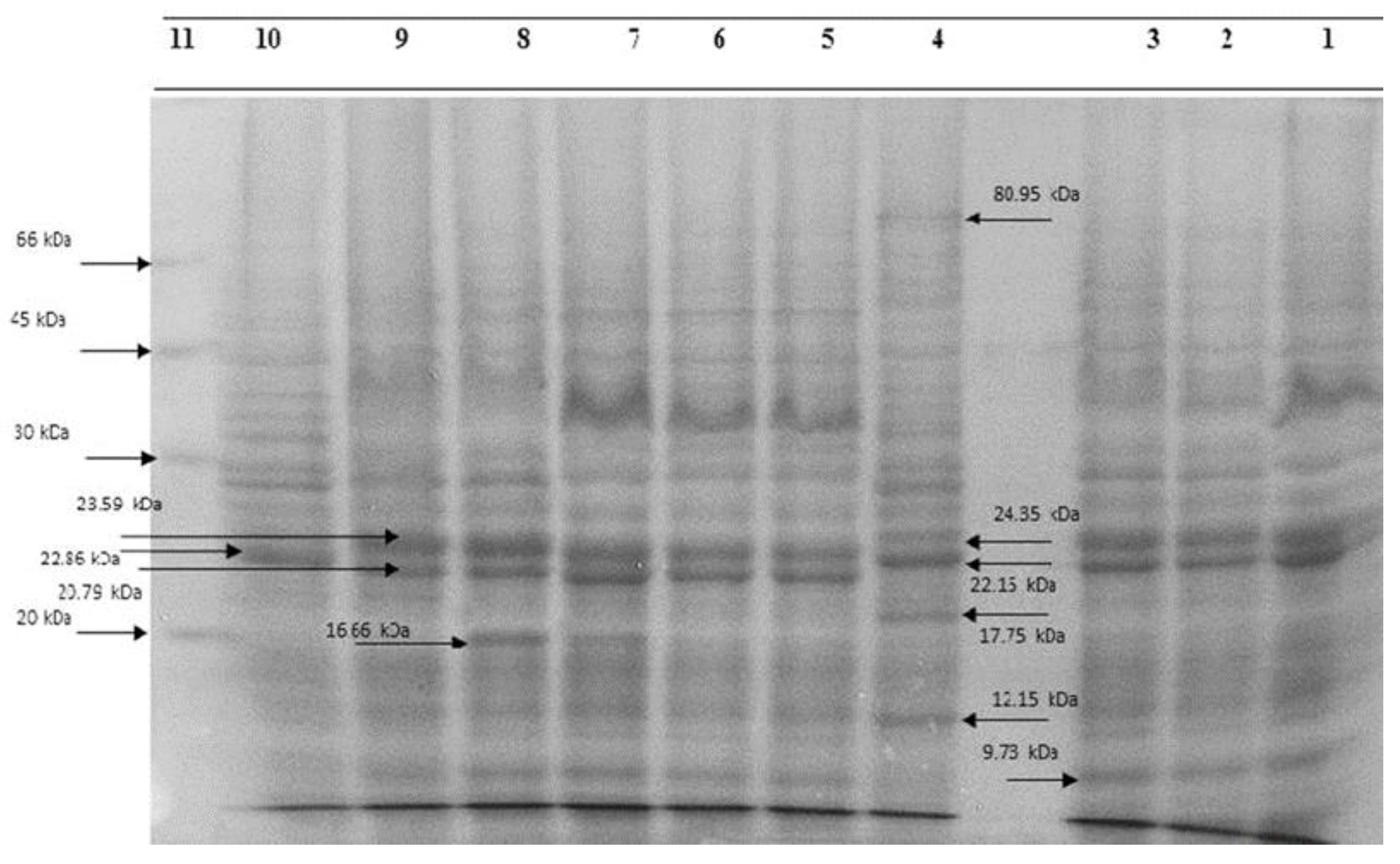
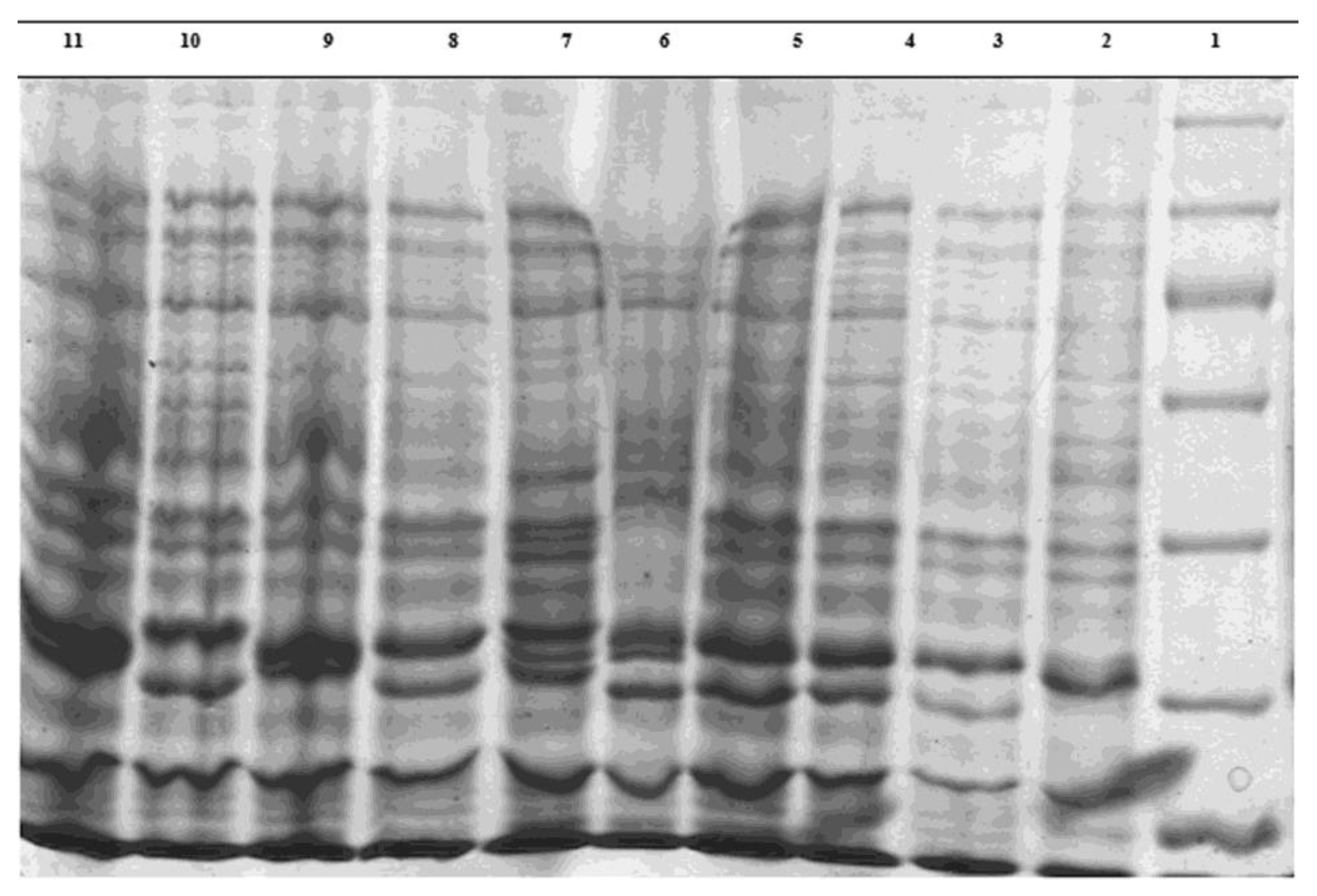
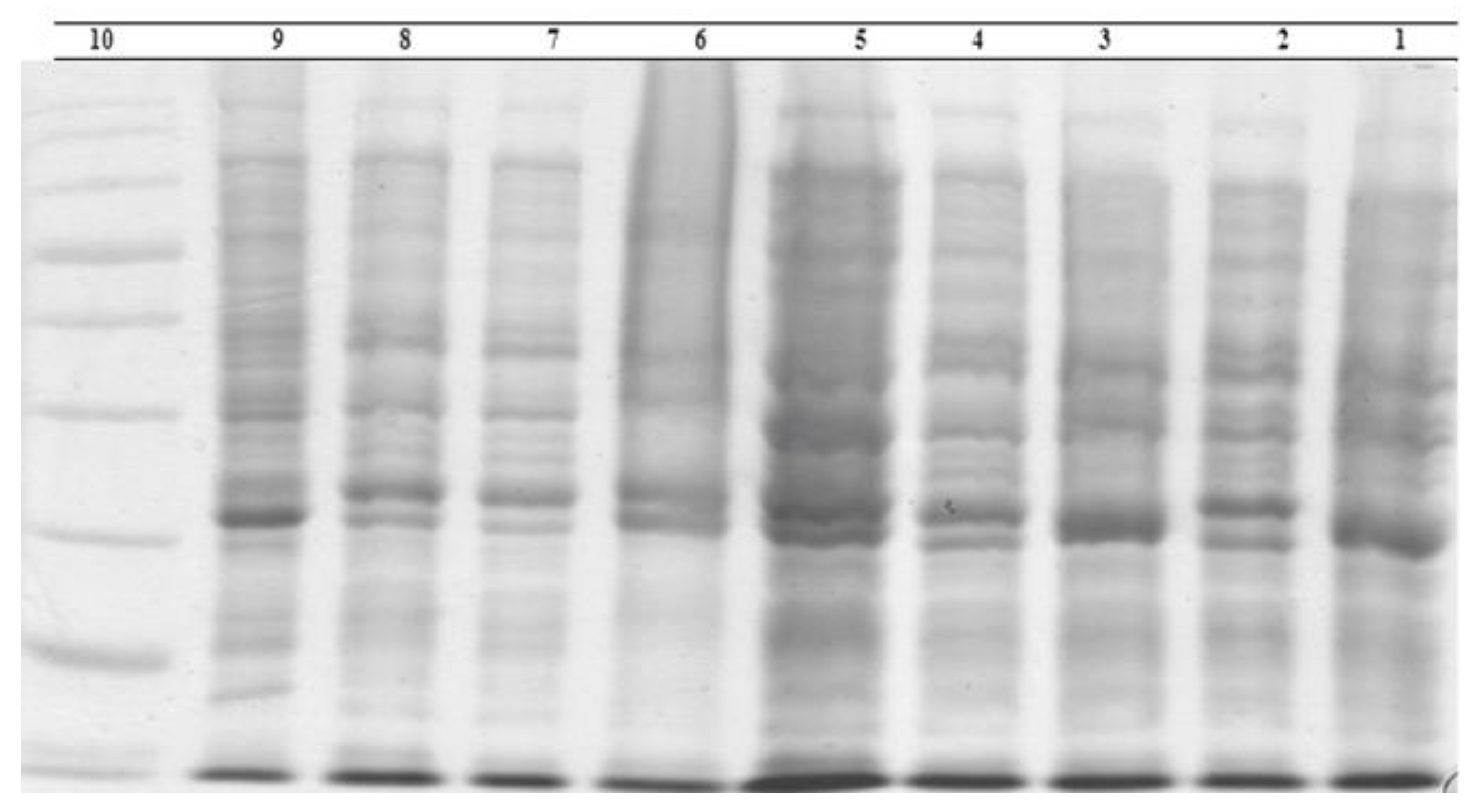
2.5. Whole-Bacteria Protein Electrophoresis
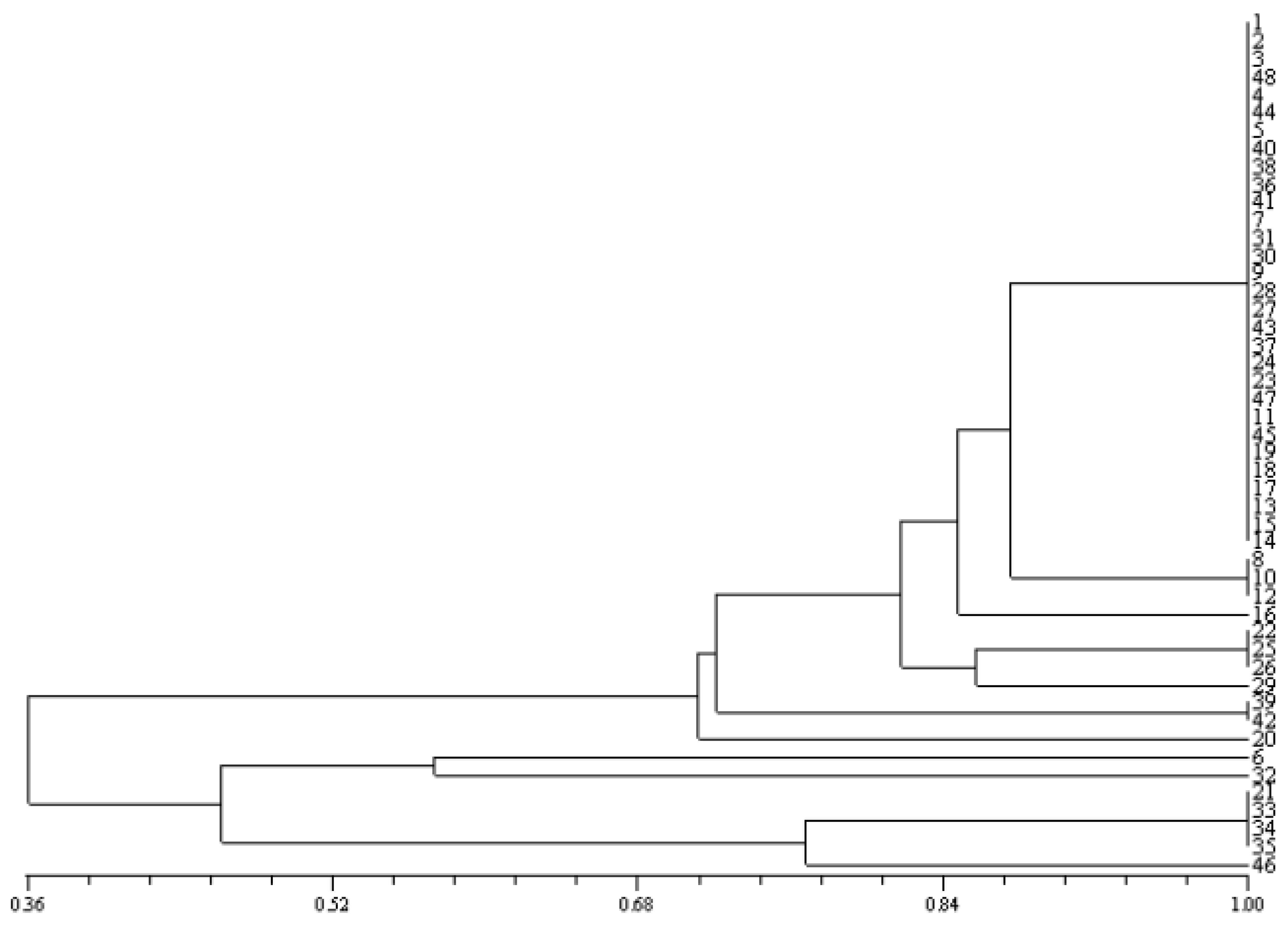
2.6. Calculating the Percentage of Similarity
2.7. Investigating E. coli Strains via Sequencing of 16S rDNA
2.7.1. DNA Extraction
2.7.2. PCR Method
2.7.3. Agarose Gel Electrophoresis and PCR Product Purification
3. Results
3.1. Phenotypic Tests of Isolated Coliforms
3.2. Electrophoresis of Bacteria Proteins
3.3. Sequencing of 16S rDNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erjavec, M.S. Introductory Chapter: The versatile Escherichia coli. In The Universe of Escherichia coli; Intech Open: Rijeka, Croatia, 2019; pp. 1–19. [Google Scholar]
- Tortorello, M.L. Indicator organisms for safety and quality-uses and methods for detection: Mini review. J. AOAC Int. 2003, 86, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Quigley, L.; OSullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F. The complex microbiota of raw milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, M.A.; Motazzim-ul-Haque, H.M.; Noor, R. Isolation and identification of pathogenic Escherichia coli, Klebsiella spp. and Staphylococcus spp. in raw milk samples collected from different areas of Dhaka city, Bangladesh. Stamford J. Microbiol. 2011, 1, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Pourmahmoodi, A.; Mohammadi, j.; Mirzai, A.; Momeni-Negad, M.; Afshar, R. Epidemiological study of traditional ice cream in Yasuj. Armaghan Danesh 2002, 8, 59–65. [Google Scholar]
- Motarjemi, Y.; Moy, G.G.; Jooste, P.J.; Anelich, L.E. Chapter 5—Milk and dairy products. In Food Safety Management; Motarjemi, Y., Lelieveld, H., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 83–117. [Google Scholar]
- Domenech, E.; Amoros, J.A.; Escriche, I. Effectiveness of prerequisites and the HACCP plan in the control of microbial contamination in ice cream and cheese companies. Foodborne Pathog. Dis. 2013, 10, 222–228. [Google Scholar] [CrossRef]
- Jooste, P.J.; Anelich, L.; Motarjemi, Y. Safety of food and beverages: Milk and dairy products. In Encyclopedia of Food Safety; Motarjemi, Y., Ed.; Academic Press: Waltham, MA, USA, 2014; p. 2304. [Google Scholar]
- Hosseini-Jazani, N.; Hadizadeh, O.; Farzaneh, H.; Moloudizargari, M. Synergistic antibacterial effects of β-Chloro-L-alanine and phosphomycin on urinary tract isolates of E. coli. BJM 2013, 1, 1–6. [Google Scholar]
- Ori, E.; Takagi, E.; Andrade, T.; Miguel, B.; Cergole-Novella, M.; Guth, B.; Camargo, C. Diarrhoeagenic Escherichia coli and Escherichia albertii in Brazil: Pathotypes and serotypes over a 6-year period of surveillance. Epidemiol. Infect. 2019, 147, e10. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Bushlaibi, M.; Alrefaei, R.; Ndegwa, E.; Kaseloo, P.; Wynn, C. Influence of prior pH and thermal stresses on thermal tolerance of foodborne pathogens. Food Sci. Nutr. 2019, 7, 2033–2042. [Google Scholar] [CrossRef]
- Dawan, J.; Juhee, A. Bacterial stress responses as potential targets in overcoming antibiotic resistance. Microorganisms 2022, 10, 1385. [Google Scholar] [CrossRef]
- Somero, G.N. The cellular stress response and temperature: Function, regulation, and evolution. J. Exp. Zool. 2020, 333, 379–397. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Kline, M.P.; Bimston, D.N.; Cotto, J.J. The heat-shock response: Regulation and function of heat-shock proteins and molecular chaperones. Essays Biochem. 1997, 32, 17–29. [Google Scholar] [PubMed]
- Miller, D.J.; Fort, P.E. Heat shock proteins regulatory role in neurodevelopment. Front. Neurosci. 2018, 12, 821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gross, C.A. Cold shock response in bacteria. Ann. Rev. Genet. 2021, 55, 377–400. [Google Scholar] [CrossRef]
- Cardoza, E.; Singh, H.C. group-mediated antibiotic stress mimics the cold shock response. Curr. Microbiol. 2021, 78, 3372–3380. [Google Scholar] [CrossRef] [PubMed]
- Keto-Timonen, R.; Hietala, N.; Palonen, E.; Hakakorpi, A.; Lindstrom, M.; Korkeala, H. Cold shock proteins: A minireview with special emphasis on Csp-family of enteropathogenic Yersinia. Front. Microbiol. 2016, 7, 1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.D.; Kwon, H.Y.; Kim, E.H.; Kim, K.W.; Briles, D.E.; Pyo, S.; Rhee, D.K. Decrease in penicillin susceptibility due to heat shock protein ClpL in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2011, 55, 2714–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, M.; Cui, L.; Iizuka, R.; Komoto, A.; Neoh, H.M.; Watanabe, Y.; Hishinuma, T.; Hiramatsu, K. walK and clpP mutations confer reduced vancomycin susceptibility in Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3870–3881. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.H.; Novak, J.T.; Knocke, W.R.; Pruden, A. Elevation of antibiotic resistance genes at cold temperatures: Implications for winter storage of sludge and biosolids. Lett. Appl. Microbiol. 2014, 59, 587–593. [Google Scholar] [CrossRef]
- Bagheri, H.; Ghaemi, A.; Aslani, M.; Mozafari, N.; Livani, S.; Dadgar, T. The prevalence of enteroaggregative Escherichia coli in cases of diarrhea in Gorgan. J. Gorgan Univ. Med. Sci. 2008, 2, 8–12. [Google Scholar]
- Pickbourn, L.; Ndikumana, L. Does Health Aid Reduce Infant and Child Mortality from Diarrhoea in Sub-Saharan Africa? J. Dev. Stud. 2019, 55, 2212–2231. [Google Scholar] [CrossRef]
- Abbasi, A.; Yoosefi, M. The study of effective factors on persisted diarrhea in under five year old children Gorgan and Agh-Ghala health center. J. Gorgan Univ. Med. Sci. 2002, 10, 36–41. [Google Scholar]
- Black, R.E.; Brown, K.H.; Becker, S.; Alim, A.R.M.A.; Merson, M.H. Contamination of weaning foods and transmission of enterotoxigenic Escherichia coli diarrhoea in children in rural Bangladesh. Trans. R. Soc. Trop. 1982, 76, 259–264. [Google Scholar] [CrossRef] [PubMed]
- El-Baz, A.H. Prevalence, molecular characterization and antimicrobial resistance of Vero toxigenic E. Coli in fresh soft cheese, ice cream and yoghurt in Mansoura city. Alex. J. Vet. Sci. 2019, 62, 38–46. [Google Scholar] [CrossRef]
- Claeys, W.L.; Cardoen, S.; Daube, G.; De Block, J.; Dewettinck, K.; Dierick, K. Raw or heated cow milk consumption: Review of risks and benefits. Food Control 2013, 31, 251–262. [Google Scholar] [CrossRef]
- Forbes, B.A.; Sahm, D.F.; Weissfeld, A. Bailey & Scott’s Diagnostic Microbiology, 12th ed.; Mosby: London, UK, 2007; pp. 1, 56. [Google Scholar]
- Goli, M.; Ezzatpanah, H.; Ghavami, M.; Chamani, M.; Nedaeinia, R. Multiplex-polymerase chain reaction as a mastitis screening test for major pathogens in dairy cattle farms at different size scales and in several parities. J. Res. Agric. Sci. 2012, 8, 23–33. [Google Scholar]
- Cox, F.E.G.; Wakelin, D.; Gillespie, S.H.; Despommier, D.D. Topley & Wilson’s Microbiology and Microbial Infections: Parasitology; Arnold, E., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1999; 608p. [Google Scholar]
- Murray, P.R.; Baron, E.J.; Jorgenson, J.H.; Pfaller, M.A.; Yolken, R.H. Manual of Clinical Microbiology; American Society for Microbiology Press: Washington, DC, USA, 2003; Volume 47, pp. 625–626. [Google Scholar]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy: The Principles of Numerical Classification; Freeman: San Francisco, CA, USA, 1973; 573p. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Khakpour, M.; Garedaghi, Y. Molecular differentiation of sheep and cattle isolates of Fasciola hepatica using RAPD-PCR. Arch. Razi Inst. 2012, 67, 109–115. [Google Scholar]
- Yang, Z.X.; Zhou, Y.B.; Xiang, X.Z.; Zhu, Z.B.; Pen, L.; Luo, Y.W.; Lu, J. The molecular ecology analysis of microbial communities in waste water–based mud. Pet. Sci. Technol. 2013, 31, 887–894. [Google Scholar] [CrossRef]
- Ong, K.H.; Wei, C.K.; Jing, Y.Q.; Zi, X.L.; Sathish, A.; Mahathir, H.; Cliff, C.; Kelyn, L.G.S.; Siyao, G.; Moon, Y.F.T.; et al. 2020. Occurrence and antimicrobial resistance traits of Escherichia coli from wild birds and rodents in singapore. Int. J. Environ. Res. Public Health 2020, 17, 5606. [Google Scholar] [CrossRef]
- World Health Organization. Ending Preventable Child Deaths from Pneumonia and Diarrhoea by 2025: The Integrated Global Action Plan for Pneumonia and Diarrhoea (GAPPD); World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Kusukawa, N.; Yura, T. Heat shock protein GroE of Escherichia coli: Key protective roles against thermal stress. Genes Dev. 1988, 2, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Soomro, A.H.; Arion, M.A.; Khaskheli, M.; Bhutto, B. Isolation of Escherichia coli from row milk productions in relation to public health sold under market conditions at Tandojam. Pak. J. Nutr. 2002, 1, 151–152. [Google Scholar]
- Fazlara, A.; Maktabi, S.; Norouzi, F. Comparative survey on predictive impediometric models for microbial load in vanilla and cocoa ice-creams produced with traditional and industrial methods. Int. Iran Congress Microbiol. 2016, 12, 69–80. [Google Scholar]
- Bonyadian, M.; Moshtaghi, H.; Akhavan-Taheri, M. Molecular characterization and antibiotic resistance of enterotoxigenic and entero-aggregative Escherichia coli isolated from raw milk and unpasteurized cheeses. Vet Res. Forum. 2014, 5, 29–34. [Google Scholar] [PubMed]
- Ko, K.S.; Hong, S.K.; Lee, K.H.; Lee, H.K.; Park, M.Y.; Miyamoto, H. Detection and identification of Legionella pneumophila by PCR-restriction fragment length polymorphism analysis of the RNA polymerase gene (rpoB). J. Microbiol. Methods 2003, 54, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Negoro, E.; Iwasaki, H.; Tai, K.; Ikegaya, S.; Takagi, K.; Kishi, S. Utility of PCR amplification and DNA microarray hybridization of 16S rDNA for rapid diagnosis of bacteremia associated with hematological diseases. Int. J. Infect. Dis. 2013, 17, 271–276. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Positive Isolates (%) | Characteristics | Positive Isolates (%) |
|---|---|---|---|
| Indole | 100 | Fluorescent color in LMX medium | 71 |
| Green metallic sheen (Eosine methylene blue) | 71 | Blue color in LMX medium | 84 |
| Violet blue colonies (Chromo cult Coliform agar) | 83 | Urease | 13 |
| Methyl red (MR) | 95 | Motility | 88 |
| Acetoin (Vogues-Proskauer Test)(VP) | 2 | Red colonies on MacConkey agar | 85 |
| Gelatin liquefaction | 8 | ||
| Coliform Phenons from Ice Cream A | Reference | From Milk | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Characteristics | 1(34) | 2(4) | 3(2) | 4(1) | 5(1) | 6(1) | 7(4) | 8(1) | E. coli | 9(13) |
| Gram reaction | − | − | − | − | − | − | − | − | − | − |
| Catalase production | + | + | + | + | + | + | + | + | + | + |
| Oxidase production | − | − | − | − | − | − | − | − | − | − |
| KCN, growth | − | − | + | − | ND | ND | ND | ND | + | (+) |
| Dulcitol | + | + | + | − | ND | ND | ND | ND | + | (+) |
| Acetate utilization | + | + | + | + | ND | ND | ND | ND | + | (+) |
| Nitrate reduction | + | + | + | + | + | + | + | + | + | 80 |
| Fluorescence production on LMX | + | − | − | − | − | − | − | − | + | 65 |
| Bluish-green color on LMX | + | + | + | + | − | − | − | − | + | + |
| Fermentative metabolism | + | + | + | + | + | + | + | + | + | (+) |
| Lysine decarboxylase | + | 50 * | − | + | − | + | 50 * | − | + | (+) |
| Ornithine decarboxylase | + | + | + | 25 * | ND | ND | ND | ND | + | (+) |
| Green metallic sheen colonies on Eosine methylen blue (EMB) agar | + | − | − | − | − | − | − | − | + | ND |
| Pigment production | − | − | + | − | − | − | − | − | − | − |
| Red colonies on MacConkey agar | + | + | + | − | − | + | − | − | + | + |
| Gelatin liquefaction | 9 * | − | − | − | − | − | 25 * | − | − | − |
| Dark blue and violet colonies on CC agar | + | + | 50 * | + | − | − | − | − | + | + |
| Acidification (Acid /acid + gas) | + | + | + | − | − | + | − | − | + | + |
| Alkali form (alkaline/acid) | − | − | − | + | + | − | − | + | − | 16 * |
| Methyl red (MR) | + | + | + | − | + | − | + | + | + | + |
| Acetoin production(Vogues-Proskauer) Test)(VP)) | − | − | − | − | − | + | − | − | − | − |
| Gas from glucose | + | + | + | − | − | + | − | − | + | + |
| H2S from cystein | − | − | − | − | − | − | − | − | − | 13 * |
| Indole production | + | + | + | + | + | + | + | + | + | + |
| Motility | + | + | + | − | + | − | − | + | + | + |
| Urease production | − | − | − | − | + | + | + | − | − | − |
| Glucose | + | + | + | + | + | + | + | + | + | + |
| Lactose | + | + | ND | − | − | + | − | ND | + | + |
| Sucrose | (+) | (+) | ND | − | − | + | − | ND | (+) | + |
| Maltose | (+) | (+) | ND | (+) | − | + | − | ND | (+) | + |
| Melibiose | (+) | (+) | ND | (+) | − | + | − | ND | (+) | + |
| Cellobiose | − | − | ND | − | − | + | − | ND | − | 17 * |
| Trehalose | + | + | ND | + | − | + | − | ND | + | + |
| Sorbitol | (+) | (+) | ND | (+) | − | + | − | ND | (+) | ND |
| Citrate | − | − | − | − | + | + | − | − | − | ND |
| Malonolate | − | − | ND | − | − | + | − | ND | − | ND |
| Mucate | + | + | (+) | (+) | ND | ND | ND | ND | + | ND |
| d-Adonitol | − | − | − | − | ND | ND | ND | ND | − | − |
| d-Arabitol | − | − | − | − | ND | ND | ND | ND | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranjbar, M.; Nedaeinia, R.; Goli, M.; Shahi, S. Evaluation of the Thermal Processes on Changing the Phenotypic Characteristics of Escherichia coli Strains from Ice Cream Compared to Non-Pasteurized Milk. Fermentation 2022, 8, 730. https://doi.org/10.3390/fermentation8120730
Ranjbar M, Nedaeinia R, Goli M, Shahi S. Evaluation of the Thermal Processes on Changing the Phenotypic Characteristics of Escherichia coli Strains from Ice Cream Compared to Non-Pasteurized Milk. Fermentation. 2022; 8(12):730. https://doi.org/10.3390/fermentation8120730
Chicago/Turabian StyleRanjbar, Maryam, Reza Nedaeinia, Mohammad Goli, and Sharifeh Shahi. 2022. "Evaluation of the Thermal Processes on Changing the Phenotypic Characteristics of Escherichia coli Strains from Ice Cream Compared to Non-Pasteurized Milk" Fermentation 8, no. 12: 730. https://doi.org/10.3390/fermentation8120730